Рисунок экосистемы озера схематичный
Рассмотрим экосистему небольшого естественного водоёма — озера.
Озёра располагаются в углублениях суши. Вода, текущая в реках, собирается в котловинах и остаётся там. Из некоторых озёр вода не вытекает — они называются бессточными. Как правило, такие озёра солёные. Из других озёр вода вытекает. Это сточные озёра.
Кто обитает в озёрах?
Стрелки связывают живые организмы в пищевые цепи, которые показывают превращения веществ «кормильцев» в вещества «едоков» и веществ «едоков» в вещества «мусорщиков». В этих превращениях энергия из пищи переходит от одних организмов к другим. Стрелка от «мусорщиков» к «кормильцам» замыкает круговорот.

А теперь давай познакомимся с обитателями озёр.
Некоторые растения-«кормильцы» очень малы. Это одноклеточные, плавающие в толще воды водоросли, образующие планктон.
|
| Растения в озере (элодея, кувшинка, рогоз, тростник) |
На дне растут большие растения, например водоросль хара. А могут быть и цветковые растения. Посмотри на знакомую тебе по аквариуму элодею.
Наконец, вблизи берегов видны многочисленные растения, которые растут в воде, а их листья и стебли расположены над водой. Это кувшинка, рогоз и тростник.
Масса одноклеточных водорослей привлекает внимание многочисленных «едоков». Среди этих «едоков» преобладают маленькие рачки — циклопы и дафнии. Они хорошо известны любителям аквариумов как корм для рыб.
|
| Озёрные «едоки» — рыбы и раки (щука, окунь, карась, плотва, речной рак) |
Рачками кормятся многие рыбы, в том числе и изображённые на рисунке карась и плотва. В свою очередь, этими рыбами нередко лакомятся хищные окунь и щука.
На дне водоёмов живут различные «мусорщики». К ним относятся всем известные раки. А кроме того, там множество микробов, которых нельзя увидеть без микроскопа.
Среди «мусорщиков» на дне можно встретить небольших красных червячков. Это мотыль — личинки комаров. Они кормятся на дне остатками умерших организмов. Материал с сайта http://worldofschool.ru
Посмотрим на первый рисунок, изображающий все связи живых организмов в озере. Обрати внимание: часть умерших «едоков» и «кормильцев» не попадает на обед к «мусорщикам». Они просто не справляются со своей работой. А это значит, что круговорот веществ в озере незамкнутый.
Откуда же берут «кормильцы» необходимые им минеральные вещества? Оказывается, из рек. Реки постоянно приносят всё новые и новые массы минеральных веществ, обеспечивая «кормильцев».
Если круговорот веществ не замкнут полностью, то условия должны постепенно меняться. Именно поэтому на месте озера рано или поздно (может быть, спустя тысячи и миллионы лет) появляется болото.
Источник: WorldOfSchool.ru
Важные абиотические факторы[править | править код]
Свет[править | править код]
Свет поставляет солнечную энергию, необходимую для управления процессом фотосинтеза, основным источником энергии стоячих систем (Бронмарк и Хэнсон, 2005). Количество полученного света зависит от комбинации нескольких факторов. Небольшие пруды могут возникнуть из-за затенения окружающими деревьями, а облачный покров может повлиять на наличие света во всех системах, независимо от их размера. Сезонные и суточные колебания также играют роль в легкости доступности, потому что чем меньше угол, под которым свет падает на воду, тем больше света теряется при отражении. Это известно как закон Бера (Гиллер и Малмквист, 1998). После того как свет проникает через поверхность, он также может быть разбросаны на частицы, взвешенных в толще воды. Это рассеяние снижает общее количество света с увеличением глубины (Мосс 1998, Калфф 2002). Озера разделены на световые и афотические зоны, предварительно принимающие свет и в дальнейшем полностью его рассеивающие, что делает его лишенным фотосинтетического потенциала (Бронмарк и Хэнсон, 2005). В соответствии с зональностью озера, считается, что пелагические и бентосные зоны лежат в пределах световой области, в то время как профундали в афотической зоне (Браун, 1987).
Температура[править | править код]
Температура является важным абиотическим фактором в стоячих экосистем, поскольку большая часть биоты пойкилотермна, где внутренняя температура тела определяется окружающей системой. Вода может нагреваться или охлаждаться посредством излучения с поверхности и теплопроводности из воздуха или окружающего субстрата (Гиллер и Малмквист, 1998). Мелкие водоемы часто имеют непрерывный градиент температуры — от теплых вод на поверхности до холодных вод на дне. Кроме того, флуктуации температуры могут быть очень большими в этих системах, как суточные, так и сезонные (Браун, 1987).
В больших озерах температурные режимы очень отличаются (Рис. 1). В регионах с умеренным климатом, например, при увеличении температуры воздуха, ледяной слой, сформированный на поверхности озера, распадается, оставляя воду при температуре приблизительно 4 ° С. Это температура, при которой вода имеет самую высокую плотность. По ходу сезона, более нагретый воздух разогревает поверхностные воды, что делает их менее плотными. Более глубокие воды остаются прохладными на ощупь и плотными из-за снижения интенсивности проникновения света. Когда начинается лето, два различных слоя с такой большой разницей температур между ними остаются стратифицированными. Нижняя холодная зона в озере называется гиполимнион.
рхняя теплая зона называется эпилимнион. Между этими зонами находится полоса быстрого изменения температуры, которая называется термоклин. В более холодный осенний сезон потери тепла на поверхности охлаждают и эпилимнион. Когда температура в двух зонах находится примерно равной, воды начинают снова перемешиваться, чтобы создать равномерную температуру, это событие называют переворотом озера. В зимнее время происходит обратная стратификация, воды вблизи поверхности охлаждаются, замерзают, а теплые, но плотные воды остаются вблизи дна. Устанавливается термоклин, и цикл повторяется (Браун, 1987, Бронмарк и Ханссон, 2005).
