Предки динозавров
Уже несколько десятилетий палеонтологи и эволюционные биологи ищут ответ на вопрос, как выглядели самые ранние родственники динозавров. Большинство учёных уверены в том, что по размерам они походили на современных кур и передвигались на двух ногах.
Однако открытие нового вида, получившего название Teleocrater rhadinus, перевернула представления исследователей. Останки древнего плотоядного родственника динозавров были найдены на территории современной Танзании. По окаменелостям можно предположить, что животное достигало 2-3 метров в длину, имело длинный хвост и длинную шею. А передвигалось оно на четырёх лапах, похожих на конечности нынешних крокодилов.
В международных исследованиях принял участие Национальный научный фонд США, и, как отмечается в пресс-релизе, T. rhadinus заполнил пробел в эволюционной истории ранних динозавров.
"Исследование проливает свет на распространение и разнообразие предков крокодилов, птиц и динозавров", — рассказывает соавтор работы Джуди Ског (Judy Skog).
её словам, представители открытого вида жили в среднем триасе, 247-242 миллионов лет назад – сразу после того, как группа рептилий, известных как архозавры, раскололась на две ветви. Представители первой эволюционировали в птиц, а вторая ведёт к аллигаторам и крокодилам. T. rhadinus и его родственники являются самыми ранними известными членами ветви птиц из архозавров.
Как сообщила группа российских палеонтологов, которая также принимала участие в исследовании, на основании находок удалось реконструировать анатомию рептилии T. rhadinus. Это позволило выделить новую группу вымерших архозавров – Aphanosauria. К ней также была отнесена рептилия Dongusuchus из Южного Приуралья. Ещё в 1990-х годах палеонтолог Андрей Сенников из Палеонтологического института им. А. А. Борисяка РАН высказал предположение, что российский Dongusuchus относится к особой группе архозавров, близкой к динозаврам, и сейчас существование этой группы потвердилось.
"Группа Aphanosauria мозаичным образом сочетает динозавровые и крокодиловые признаки, она появилась ещё до разделения двух этих линий", — поясняет учёный. В частности, позвонки, плечевая кость и челюсть у Teleocrater и других представителей этой промежуточной группы устроены как у динозавров, а пяточная кость и голеностопный сустав – как у крокодилов. Как доказывает находка Dongusuchus в Оренбуржье, такие "гибридные" существа когда-то бродили и по территории России.

Любопытно, что останки этой группы ящеров впервые были обнаружены ещё в 1933 году в той же Танзании британской командой Френсиса Рекса Паррингтона (Francis Rex Parrington). Тогда палеонтологи не смогли классифицировать животное: у них имелось лишь несколько позвонков, задние конечности и части передних, а также кости плечевого и тазового поясов. Находки отправились в Лондонский музей естествознания.
В 1950-х сотрудник музея Алан Чериг (Alan Charig) подробно изучил останки из Танзании и предположил, что они принадлежат некоему неизвестному предку динозавров (он посмертно указан в качестве одного из авторов текущей публикации). Именно Чериг предложил видовое название Teleocrater rhadinus.
Позднее американские палеонтологи заинтересовались останками древнего родственника динозавров и в 2015 году провели дополнительные раскопки на месте их обнаружения. Так были найдены новые окаменелости, которые позволили восстановить практически полный скелет и определить, что он принадлежит представителю архозавров с крокодилоподобными и птицеподобными чертами.
Палеонтологи планируют вновь отправиться в Танзанию в мае этого года, чтобы найти ещё больше останков и составить полный скелет T. rhadinus.
"Очень увлекательно решать такие головоломки. Мы можем наконец собрать воедино разрозненные окаменелости и пролить свет на широкие анатомические и биогеографические тенденции этой группы животных", — заключает соавтор работы палеобиолог Мишель Стокер (Michelle Stocker).
Статья с описанием раннего предка динозавров опубликована в журнале Nature.
Напомним, что ранее учёные в ходе изучения предка тираннозавров выявили пример необычной эволюции.
Кстати, мы также рассказывали об исследовании, которое доказало: динозавры и их предки могли видеть сны.
Источник: www.vesti.ru
Однако и они уже летали в меловом периоде. Так, в 2005 году на антарктическом острове Сеймур обнаружили кости птицы, необычайно похожей на современную утку, – вегависа.
Джулия Кларк из Техасского университета в Остине провела классический анатомический анализ остатков вегависа (Vegavis iaai) возрастом 67 миллионов лет и сделала цифровую реконструкцию его скелета, что позволило отнести его к той же группе, куда входят современные утки и гуси.