Ветер[править | править код]
В открытых системах ветер может создать турбулентные, спиральные поверхностные токи, называемые вращения Ленгмюра. Эти токи еще недостаточно хорошо изучены, но очевидно, что они включают в себя некоторое взаимодействие между горизонтальными поверхностными течениями и поверхностными гравитационными волнами. Видимым результатом этих вращений, который можно увидеть в любом озере, являются поверхность из линий, которые проходят параллельно направлению ветра. Плавучие частицы и мелкие организмы концентрируются во вспениваниях на поверхности, не плавучие объекты находятся в восходящем потоке между двумя вращениями. Объекты с нейтральной плавучестью, как правило, должны быть равномерно распределены в толще воды (Калфф, 2002; Бронмарк и Ханссон, 2005). Эта турбулентность перемешивает питательные вещества в толще воды, что делает её важным для многих пелагических видов, однако её влияние на донные и придонные организмы является минимальной (Калфф, 2002). Степень питательной циркуляции зависит от системы факторов, таких как сила и продолжительность ветра, а также глубины и производительности озера или бассейна.
Химия[править | править код]
Кислород необходим для дыхания организмов. Количество кислорода, присутствующего в стоячей воде, зависит от: 1) области прозрачной воды, имеющей доступ к воздуху, 2) циркуляции воды в системе и 3) количества кислорода, который образуется и используется организмами (Браун, 1987). В мелководных, богатых растительностью бассейнах могут быть большие колебания большой концентрации кислорода, происходящие из-за фотосинтеза в течение дня и очень низких его значений в ночь, когда дыхание является доминирующим процессом первичных производителей. Тепловое расслоение в больших системах также может влиять на количество кислорода, присутствующего в различных зонах. Эпилимнион является зоной, богатой кислородом, так как он быстро циркулирует, получая кислород из контакта с воздухом. Гиполимнион, в свою очередь, циркулирует очень медленно и не имеет атмосферного контакта. Кроме того, в гиполимнионе меньше зеленых растений, так что меньше кислорода высвобождается в результате фотосинтеза. Весной и осенью, когда эпилимнион и гиполимнион смешиваются, кислород распределяется более равномерно в системе (Браун, 1987). Низкий уровень кислорода характерен профундали из-за накопления разлагающихся растительности и животных остатков, которые падают «дождями» вниз из пелагических и бентальных зон и невозможности поддержки первичными производителями (Браун, 1987).
Фосфор важен для всех организмов, потому что это — компонент ДНК и РНК и вовлечен в метаболизм клетки как компонент ATP и АВТОМАТИЧЕСКОЙ ОБРАБОТКИ. Кроме того, фосфор в больших количествах не найден в пресноводных системах, ограничившись фотосинтезом в основных производителях, делая его главным детерминантом непроточного системного производства. Цикл фосфора сложен, но модель, обрисованная в общих чертах ниже, описывает основные положения. Фосфор, главным образом, входит в водоем или озеро через сток водораздела или атмосферным осаждением. После входа в систему реактивная форма фосфора обычно поднимается морскими водорослями и макрофитами, которые выпускают нереактивный состав фосфора как побочный продукт фотосинтеза. Этот фосфор может дрейфовать вниз и стать частью бентического или профундического осадка, или может быть повторно минерализован к реактивной форме микробами в водяном столбе. Точно так же нереактивный фосфор в осадке может быть повторно минерализован в реактивную форму (Бренмарк и Ханссон, 2005). Отложения обычно более богаты фосфором, чем вода озера, однако необходимо учитывать, что у этого питательного вещества может быть долгое время расположения там, прежде чем оно будет повторно минерализовано и введено в систему (Калфф, 2002).
Непроточная биотическая система[править | править код]
Бактерии[править | править код]
Бактерии присутствуют во всех областях непроточных вод. Свободно живущие формы находятся в разлагающихся органических материалах, биопленка на поверхностях скал и заводов подвешены в водяном столбе и в отложениях бентических и профундических зон. Также другие формы находятся в кишках непроточных животных как паразиты или в качестве сотрапезника.[3] Бактерии играют важную роль в системном метаболизме посредством переработки питательных веществ,[2] который обсужден в секции трофических отношений.
Первичные производители[править | править код]
Морские водоросли, включая фитопланктон и перифитон, являются основными фотосинтезаторами в водоемах и озерах. Многие разновидности имеют более высокую плотность, чем вода, что заставляет их снизиться и остаться в бентосе. Чтобы бороться с этим, фитопланктон разработал механизм изменения плотности путём формирования вакуоли и газовых пузырьков, или изменяя их форму, чтобы вызвать сопротивление, замедляя спуск. Очень сложная адаптация, используемая небольшим количеством разновидностей, является кнутом, подобным хвосту, который может установить вертикальное положение и позволить движение в любом направлении.[2] Фитопланктон может также поддерживать своё присутствие в водяном столбе, будучи распространенным во вращениях Лангмюра.[3] Морские водоросли перифит, с другой стороны, присоединены к основанию. В озерах и водоемах они могут покрыть все бентические поверхности. Оба типа планктона важны как источники пищи и как кислородные поставщики.[2]
Водяные растения, живущие и в бентических, и в морских зонах могут быть сгруппированы согласно их манере роста: 1) всплывающий = прикрепленный к дну, но с листьями и цветами, расположенными на поверхности, 2) с плавающими листьями = прикрепленный к дну, но с плавающими листьями, 3) подводный = растущий ниже поверхности и 4) свободно плавающие макрофиты = не внедренные в основании и плавающие на поверхности.[1] Эти различные формы макрофитов обычно находятся в различных областях бентической зоны вместе со всплывающей растительностью, близкой к береговой линии, тогда как макрофиты с листом плавания сопровождаются подводной растительностью. Свободное плавание макрофитов может происходить где угодно на поверхности системы.[2]
Водяные растения более подвижны, чем их земные коллеги, потому что вода имеет более высокую плотность, чем воздух. Это делает структурную жесткость растений неважной в озерах и водоемах (кроме воздушных стеблей и листьев).