2016 году палеонтологи изучили второй, более полный скелет вегависа и установили, что эта птица не только выглядела как утка, но и, вполне вероятно, крякала по-утиному. В окаменелости сохранился отпечаток самого древнего известного сиринкса – своеобразного «динамика», как две капли воды схожего с голосовым аппаратом современных гусеобразных. «Вегависа можно назвать одной из важнейших окаменелостей, проливающих свет на происхождение современных птиц», – объясняет Дэниел Филд, биолог-эволюционист из Батского университета.
А годом раньше группа ученых под руководством профессора орнитологии Йельского университета Ричарда Прума изучила гены 198 видов обитающих на планете птиц и сопоставила результаты с данными по окаменелостям. Согласно построенному ими подробному филогенетическому древу пернатых, лишь три современные группы птиц возникли до начала кайнозойской эры, когда и начался расцвет известных сегодня отрядов.
Чтобы увидеть таких птиц воочию, я направляюсь в город Кеммерер (штат Вайоминг). Он находится в 160 километрах к северо-востоку от Солт-Лейк-Сити среди холмов с осыпающимися склонами и в буквальном смысле построен на костях, возраст которых составляет 52 миллиона лет. Здешние суровые засушливые земли известны прежде всего множеством окаменевших рыб, что, в общем-то, закономерно, ведь в середине палеогенового периода тут, посреди субтропической болотистой равнины, располагалось огромное озеро – жители юга Флориды легко узнали бы в этих местах родные соленые заводи. Донные отложения водоема, как и меловые озера Китая, «заморозили во времени» целую экосистему, включая множество древних птиц. Палеонтологи обнаружили здесь более сотни целых птичьих скелетов. «Это самое полное местонахождение эоценовой эпохи», – поясняет Лэнс Гранде, хранитель Филдсовского музея и мой проводник к месту раскопок.
Лэнс посещает это палеонтологическое эльдорадо, известное под будничным названием «озеро окаменелостей», уже более 40 лет. Обычно летом он по несколько недель проводит раскопки на частной земле, которую берет в аренду.
В конце июня, когда я присоединяюсь на неделю к его команде, здесь стоит страшная жара. Работающие на одном из холмов старшеклассники-энтузиасты и волонтеры из музея обучают меня, как нужно извлекать массивные каменные плиты из отложений и высматривать на них следы древней жизни. Однажды поздним утром, когда земля вокруг буквально дымилась под испепеляющими солнечными лучами и я была в седьмом поту, Лэнс попросил меня прерваться: он хотел показать что-то необыкновенное. Рабочий с соседнего карьера принес… птицу.
Потенциальный трофей прибыл к нам аккуратно уложенным в пыльном кузове пикапа. Остатки пернатого лишь чуть-чуть выступали из-под слоя известняка, однако я отчетливо разглядела тонкие косточки и отпечаток крыла с перьями. Лэнс захотел повнимательнее ознакомиться с находкой, так что мы бережно завернули ее и помчались в местную больницу, чтобы сделать рентген. Рентгенологи встретили нас без ожидаемого мной удивления – очевидно, мы привезли к ним далеко не первого «пациента». Изучив несколько снимков, к концу нашего спонтанного визита Лэнс убедился, что внутри каменной плиты был скрыт целый скелет, – находку стоило забрать с собой.
Такой путь – из Кеммерера в Чикаго – проделало множество птиц, обнаруженных здесь. Сейчас они обустроились на выставочных стендах и в хранилищах Филдсовского музея. Несколько недель спустя после нашей экспедиции я побывала в легендарном музее. Мне удалось рассмотреть вблизи древнего попугая, представителя певчих воробьиных, а также одну из недавно описанных учеными птиц-мышей. Все эти остатки доказывают: в экосистемах середины палеогенового периода сложилось необыкновенное разнообразие пернатых. «Каждый второй найденный образец птицы оказывается чем-то совершенно новым, – делится Лэнс. – Просто поразительно!»
В наше время птицы-мыши, вроде данного музейного образца, встречаются лишь в Африке к югу от Сахары. Однако хрупкая окаменелость, хранящаяся в этих пузырьках, свидетельствует о том, что древние птицы-мыши Tsidiiyazhi abini около 60 миллионов лет назад обитали на юге Северной Америки. Благодаря редкой находке ученым удалось уточнить время разделения разных ветвей родословного древа птиц и подтвердить, что они претерпели настоящий эволюционный взрыв в начале кайнозойской эры.
тография сделана в Музее естественных наук и истории Нью-Мексико, Альбукерке.
Последние годы оказались богатыми на находки для ученых, пытающихся разобраться с тем, как налаживалась жизнь пернатых в кайнозойскую эру. Недавно в Нью-Мексико палеонтологи извлекли остатки еще одной птицы-мыши, жившей 62 миллиона лет назад, которую назвали Tsidiiyazhi abini, что можно перевести как «маленькая утренняя птичка». Сейчас она считается одной из древнейших кайнозойских птиц. Вроде бы мелочь, но очень важная. Во-первых, находка показывает, что эти небольшие длиннохвостые птички, оби-тающие в наши дни лишь в Африке к югу от Сахары, появились совсем в другой части света и в первой половине кайнозойской эры населяли леса почти по всей планете. А во-вторых, птицы-мыши, прекрасно умеющие карабкаться по стволам деревьев, представляют собой специализированных потребителей плодов именно цветковых растений и сейчас помогают многим видам распространять семена далеко от родительского растения. (Свое название пичуги получили за то, что пушисты и умеют продираться сквозь самые густые и колючие кусты.)
Не менее интересная находка – гигантский пингвин вайману (Waimanu manneringi) возрастом 61 миллион лет, недавно обнаруженный в Новой Зеландии. Присвоенное ему учеными имя на языке майори означает «водяная птица» – вайману действительно уже не летал, а только плавал, хотя по стилю плавания больше напоминал современных гагар (с которыми они близкие родственники), чем пингвинов, – греб лапами, а не крыльями.
оме того, изучение пигментов в перьях другого ископаемого пингвина, инкаяку (Inkayacu paracasensis) – гиганта полутораметровой длины, – выявило, что древние пингвины были не иссиня-черными, как современные, а, скорее, красновато-коричневыми. (Инкаяку, что с языка коренного народа Перу, кечуа, переводится как «король воды», з6 миллионов лет назад нырял за рыбой у тихоокеанского побережья Южной Америки.) В этой, казалось бы, мелочи – ну, подумаешь, черный он был или бурый – кроются большие эволюционные преобразования: ведь пигментные тельца, меланосомы, у черных пингвинов крупные и выложены в перьях ровными плоскими рядами, что позволяет уменьшить сопротивление среды при движении под водой, придавая перьям упругость и жесткость.
Все эти окаменелости, похоже, неплохо вписываются в данные последних генетических исследований: в 2014–2015 годах вышло несколько работ, в которых ученые, изучив полные геномы 48 ныне живущих птиц, и пришли к выводу, что бурный расцвет разнообразия современных пернатых начался именно в кайнозойскую эру.
«Эволюции потребовались десятки миллионов лет, чтобы на свет появились маленькие крылатые динозавры, способные летать, взмахивая передними конечностями. Такое строение тела оказалось весьма удачным, – подытоживает Стивен Брюсатт, палеонтолог из Школы геонаук Эдинбургского университета. – И ныне этим существам принадлежит весь мир!»
Озеро окаменелостей
Хотя ископаемые рыбы (1) встречаются в известняках «озера окаменелостей» в Вайоминге гораздо чаще, отсюда уже извлечено более сотни полных отпечатков птиц. Среди них встречаются птицы-мыши (2), попугаи (3), нелетающие представители отряда журавлеобразных (4) и фрегаты (5). В слоях возрастом 52 миллиона лет скрыта богатейшая «коллекция» невероятно прекрасно сохранившихся окаменелостей – вроде этого отпечатка древней певчей птицы (6). Такое обилие находок великолепной сохранности создает, по словам Лэнса Гранде из Филдсовского музея, «идеальный шторм в палеонтологии». Фотографии сделаны в Филдсовском музее естественной истории, Чикаго.
Источник: nat-geo.ru
Международная группа палеонтологов из США, Великобритании, ЮАР, Швеции и России (Казанского университета, Палеонтологического института им. А.А. Борисяка РАН) восстановили облик предшественников динозавров. По окаменелостям, найденным при раскопках на юге Танзании в 2015 году, они описали новый вид существа, жившего 245 млн лет, назвав его Teleocrater rhadinus.
Это было недостающее звено в эволюции между архозаврами и динозаврами. Древние пресмыкающиеся архозавры были предками, с одной стороны, крокодилов, с другой — динозавров и птиц. Но если от архозавров к крокодилам по палеонтологических находкам прослеживалась четкая эволюционная линия с переходными формами существ, то в направлении динозавров в линии эволюции зияла пустота, динозавры появились на Земле словно ниоткуда. Единственное, что могли сделать ученые, это по редким находкам триасовых окаменелостей воссоздать гипотетических предков динозавров, сильно смахивавших на ощипанную курицу, довольно шустро бегавшую на двух ногах.
Нынешняя реконструкция по танзанийским находкам, относящимся к середине триасового периода, заставляет по-новому взглянуть на предка динозавров. Он был довольно крупным существом длиной от двух до трех метров, высотой «в холке» около метра и весом 10–30 кг. С длинной шеей и массивным длинным хвостом внешне он был похож на динозавров. Но передвигался Teleocrater rhadinus на четырех конечностях (а не на двух, как динозавры), а анатомия голеностопного сустава и пропорции задних конечностей у него были характерные для крокодилов. Судя по редким и большим острым зубам, он был хищником.