ким образом, листья и стебли большинства водяных растений используют меньше энергии, чтобы построить и поддерживать древесную ткань, вместо этого инвестируя избыток этой энергию в быстрый рост.[1] Чтобы бороться с давлением, вызываемым ветром и волнами, растения должны быть и гибкими, и жесткими. Свет, глубина воды и типы основания — наиболее важные факторы, управляющие распространением затопленных водяных растений.[4] Макрофиты — источники еды, кислорода и среда обитания в бентической зоне, но не могут проникнуть через глубины афотической зоны и, следовательно, не найдены там.[1][5]
Беспозвоночные[править | править код]
Зоопланктон — крошечные животные, временно подвешенные в водяном столбе. Как фитопланктон, эти виды разработали механизмы, которые препятствуют их снижению в более глубокие воды, включая вызывающие сопротивление формы тела и активно щелкающие придатки, такие как антенны или шипы.[1] Существование в водяном столбе может иметь преимущества с точки зрения питания, но в этой зоне отсутствуют листья зоопланктона refugia, уязвимого для хищничества. В ответ некоторые разновидности, особенно вид Дафний совершают ежедневные вертикальные миграции в водяном столбе, пассивно снижаясь к более темным, более низким глубинам в течение дня и активно двигаясь к поверхности в течение ночи. Кроме того, поскольку условия в непроточной системе могут быть довольно переменными в различные сезоны, зоопланктон имеет способность переключиться с откладывания обычных яиц к покоящимся яйцам, когда отсутствует еда, температура падает ниже 2 °C, или если высоко изобилие хищника. У этих покоящихся яиц есть состояние покоя или период дремоты, который позволяет зоопланктону дожидаться условий, которые более благоприятны для выживания, когда они наконец вылупятся.[6] Беспозвоночные, которые населяют бентическую зону, преимущественно доминируют над маленькими разновидностями и являются видами, богатыми по сравнению с зоопланктоном открытой воды. Они включают Ракообразных (например, крабы, речной рак и креветки), моллюски (например, моллюски и улитки), и многочисленные типы насекомых.[2] Эти организмы главным образом найдены в областях роста макрофитов, богатые ресурсами, очень насыщенная кислородом вода и самая теплая часть экосистемы. Структурно разные «кровати» макрофитов — важные места для накопления органического вещества, предоставляющие идеальную область для колонизации. Отложения и растения также предлагают большую защиту от хищных рыб.[3]
Очень немного беспозвоночных в состоянии населять холодную, темную и бедную кислородом профундическую зону. Те, которые могут, часто красного цвета из-за присутствия больших количеств гемоглобина, который значительно увеличивает количество кислорода, который несут к клеткам.[1] Поскольку концентрация кислорода в этой зоне низкая, большинство видов строит тоннели или занимает его у тех, кто может перетащить его сквозь толщу воды с минимальными потерями энергии.[1]
Рыбы и другие позвоночные[править | править код]
У рыб есть диапазон физиологической терпимости, который зависит от того, к каким разновидностям они принадлежат. У них различные летальные температуры, требования растворенного кислорода и необходимость в метании икры, которые основаны на их уровнях, активности и поведении. Поскольку рыбы очень мобильны, они в состоянии менять неподходящие абиотические факторы в одной зоне, просто двигаясь к другой. Едок детрита в профундической зоне, например, который попал в пониженную концентрацию кислорода, может питаться ближе к бентической зоне. Рыба может также изменять своё место жительства во время различных периодов её жизненного цикла: вначале оседает в гнезде, затем перемещается в слабую бентическую зону, чтобы развиться в защищенной окружающей среде с пищевыми ресурсами, и, наконец, в морскую зону, как взрослый.
Также другие позвоночные таксоны населяют непроточные системы. Они включают амфибий (например, саламандры и лягушки), рептилий (например, змеи, черепахи и аллигаторы), и большое количество разновидностей водоплавающей птицы.[5] Большинство этих позвоночных животных тратит часть своего времени в земных средах обитания, и, таким образом, непосредственно не затронуты абиотическими факторами в озере или водоеме. Многие виды рыбы важны как потребители и разновидности добычи более крупным (упомянутым выше) позвоночным животным.
Трофические взаимоотношения[править | править код]
Первичные производители[править | править код]
Непроточные системы получают большую часть своей энергии от фотосинтеза, который выполняется водяными растениями и морскими водорослями. Этот коренной процесс включает в себя комбинирование углекислого газа, воды и солнечной энергии, чтобы произвести углеводы и растворенный кислород. В озере или водоеме, потенциальный темп фотосинтеза обычно уменьшается с глубиной из-за ослабления интенсивности света. Однако фотосинтез часто протекает, главным образом, в нескольких миллиметрах от поверхности, вероятно, из-за препятствования ему ультрафиолетовыми лучами. Точная глубина и фотосинтетические темпы этой кривой — определенная система и зависит от: 1) всей биомассы фотосинтезирующих клеток, 2) количества материалов, уменьшающих лучи и 3) изобилия и частотного диапазона пигментов, абсорбирующих свет (т.е. хлорофиллы) в фотосинтезировании клеток.[5] Энергия, созданная этими первичными производителями, важна для сообщества, потому что она передается более высоким трофическим уровням через потребление.