Теперь Teleocrater rhadinus ликвидировал пробел в эволюционном древе и занял место на его развилке, ведущей от архозавров к крокодилам в одну сторону и к динозаврам, птеродактилям и птицам в другую.
В триасовом периоде эти пресмыкающиеся расселились по всей Пангее. А когда в следующем юрском периоде гигантский сверхконтинент раскололся на Лавразию (позже она, в свою очередь, распалась на Евразию и Северную Америку) и Гондвану (давшую начало всем остальным современным материкам), триасовые животные столкнулись уже с настоящими динозаврами. И если крокодилы выжили, уйдя с суши в моря и пресные водоемы, то Teleocrater rhadinus и его родичи (ученые их объединили в группу Aphanosauria — скрытые, или тайные ящеры) вчистую проиграли в жизненной борьбе более мощным и более стремительным юрским динозаврам и исчезли с лика Земли.
Как считают палеонтологи, открывшие Teleocrater rhadinus, следы жизни этих еще недодинозавров следует искать на территории современных России, Индии и Бразилии, во всяком случае, именно здесь отдельные фрагменты их скелетов уже находили ранее.
Сергей Петухов
Источник: www.kommersant.ru
Предки динозавров и птиц
Современные виды птиц и крокодилов произошли от древней группы архозавров (Archosauria), которые обитали на суше и в воздухе около 252–66 млн лет назад. Самые известные представители этой группы — динозавры, от которых, как принято считать, и происходят птицы. Но кроме динозавров существовали множество других групп архозавров. Ученые делят их на две крупные эволюционные линии — крокодилью и птичью.
Птичья линия архозавров (птицестопые) — это динозавры и их близкие родственники, в том числе птицы. В основном птицестопые — двуногие и легкотелые длинношеие животные. Но их эволюционная линия не была изучена — наука не знала о наиболее примитивных представителях этой группы. Статья, которая опубликована в Nature, раскрывает этот этап эволюции архозавровых рептилий.
Рептилия с признаками динозавра и крокодила
Ученые описали новый вид архозавра, который жил около 247–235 млн лет назад на территории современной Танзании. Его назвали Teleocrater rhadinus. У телеократера были признаки как динозавров (особенности строения костей черепа и позвоночника), так и крокодилов (строение голеностопного сустава). Высота архозавра вида Teleocrater rhadinus достигала трех метров. Они жили на древнем континенте Пангея.
Благодаря открытию этого вида ученые смогли выявить отдельную группу, о которой раньше ничего не было известно. Ее представителей назвали афанозаврами (Aphanosauria).
Афанозавры были стройными четвероногими хищниками с длинной шеей и хвостом — этим они были были похожи на динозавров, но с короткими конечностями — почти как у крокодилов.
В состав группы входили несколько рептилий из разных уголков Пангеи — некогда единого суперконтинента на планете. Известно, например, что афанозавры обитали в Бразилии, Индии и России.
Почему открытие афанозавров — это важно
Палеонтологи выяснили, что животные этой группы более родственны динозаврам и птицам, чем крокодилам. Афанозавры признаны самыми примитивными представителями линии птицестопых архозавров. Это открытие заполнило очень важный пробел в эволюции архозавров — от массивных четвероногих животных, похожих на современных крокодилов (примитивных архозавров), до более стройных и быстро бегающих динозавров.
Главный вывод ученых заключается в том, что наиболее примитивные предки динозавров и птиц были четвероногими животными с лапами как у крокодилов. А уже это заставляет пересмотреть традиционную гипотезу о том, что предки динозавров были стройными, двуногими, быстро бегающими животными.
Читайте подробности об открытии в материале «Элементов» «Обнаружена новая группа ископаемых рептилий — отдаленных родственников динозавров и птиц»
Источник: tvrain.ru
Динозавры первыми «встали на ноги»
Одно из основных направлений эволюции диапсид – формирование ажурной и подвижной структуры черепа.
Кроме того, у диапсид могут постоянно отрастать зубы взамен поломанных или утраченных. За это «удобство» приходится дорого платить: беспорядочная смена зубов исключает окклюзию (точную подгонку верхних и нижних зубов друг к другу). Поэтому у диапсид отсутствует зубной аппарат режущего типа.
Диапсиды по-своему решили общую для наземных позвоночных проблему передвижения на суше. Многие из них перешли к бипедии, т. е. к хождению на двух ногах, что сопровождалось редукцией передних конечностей, а некоторые вовсе утратили ноги, как, например, змеи.
Ко всему, диапсиды, по-видимому, никогда не обладали гомойотермией – физиологически обусловленной стабильной температурой тела. Исключение составляют только птицы, да у некоторых крупных динозавров инерционное сохранение более или менее постоянной температуры обеспечивалось благодаря большим размерам тела.