Бактерии[править | править код]
Подавляющее большинство бактерий в озерах и водоемах получает энергию, перерабатывая останки животных и растительности. В морской зоне мертвая рыба и случайный аллохтонный опад — примеры макрочастиц грубого органического вещества (CPOM>1 мм). Бактерии перерабатывают их в микрочастицы органического вещества (FPOM<1 мм) и затем далее в удобоваримые питательные вещества. Маленькие организмы, такие как планктон, также характеризуются как FPOM. Очень низкие концентрации питательных веществ потеряны во время разложения, потому что бактерии используют их, чтобы построить их собственную биомассу. Бактерии, однако, поглощаются простейшими, которые, в свою очередь, потребляются зоопланктоном, и затем выше по трофическим уровням. Питательные вещества, включая те, которые содержат углерод и фосфор, повторно вводятся в водяной столб на любом из уровней этой пищевой цепи через выделения или смерть организма, делая их доступными снова для бактерий. Этот цикл регенерации известен как микробная петля и является ключевым компонентом непроточных пищевых сетей.[2]
Разложение органических материалов может продолжиться в бентических и придонных зонах, если останки проваливаются по водяному столбу, прежде чем быть полностью переваренными морскими бактериями. В самом большом изобилии бактерии найдены в отложениях, где они, как правило, в 2—1000 раз более распространены, чем в водяном столбе.[6]
Придонные беспозвоночные[править | править код]
У придонных беспозвоночных из-за их высокого богатства разновидностей есть много методов захвата добычи. Фильтрующие создают ток через сифоны или бьющиеся ресницы, чтобы протянуть воду и её пищевое содержание к себе для процеживания. Травоядные используют приспособления для очистки, измельчения и рубки, чтобы питаться морскими водорослями и макрофитами. Собиратели прочесывают отложения, выбирая определенные частицы с хищными придатками. Беспозвоночные, питающиеся отложениями, потребляют осадок без разбора, переваривая любой органический материал, который он содержит. Наконец, некоторые беспозвоночные хищники захватывают и поглощают живых животных.[2][7] Придонная зона является родиной уникальной группы фильтрующих, которые используют слабые движения тела, чтобы протянуть ток через норы, которые они создали в отложениях. Этот способ кормления требует наименьшего количества движений, позволяя этим разновидностям сохранить энергию.[1] Небольшое количество беспозвоночных таксонов является хищниками в придонной зоне. Эти разновидности, вероятно, из других областей и прибывают в эти глубины только питаться. Подавляющее большинство беспозвоночных в этой зоне питаются отложениями, получая энергию из окружающих остатков.[7]
Рыбы[править | править код]
Размер рыбы, подвижность и чувствительные способности позволяют им иметь обширную базу добычи, покрывая многократные области зонирования. Как беспозвоночные, пищевые привычки рыбы могут быть категоризированы в гильдии. В морской зоне травоядные животные пасутся на придонных макрофитах или фитопланктоне, забираемых из водяного столба. Плотоядные животные включают рыб, которые питаются зоопланктоном в водяном столбе (zooplanktivores), насекомых на поверхности воды, на бентических структурах, или в осадке (насекомоядные), и тех, которые питаются другой рыбой (хищники). Рыб, которые потребляют остатки и получают энергию, перерабатывая этот органический материал, называют детритофагами. Всеядные существа потребляют разнообразную добычу, включая растительный, животный и органический материал. Наконец, члены паразитной гильдии потребляют пищу от различных хозяев, обычно других рыб или больших позвоночных животных.[2] Таксоны рыбы гибки в своих питательных ролях, изменяя свои диеты в связи с доступностью добычи и условиями окружающей среды. Много разновидностей также подвергаются диетическим изменениям по мере того, как они развиваются. Поэтому вполне вероятно, что единственная рыба занимает разнообразные питательные гильдии на протяжении всей своей жизни.[8]
Пищевые цепи непроточных водоемов[править | править код]
Как отмечено в предыдущих секциях, непроточная биоматерия связана в сложной паутине трофических отношений. Эти организмы, как можно полагать, свободно связаны с определенными трофическими группами (например, основные производители, травоядные животные, первичные плотоядные животные, вторичные плотоядные животные, и т.д.). Ученые развили несколько теорий, чтобы понять механизмы, которые управляют изобилием и разнообразием в пределах этих групп. Очень похоже, нисходящие процессы говорят о том, что изобилие таксонов добычи зависит от действий потребителей на высших трофических уровнях. Как правило, эти процессы работают только между двумя трофическими уровнями без эффекта на другие. В некоторых случаях, однако, водные системы испытывают трофический каскад; например, это может происходить, если первичные производители испытывают меньшее давление от травоядных животных, потому что эти травоядные животные подавлены плотоядными животными. Восходящие процессы функционируют, когда изобилие или разнообразие членов более высоких трофических уровней зависит от доступности или качества ресурсов более низких уровней. Наконец, объединенная теория регулирования, bottom-up&top-down, объединяет предсказанные влияния доступности ресурса и потребителей. Это предсказывает, что трофические уровни, близкие к самым низким трофическим уровням, будут больше всего под влиянием восходящих сил, в то время как нисходящие эффекты должны быть самыми сильными на высших уровнях.[2]
Разнообразие и образцы сообществ[править | править код]
Богатство на локальном уровне[править | править код]
Биоразнообразие непроточной системы увеличивается с площадью поверхности озера или водоема. Это заключается в более высокой вероятности нахождения наземных разновидностей в большой системы. Кроме того, поскольку у больших систем, как правило, более многочисленное население, шанс исчезновения видов уменьшен.[9] Дополнительные факторы, включая температурный режим, pH-фактор, доступность питания, сложность среды обитания, процент видообразования, соревнование и хищничество связаны с числом разновидностей, представленных в пределах систем.[2][4]
Наследование образцов в планктонных сообществах — модель PEG[править | править код]
Фитопланктон и сообщества зоопланктона в системах озера подвергаются сезонной последовательности относительно питательной доступности, хищничества и соревнования. Соммер и др.[10] описал эти образцы как часть модели Plankton Ecology Group (PEG) с 24 заявлениями, построенными из анализа многочисленных систем. Следующее включает в себя подмножество этих заявлений, как объяснил Бренмарк и Ханссон[2], последовательность иллюстрирования через единственный сезонный цикл:
Зима: 1. Увеличенная питательная и световая доступность приводит к быстрому росту фитопланктона к концу зимы. Доминирующие разновидности, такие как диатомовые водоросли, являются маленькими и имеют быстрые возможности роста. 2. Этот планктон потребляется зоопланктоном, которые становятся доминирующими таксонами планктона.
Весна: 3. Происходит фаза очищения воды, поскольку население фитопланктона истощается из-за увеличенного хищничества растущим числом зоопланктона.
Лето: 4. Изобилие зоопланктона уменьшается в результате уменьшенной добычи фитопланктона и увеличенного хищничества молодью. 5. С увеличенной питательной доступностью и уменьшенным хищничеством от зоопланктона, развивается разнообразие сообщества фитопланктона. 6. В то время как лето продолжается, питательные вещества становятся исчерпанными в предсказуемом порядке: фосфор, кварц и затем азот. Изобилие различных разновидностей фитопланктона варьируется относительно их биологической потребности в этих питательных веществах. 7. Зоопланктон маленького размера становится доминирующим типом зоопланктона, потому что они менее уязвимы, чтобы быть пойманными хищниками.
Спад: 8. Хищничество рыб уменьшено, понижаются температуры, и зоопланктон всех размеров увеличивается численно.
Зима: 9. Низкие температуры и уменьшенная легкодоступность приводят к более низким показателям основного производства и уменьшению численности фитопланктона. 10. Воспроизводство в зоопланктоне уменьшается из-за понижений температуры и уменьшения числа добычи.
Модель PEG представляет идеализированную версию этой последовательности, в то время как естественные системы известны их изменением.[2]
Широтные модели[править | править код]
Есть хорошо зарегистрированный глобальный пример, который коррелирует уменьшающееся разнообразие растений и животных с увеличивающейся широтой, то есть меньше разновидностей при движении к полюсам. Причина этого примера является одной из самых больших загадок для экологов сегодня. Теории для её объяснения включают энергетическую доступность, климатическую изменчивость, волнение, конкуренция, и т.д.[2] Несмотря на этот глобальный градиент разнообразия, этот пример может быть неубедительным для пресноводных систем по сравнению с глобальными морскими и земными системами.[11] Это точно может быть связано, поскольку Хиллебранд и Азовский[12] нашли, что меньшие организмы (протозоа и планктон) не сильно следовали ожидаемой тенденции, в то время как большие разновидности (позвоночные животные) следовали. Они приписывали это лучшей способности меньших организмов к распространению, что привело к высокому глобальному заселению.[2]
Природные циклы озера[править | править код]
Формирование озер[править | править код]
Озера могут быть сформированы различными путями, но наиболее распространенные кратко обсуждены ниже. Старейшие и крупнейшие системы являются результатом тектонической деятельности. Щелевые озера в Африке, например, результат сейсмической активности вдоль линии разделения двух тектонических плит. Сформированные льдом озера создавались, когда ледники отступали, оставляя позади провалы в формах ландшафта, которые тогда были заполнены водой. Наконец, озера-старицы являются речными, появляясь, когда блуждающий речной изгиб отсекается от главного канала.[2]
Естественное вымирание[править | править код]
Все озера и водоемы получают осадки. Так как эти системы действительно не расширяются, логично предположить, что они будут все более и более становиться мелкими, в конечном счете становясь заболоченными местами или земной растительностью. Продолжительность этого процесса должна зависеть от комбинации уровня отложения осадка и глубины. Мосс[5] дает пример Озера Танганьика, которое достигает глубины 1500 м и имеет ставку отложения осадка 0.5 мм/год, предполагая, что при отложении осадка без вмешательства антропогенных факторов эта система должна исчезнуть приблизительно через 3 миллиона лет.[5]
Вмешательство человека[править | править код]
Подкисление[править | править код]
Двуокись серы и окислы азота естественным путём исходят из вулканов, органических соединений в почве, заболоченных мест и морских систем, но большинство этих составов образуются при сгорании угля, нефти, бензина и плавления руд, содержащих серу.[3] Эти вещества распадаются во влаге атмосферы и входят в непроточные системы с помощью кислотного дождя.[1] У озер и водоемов, которые содержат основу, богатую карбонатами, есть естественный буфер, не приводящий ни к какому изменению pH-фактора. Системы без этой основы, однако, очень чувствительны к кислотным добавкам, потому что у них низкая мощность нейтрализации, приводящая к снижениям pH фактора даже из-за маленьких порций кислоты.[3] При рН≈5-6 уничтожаются многие виды водорослей и сокращается биомасса, что приводит к увеличению водной прозрачности – характерная особенность окисленных озер. В то время как pH-фактор становится ниже, вся фауна уменьшает своё разнообразной. Самой значительной особенностью является разрушение воспроизводства рыбы. Таким образом, популяцию в конечном счете составляют из немногие, старые особи, которые в конечном счете умирают и оставляют системы без рыб.[2][3] Кислотные дожди были особенно вредны для озер в Скандинавии, западной Шотландии, западном Уэльсе и северных восточных Соединенных Штатах.