К ДИАПСИДАМ (т. е. имеющим в черепе два отверстия для выхода челюстной мускулатуры и две скуловые дуги) относятся динозавры в широком смысле, их триасовые предшественники – архозавры и потомки: крокодилы, ящерицы, змеи, а также птицы.
Для диапсид характерна ажурная и подвижная структура черепа. Например, ящерицы и змеи, раскрывая рот, не только опускают вниз нижнюю челюсть, но могут также приподнимать и опускать переднюю часть верхней челюсти. Нижнечелюстной сустав может раздвигаться. Обе половинки нижней челюсти соединены между собой эластичными связками. У диапсид фактически нет мозговой коробки: полость, в которой расположен мозг, открыта спереди и представляет собой подобие трубки.
Сам мозг как у древних, так и у современных диапсид (кроме птиц) очень примитивен.
Конструктивное единство элементов черепа у синапсид позволило им уже в триасе сформировать монолитную черепную коробку. Число костей нижней челюсти у синапсид неуклонно уменьшалось, и в итоге из шести костей осталась только одна – зубная.
Такое монолитное строение черепа стало, по существу, преадаптацией к дальнейшему развитию мозга.
Млекопитающие (а также пеликозавры и терапсиды, в том числе териодонты — «зверозубые рептилии») относятся к СИНАПСИДАМ, у которых в черепе имеется только одно отверстие (височное окно) для выхода челюстной мускулатуры
В отличие от диапсид, у синапсид никогда не формировалась подвижная конструкция черепа: со временем последний, наоборот, становился все более монолитным. Это можно рассматривать как преадаптацию к совершенствованию мозга, которое, впрочем, началось гораздо позже.
Со временем количество зубов и их положение в челюсти у синапсид стабилизировалось, что сделало возможным окклюзию. Это, в свою очередь, позволило осуществлять первичную обработку пищевого комка, чего никто из диапсид – ни динозавры, ни современные крокодилы, ящерицы и змеи – делать не способны.
В отношении опорно-двигательного аппарата синапсиды вначале уступали диапсидам: ни в перми, ни в триасе они так и не не научились передвигаться на двух ногах. Передвижение же на четырех конечностях эффективно, только если существо имеет вертикально поставленные конечности, а пермо-триасовые синапсиды как бы «висели» между своими ногами. Латеральное (боковое) положение конечностей сохраняли еще и некоторые мезозойские млекопитающие – такое положение крайне неэкономно с точки зрения расхода мышечной энергии
В совершенствовании локомоторного аппарата синапсиды вначале уступали диапсидам. Перевести ноги в вертикальное положение синапсидам удалось лишь в конце триаса, уже на стадии млекопитающих – для этого потребовались очень крупные преобразования опорно-двигательного аппарата.
Что касается гомойотермии, то, в отличие от диапсид, первые попытки перехода к ней, по-видимому, были у синапсид уже в карбоне. Вспомним пеликазавров с их огромным «парусом» на спине, который специалисты все более склонны рассматривать как средство стабилизации температуры тела.
Диапсиды и синапсиды: великое противостояние
Именно синапсиды первыми в истории Земли создали в пермском периоде богатую и разнообразную фауну позвоночных суши. Только два региона сохранили для нас в большом количестве ее окаменевшие остатки: Южная Африка и Русская платформа (бассейн Северной Двины и Башкирия). Среди пермских синапсид были крупные сильные хищники размером с тигра, с огромными кинжаловидными клыками – горгонопсиды (иностранцевия и др.); были охотники на мелкую добычу, аналоги куньих и псовых; существовали и растительнояды, например, листрозавры, каннемейериды – большие тяжеловесные животные.
К началу юры диапсиды «переиграли» синапсид, очевидно, в основном именно благодаря изобретению двуногой локомоции. Хищные диапсиды (динозавры) полностью вытеснили хищных синапсид. Интересно, что растительноядные синапсиды пережили свое время и сосуществовали с хищниками из диапсид. Как будет рассмотрено ниже, растительноядность вообще, в том числе на базе диапсид, формировалась в процессе эволюции «с трудом». В целом же биота синапсид была почти нацело «стерта» сообществом диапсид, господство которых продолжалось с конца триаса до конца мела.
Расцвет диапсид закончился в позднем мелу, однако многие диапсиды (крокодилы, змеи и ящерицы) прекрасно существуют и в наши дни, а птицы делят с млекопитающими господствующее положение среди позвоночных животных суши. Но история синапсид на этом не закончилась, напротив – в кайнозое для них наступила новая эпоха расцвета. Как же синапсиды пережили долгий период господства диапсид, какие изменения они претерпели за это время, и чем было обусловлено их «возрождение» уже в виде млекопитающих?
Давид против Голиафа
Первые представители млекопитающих произошли в позднем триасе от мелких, эволюционно продвинутых териодонтов. Эти неспециализированные плотояды сумели закрепиться в динозавровой биоте лишь потому, что были очень мелкими, размером с мышь, что позволило им выйти из-под пресса хищных динозавров.
Например, для тиранозавра высотой до 5 м и весом 1,5—2 т охота на жертву, вес которой 10—20 г, была не только бессмысленной, с точки зрения энергетических затрат, но и безуспешной. Это поставило мезозойских синапсид вне пищевых цепей и блоков динозавровой биоты и дало им возможность просуществовать «в тени» динозавров более 150 млн лет, что составляет примерно две трети истории класса млекопитающих.
Две трети своей истории млекопитающие буквально «проскакивали между пальцами» крупных хищных динозавров благодаря своим мелким размерам
Важным событием в эволюции наземной биоты было появление многобугорчатых в юрском периоде: фактически впервые в истории Земли млекопитающие сформировали эффективного растительнояда в мелком размерном классе. Дело в том, что переваривать грубую растительную пищу позвоночные способны только с помощью симбиотических кишечных микроорганизмов. Все растительноядные диапсиды были очень крупными, потому что большая масса тела обеспечивала инерционное сохранение относительно стабильной температуры, необходимой для существования кишечной флоры. В мелком размерном классе такое невозможно: кишечная флора будет погибать при любом охлаждении тела (например, ночью). Поэтому эффективное растительноядение у мелких позвоночных возможно только при наличии гомойотермии, присущей млекопитающим.
Первые относительно крупные теплокровные хищники-млекопитающие размером с кошку представляли ощутимую угрозу для детенышей динозавров мелового периода
Остальные млекопитающие юры и мела были также мелкие существа, преимущественно плотоядные. Исключение составляют триконодонты. В юре и в раннем мелу они, как и все другие млекопитающие, были очень маленькими, размером не больше мыши. Однако во второй половине мела среди них появился макрохищник, гигант среди млекопитающих, достигавший размеров современной кошки или вирджинского опоссума (Jenkins, Schaff, 1988).
До этого момента биоты млекопитающих и динозавров практически не перекрывались, но теперь ситуация изменилась. Детеныши динозавров, только что вылупившиеся из яиц, были не крупнее курицы, имели хрящевой скелет, а по ночам не могли активно двигаться из-за охлаждения. Наверняка, они были легкой и желанной добычей триконодонтов. Эти агрессивные хищники, способные охотиться по ночам благодаря своей теплокровности, могли существенно повлиять на судьбу выводков меловых динозавров (Агаджанян, 2003). Эта гипотеза недавно получила подтверждение: китайские палеонтологи описали триконодонта Repenomamus giganticus с крупными островершинными зубами и массивной челюстью (длина его тела превышала 1 м, а вес достигал 12—14 кг), в желудке которого были найдены остатки молодого растительноядного динозавра (Hu et al., 2005).
Роль мезозойских млекопитающих в структуре биоты Земли не ограничивалась подобным ощутимым прессом на динозавров. У вышеупомянутых многобугорчатых щечные зубы имели структуру коронки по типу хомякообразных, а резцы по форме и строению напоминали резцы грызунов. С точки зрения функции и положения в сообществе многобугорчатые являлись высокоспециализированными «грызунами». Анализ их жевательного аппарата показал, что они питались довольно крупными объектами (Gambaryan, Kielan-Jaworowska, 1995). В юре и первой половине мела это могли быть орешки голосемянных (цикадовых, гинговых, араукариевых), в течение второй половины мела и в кайнозое – плоды покрытосемянных, в том числе бука, дуба и др.
Утконос (Ornithorhynchus anatinus), наряду с ехидной, – единственные сохранившиеся представители древнейшей группы яйцекладущих млекопитающих. Обитает утконос в Австралии и Тасмании. В длину достигает полуметра, вес – до 2 кг. Хвост как у бобра, лапы снабжены плавательной перепонкой: животное ведет полуводный образ жизни. Самка откладывает и насиживает 1—2 яйца.
У самцов на задних лапах – ядовитая шпора. Есть предположение, что многие мезозойские млекопитающие могли иметь подобное оружие и использовать его для защиты от динозавров (Hurum et al., 2006)
Радиации многобугорчатых и покрытосемянных в начале второй половины мела совпадают по времени и, по-видимому, были взаимообусловлены, т. е. эти млекопитающие могли играть важную роль в расселении голосемянных, а затем и покрытосемянных. Адаптивный тип этой группы млекопитающих был настолько эффективен, что многобугорчатые перешагнули рубеж мел—кайнозой и просуществовали достаточно долго, до конца олигоцена, успешно конкурируя одно время даже с грызунами.