Эвтрофикация[править | править код]
Эвтрофицированные системы содержат высокую концентрацию фосфора (~30 +µg/L), азота (~1500 +µg/L), или обоих сразу.[2] Фосфор входит в непроточные воды из сточных вод, от необработанных сточных вод или от стока с сельхозугодий. Азот главным образом прибывает из сельскохозяйственных удобрений в стоках или выщелачивания и последующего стекания грунтовой воды. Это увеличение питательных веществ, требуемых для первичных производителей, приводит к крупному увеличению роста фитопланктона, который называют цветением планктона. Это цветение уменьшает водную прозрачность, приводя к потере подводных растений. Вследствие сокращения структуры среды обитания возникают негативные воздействия на виды, которые используют его для размножения, созревания и жизни. Кроме того, большое количество коротко живущего фитопланктона приводит к большому количеству мертвой биомассы, осаждаемой на дне.[5] Бактериям нужно большое количество кислорода, чтобы переработать этот материал, что приводит к уменьшению концентрации кислорода в воде. Это особенно ярко заметно в стратифицированных озерах, когда термоклин предотвращает перемешивание богатой кислородом воды на поверхности с более низкими уровнями. Низкие или бескислородные условия уничтожают существование многих таксонов, которые физиологически не терпимы к этим условиям.[2]
Агрессивные виды[править | править код]
Агрессивные виды были введены в непроточные системы посредством целенаправленных действий (например, проекты снабжения и через продовольственные виды), а также неумышленных событий (например, в балласте воды). Эти организмы могут затронуть местных жителей, конкурируя за добычу или среду обитания, хищничество, произвести изменение среды обитания, гибридизацию или привнесение вредных болезней и паразитов.[13] Относительно аборигенного вида, захватчики могут вызвать изменения в размере и структуре возраста, распределении, плотности, приросте населения, и могут даже вести население к исчезновению.[2] Примеры знаменитых захватчиков непроточных систем включают дрейссену и морскую миногу в Великих озерах.
См. также[править | править код]
- United States Environmental Protection Agency – Great Lakes Ecosystems
- United States Environmental Protection Agency – Limnology Primer (PDF file)
- Freshwater environmental quality parameters
- Limnology
- Lake aeration
- Man-made lentic water bodies of Maharashtra
Источник: ru.wikipedia.org
Источником энергии для озера служит солнечный свет. При прохождении света через воду некоторая его часть поглощается планктоном и используется в процессе фотосинтеза; остальной свет поглощает вода, так что с увеличением глубины освещенность уменьшается. В глубоких озерах существует некий компенсационный уровень, т.е. глубина, на которую проникает столько света, что зеленые растения едва могут поддерживать свое существование: фотосинтез у таких растений (образование сложных органических веществ и кислорода) в точности уравновешивает их дыхание (расход пищи и кислорода).
Рисунок 1: Структура озёрной экосистемы. Показаны
некоторые организмы, обитающие в различных частях озера.
Глубина, на которой находится компенсационный уровень, зависит от прозрачности воды. Выше этой глубины растения выделяют больше кислорода, чем потребляют, так что избыточный кислород могут использовать другие организмы; ниже этой глубины фотосинтез не может обеспечить дыхание, поэтому организмам доступен лишь кислород, поступающий с водой из более поверхностных слоев озера.
Укореняющиеся водные растения, такие, как водяные лилии и камыши, растут на мелководных участках озера. Среди этих растений находят себе убежище и пищу рыбы, головастики, насекомые и другие членистоногие, моллюски и черви. На открытой поверхности озера обитают свободно плавающие растения, которым необходим свет, и животные, которым необходимо обилие кислорода, — многие рыбы и мелкие членистоногие (рисунок 1).
На большей глубине, где меньше кислорода и недостаточно света для протекания фотосинтеза, главным источником энергии служат мертвые растения и животные, опускающиеся сверху. Организмы-редуценты, рыбы и беспозвоночные, способные переносить низкое содержание кислорода, питаются этими остатками, а также поедают друг друга.
Сильное влияние на жизнь озера оказывает температура. Вода характеризуется одной уникальной особенностью: при 4°С она обладает наибольшей плотностью. Поэтому слои воды, имеющие температуру 4°С, опускаются ниже слоев, имеющих более высокую или более низкую температуру. В результате этого зимой вода с температурой ниже 4°С поднимается вверх и, охлаждаясь до 0°С, замерзает, покрывая озеро слоем льда.
Рисунок 2: Температура воды в разное время года на
разной глубине в озере, находящемся в умеренной зоне.
Этот слой служит изоляцией, предохраняя лежащую под ним воду от полного замерзания, так что обитатели озера выживают под слоем льда в воде, которая в течение всей зимы сохраняет температуру от 0 до 4°С (рисунок 2). Весной солнце растапливает лед и нагревает поверхностные слои воды, которые с приближением их температуры к 4°С погружаются вниз, заставляя лежащую под ними более холодную воду подниматься вверх. Этому весеннему перемешиванию способствуют ветер и волны. Такое перемешивание имеет важное значение, потому что вода, поднимающаяся вверх, приносит с собой питательные вещества, содержащиеся в донных отложениях, и обитающие у поверхности фотосинтезирующие организмы могут использовать их. В свою очередь вода, перемещающаяся из поверхностных слоев озера в придонные, приносит кислород редуцентам и другим обитателям дна озера.
Рисунок 3: Процесс исчезновения олиготрофного озера.
А. Олиготрофное озеро.Б. По мере увеличения притока в озеро питательных веществ и осадочных материалов его продуктивность возрастает и оно становится эвтрофным.
В. В дальнейшем озеро превращается в болото, и покрывается растительностью.
Г. В конце концов это место превращается в сушу. Обычно эта последовательность событий занимает несколько тысяч лет даже для таких мелководных озер, как озеро Эри. Глубина некоторых олиготрофных озер, например озера Тахо, так велика, что для них процесс этот растянулся бы на неограниченно долгое время, если бы эвтрофикация не ускорялась загрязнением.
Если плавая летом в озере с приятной и теплой водой мы опустим ноги вниз, то ощутим, что вода там гораздо холоднее. Это происходит потому, что нагретый солнцем поверхностный слой воды остается на месте, не смешиваясь с лежащей под ним более плотной и холодной водой, температура которой в глубоком озере остается на уровне 4°С. Граница между теплой и холодной водой называется термоклиной и летом она постепенно опускается все ниже и ниже. Осенью термоклина начинает вновь подниматься, по мере того как верхний слой воды охлаждается, что в конце концов приводит к осеннему перемешиванию. Рыбаки, занимающиеся ловлей форели, знают, что весна и осень-это то время, когда можно добыть форель в поднимающейся вверх холодной воде. Форели нужна вода, богатая кислородом, а поскольку в холодной воде растворяется больше кислорода, чем в теплой, форель проводит лето на большей глубине, где вода холоднее.
По продуктивности озера можно разделить на две группы. Эвтрофные («многокормные») озера — относительно мелководные, богаты элементами питания и бедны кислородом. В отличие от них олиготрофные («малокормные») озера обычно более глубокие, с более крутыми берегами и менее богаты элементами питания; вода в них прозрачная и до самого дна может содержать много кислорода. При нормальном течении событий озеро медленно наполняется осадочным материалом и мертвым органическим веществом и постепенно эвтрофицируется. В конце концов оно превращается в верховое или низинное болото, а затем и в сушу (рисунок 3). В случае глубокого олиготрофного озера этот процесс может тянуться миллионы лет.
Одним из последствий загрязнения среды, вызванного человеком, является ускорение процесса эвтрофикации. На первый взгляд повышение продуктивности может показаться желательным, однако, как мы увидим ниже, это не так. Олиготрофные озера с чистой водой и рыбой гораздо более привлекательны и полезны человеку, чем эвтрофные озера, заросшие водорослями и сорняками и населенные карпом и мелкими представителями карповых.
В природные экосистемы весь фосфор изначально поступает из горных пород, а поскольку многие экосистемы расположены на породах, содержащих мало фосфора, их продуктивность ограничивается недостатком этого элемента.
Люди непрерывно вносят в озера фосфор вместе со сточными водами, детергентами или удобрениями, стекающими с обрабатываемых земель. Это повышает продуктивность озера и ускоряет его эвтрофикацию, изменяя характер озера.
Самый явный симптом эвтрофикации — помутнение воды, создаваемое усиленным размножением планктона. Другой признак-уменьшение численности таких рыб, как форель, любящих чистую воду, богатую кислородом. Загрязняемое элементами питания озеро проходит через несколько стадий, прежде чем достигнет хорошо знакомого всем состояния с вонючей тиной на дне и берегами, усеянными погибшей рыбой. Вначале загрязнение элементами питания способствует «цветению», или необычайно сильным взрывам численности популяции, водорослей. Некоторые водоросли выделяют токсины, отравляющие рыбу и делающие воду непригодной для питья. При отмирании водорослей бактерии разлагают их ткани и используют большую часть кислорода. В иле, лишенном кислорода, разложение мертвых растений некоторыми бактериями сопровождается выделением сероводорода (газа с запахом тухлых яиц). В «здоровом» озере обитающие на дне фотосинтезирующие бактерии используют этот сероводород по мере его образования. Однако при большом содержании в озере удобрений или сточных вод у его поверхности появляется так много водорослей и бактерий, что до дна доходит очень мало света и фотосинтезирующие серные бактерии растут плохо. Они не могут использовать весь образующийся сероводород, придающий загрязненному озеру его специфический запах.
Процесс эвтрофикации нельзя устранить, но его можно замедлить и временно повернуть в обратную сторону. Если прекратить сбрасывание в озеро сточных вод, то водоросли, питающиеся за счет этих вод, погибнут и опустятся на дно, причем замещения их происходить не будет. Вода станет более прозрачной, обогатится кислородом и популяции рыб восстановятся.
Такое загрязнение возникает в результате внесения в воду дополнительного тепла, например в тех случаях, когда электростанция использует озерную воду для охлаждения турбин. Поскольку при более высокой температуре рост растений ускоряется, при тепловом загрязнении продуктивность озера возрастает и, так же как и при загрязнении биогенными элементами, эвтрофикация ускоряется. Помимо этого повышение температуры воды может задержать и даже подавить размножение, развитие или рост некоторых рыб и беспозвоночных; при достаточно высокой температуре организмы просто гибнут. Неестественное подогревание воды может также увеличить промежуток между весенним и осенним перемешиваниями, нарушая нормальные сроки пополнения запасов элементов питания у поверхности воды и запасов кислорода вблизи дна.