Современные млекопитающие подразделяются на два подкласса: прототерии и терии. И те и другие были известны уже в позднем триасе. В мезозойских сообществах млекопитающих господствовали прототерии, а терии были «изгоями». В кайнозое ситуация изменилась на противоположную. Чтобы понять причины этого, необходимо сделать экскурс в анатомию.
Между молоточком и наковальней
Имеющиеся данные по строению черепа (Kermack, Kielan-Jaworowska, 1971) и среднего уха однопроходных подтвердили глубокие различия между прототериями и териями, что дает основание предполагать, что они произошли от разных групп триасовых териодонтов. Напомним, что в ходе эволюции нижняя челюсть синапсид постепенно теряла кости, расположенные сзади от зубной кости, которая в свою очередь неуклонно увеличивалась, разрастаясь назад. На стадии первых млекопитающих она пришла в соприкосновение с чешуйчатой костью черепной коробки. В результате возникло новое сочленение нижней челюсти с черепом.
Судя по анатомическим особенностям, разные эволюционные линии млекопитающих разошлись еще на уровне предков
У млекопитающих позднего триаса функционировали оба сочленения: старое (сочленованная кость нижней челюсти – квадратная кость черепной коробки) и новое (зубная кость – чешуйчатая кость). Затем старый сустав исчез, а новый сохранился.
Кости, формировавшие старый сустав, преобразовались в элементы звукопередаточного аппарата. Угловая и сочленованная кости нижней челюсти превратились в барабанную кость и молоточек, квадратная кость черепной коробки – в наковальню (еще один элемент звукопередающего механизма, стремечко, имелся уже у рептилий).
Строение и эволюционная история барабанной кости, наковальни и молоточка различна у прототериев и териев.
По-видимому, именно эти особенности определили различия при модернизации черепа у этих двух групп млекопитающих. Черепа у всех прототериев очень однотипные и практически не менялись на протяжении 200 млн лет вплоть до настоящего времени. У териев, напротив, трехчленность звукопередающего механизма сделала возможной почти неограниченную модернизацию черепа в процессе эволюции и породила многообразие их строения.
Правота этих рассуждений подтверждается тем, что начиная примерно с олигоцена, когда у териев завершилось формирование основных типов черепов, у многих из них срастаются молоточек и наковальня (Weber, 1927). Когда отпадает нужда хотя бы в одном сочленении, эволюция от него избавляется.
Мезозойские прототерии и терии различались также по строению зубного аппарата. По-видимому, изначально высокая специализация зубов прототериев также препятствовала их дальнейшей эволюции: будучи высоко адаптированными к своей динозавровой биоте, они «заблокировали» возможность изменения зубного аппарата.
Все эти принципиальные различия позволяют настаивать на том, что эволюционные линии прототериев и териев разошлись еще на уровне их предков – териодонтов. Современные однопроходные – это, по существу, животные териодонтового уровня, которые в условиях длительной изоляции на Австралийском континенте сохранили до наших дней многие признаки триасовых предков млекопитающих.
Колыбель для мозга
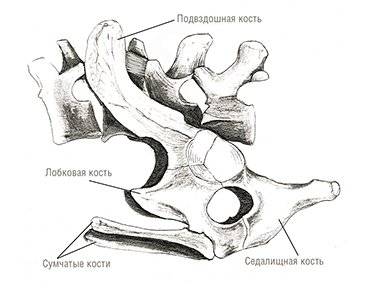
Что же касается обеих современных групп териев – сумчатых и плацентарных – то их представители известны начиная с раннего мела. Однако совершенная плацентарность, проявляющаяся в длительном вынашивании детенышей, по-видимому, развилась не раньше конца мела или начала кайнозоя. Об этом свидетельствуют так называемые сумчатые кости, имеющиеся у териодонтов, всех современных однопроходных и сумчатых.
Эти кости имелись и у ранних плацентарных вплоть до конца мела (Novacek et al., 1997). Однако с начала кайнозоя судьбы сумчатых и плацентарных разошлись: они оказались разделенными на двух блоках континентов (Северном и Южном), и их развитие пошло различными путями.
До возникновения совершенной плацентарности уровень развития головного мозга у всех млекопитающих был почти одинаков. Сумчатые, обитатели южных материков, сохраняют примитивное строение мозга и по сей день. Почему так произошло? В целом сумчатость, как тип размножения, выгоднее для сохранения популяции, чем плацентарность. Самка сумчатых в случае опасности, в отличие от плацентарных, просто выбрасывает детеныша из сумки и убегает. Новое зачатие у сумчатых возможно почти сразу после рождения детеныша. Убегая, самка спасает себя – полноценного производителя, оплодотворенную яйцеклетку и иногда висящего на соске новорожденного детеныша.
Однако при этом большую часть ответственности за судьбу детеныша, размером и формой напоминающего фасолину, несет уже не мать, а он сам, и прежде всего его головной мозг. А успешное функционирование системы и одновременно ее глубокие структурные преобразования невозможны. Кроме того, у детенышей сумчатых развивается более мощная по сравнению с детенышами плацентарных мускулатура, необходимая для сосания, что тормозит развитие черепной коробки и, соответственно, мозга.
Можно предположить, что именно плацентарность сыграла роль спускового механизма для неуклонного прогрессивного развития мозга
Плацентарность максимально продлила внутриутробный период и тем самым сняла с мозга детеныша ответственность за контроль над развивающимся организмом. Можно предположить, что она сыграла роль спускового механизма для развития центральной нервной системы – одного из важнейших условий для прогрессивной модернизации мозга в индивидуальном и историческом развитии. Именно поэтому плацентарные – бывшие изгои динозавровой биоты – дали в кайнозое огромное разнообразие форм и стали господствующей группой.
Кроме того, на южных материках Земли млекопитающие жили на сравнительно небольших и однородных в физико-географическом смысле территориях в условиях ослабленной конкуренции. На огромных же пространствах северных материков условия обитания и сообщества наземных позвоночных были значительно многообразнее, а конкуренция – жестче. В этой ситуации для эволюционного успеха было недостаточно одних лишь морфологических, структурных преимуществ: уже в раннем кайнозое в группе млекопитающих наблюдается быстрая эволюция головного мозга. Палеонтологическая летопись сохранила прямые свидетельства увеличения и усложнения мозга плацентарных на протяжении кайнозоя, что по-видимому, привело к значительному усложнению поведения млекопитающих.
Сложное поведение и обучаемость в ходе эволюции начинали развиваться во многих группах животных, но только у плацентарных млекопитающих эта важнейшая эволюционная тенденция достигла своего наиболее яркого и полного выражения. В течение кайнозоя постоянно совершенствовалась способность плацентарных передавать информацию от поколения к поколению негенетическим путем. Закономерным итогом этого неуклонного прогрессивного развития мозга стало появление человека разумного.
Литература
Агаджанян А. К. Вопросы ранней радиации млекопитающих // Палеонтологический журнал. 2003. № 1. С. 78—91
Татаринов Л. П. Териодонты СССР// Труды Палеонтологического института РАН. М.: Наука, 1974. Т. 143. 250 с.
Татаринов Л. П. Морфологическая эволюция териодонтов и общие проблемы филогенетики. М.: Наука, 1976. 258 с.
Gambaryan P. P., Kielan-Jaworowska Z. Masticatory musculature of Asian taeniolabidoid multitubrculate mammals // Acta Paleontologica Polonica.1995. V. 40. N 1. P. 45—108
Gambaryan P. P., Kielan-Jaworowska Z. Sprawling versus parasagittal stance in multituberculate mammals // Acta Paleontologica Polonica.1997. V. 42. N 1. P. 13—44
Hu Yaoming, Meng Jin, Wang Yuanqing, Li Chuankui. Large Mesozoic mammals fed on young dinosaurua // Nature. 2005. V. 433. P. 149—152
Jenkins F. A. Jr., Schaff Ch. R. The early Cretaceous mammal Gobiconodon (Mammalia, Triconodontia) from the Cloverly Formation in Montana. // J. Vertebrate Paleontology.1988. V. 8. N 1. P. 1—24
Kielan-Jaworowska Z., Gambaryan P. P. Postcranial anatomy and habitas of Asian multituberculate mammals // Fossils and Strata. 1994. N 36. 92 p.
Novacek M. J., Rougier G. W., Wible J. R., McKenna M. C., Dashzeveg D., Horovitz I. Epipubic bones in eutherian mammals from the Late Cretaceous of Mongolia // Nature. 1997. V. 389. P. 483—486
Hurum J. H., Presley R., Kielan-Jaworoeska Z. The middle ear in multituberculate mammals // Acta Paleontologica Polonica.1996. V. 41. N 3. P. 253—275
Rougier G. W., Wible J. R., Novacek M. J. Middle-ear ossicles of the Multituberculate Kryptobaatar from the Mongolian Late Cretaceous: implications for mammaliamorph relationships and the evolution of auditory apparatus // Novitates American Museum. 1996. N 3187. 43 p.
Исследования поддержаны грантом РФФИ №08-04-00483-а, программой Президиума РАН «Происхождение и эволюция биосферы» и программой ОБН РАН «История биоресурсных видов и сообществ млекопитающих как основа оценки их современного состояния и перспектив сохранения в будущем»
Источник: scfh.ru