Источник: info.sotvorenie.kiev.ua
Источники возникновения
На Земле возникали озера в результате тектонических сдвигов породы, отступления ледников при таянии или изменения русел рек. К ним можно относить пруды, более мелкие водные образования. Общее у них, то, что это замкнутые экосистемы с тенденцией к исчезновению.
Неважно, является водоем сточным, то есть из которого вытекает вода, или бессточным, экосистема озера постепенно будет трансформироваться в сторону преобладания в ней растительного над животным миром. Затем превратиться в болото и, в конце концов, высохнет и исчезнет. Быстрота такого превращения зависит лишь от величины и глубины водного объекта.
Структура системы и основные факторы влияния
Экосистема озера представляет собой видовую совокупность, существующую в границах водного объекта, и взаимодействующая между собой. Трофическая цепь типичная и состоит из продуцентов – растений и водорослей, консументов – рыбы, рептилии, водоплавающей птицы, некоторые видов животных, а также редуцентов – бактерий, червей и ракообразных.
Схематическая иллюстрация экосистемы озера.
Соленая в озере вода или пресная, влияет только на видовую структуру, в которой преобладают живые организмы, приспособленные к существованию в воде с большим или меньшим содержанием соли.
Основными факторами, влияющими на систему, являются солнце, температура воды и количество содержащегося в ней кислорода.
Главным и определяющим из них – солнце. Вступая во взаимодействие с водой, солнечная энергия изменяет, а именно повышает, температуру последней. Это, в свою очередь, влияет на процесс фотосинтеза, то есть производства кислорода, его содержание и растворимость в воде.
По количеству поступающей солнечной энергии, водную массу озера можно разделить на горизонтальные слои или пласты.
В летний период верхний слой получает максимальное количество солнечной энергии. Он нагревается. Продуценты активно перерабатывают солнечную энергию в кислород. Фауна в верхнем слое играет роль консументов. Это в основном водоплавающие животные и птицы, рептилии, некоторые виды рыб и насекомые.
Следующий пласт воды играет «заградительную» функцию между разными температурными слоями, расположенными над и под ним. Этот слой с максимальной плотностью воды, которая возникает, когда температура ее +4°С. Он сдерживает перемешивание слоев воды озера. Обычно перемешивание происходит весной и осенью. В результате чего происходит обмен кислородом и питательными веществами.
Солнечный свет, доходя до придонного слоя, сильно рассеивается. На дно попадают остатки живых организмов и отходы их жизнедеятельности. Придонный слой населяют редуценты – раки, черви, личинки насекомых, бактерии и микроорганизмы. Очень редки рыбы. Главная их функция переработка органических отходов. Последний этап пищевой цепи, перед началом нового.
На этом этапе и происходит тот сбой, который, в конечном счете ведет к исчезновению озера. Условия существования не позволяют справиться с переработкой отходов полностью. А верхний слой, подпитанный во время перемешивания, увеличивает биомассу. Отходы увеличиваются, а остатки накапливаются. Они превращаются в ил, а затем в торф. Озеро начинает мелеть и исчезать.
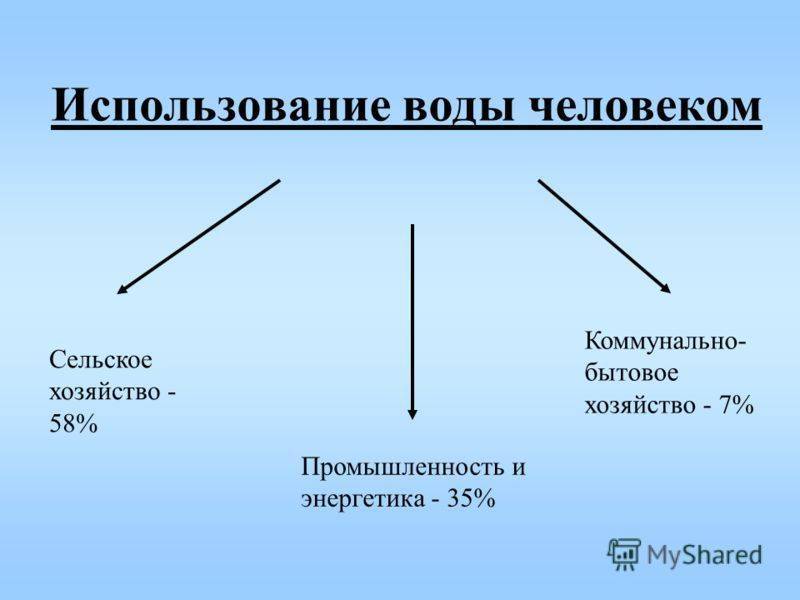
Использование человеком
Использование озера человеком можно описать предельно коротко. Человек берет из него воду и пищу, и возвращает неочищенную воду и отходы.
Перед тем как полностью исчезнуть, озеро превращается в болото. Донный ил становится торфом. Торф обладает способностью сохранять влагу. Накапливая ее в период таяния снегов или дождей, он затем отдает ее ручьям и тем поддерживают уровень воды в больших водоемах и в грунтовых водах. Человек добывая торф, как природное топливо или удобрение, проводя мелиоративные работы и осушая болота, изменяет водный режим региона со всеми вытекающими отсюда последствиями.
Экосистема озера не содержит фосфор, азот и другие вещества, стимулирующие рост растений. Сточные воды промышленных предприятий, сбросы канализационных систем городов, неочищенные бытовые стоки и, главное, вода, сходящая после ливней и таяния снегов с земель, используемых под сельскохозяйственные нужды, содержит эти вещества. А они ускоряют рост и увеличивают количество биомассы, особенно сине-зелёных водорослей.
Тот же эффект при сбросе теплой воды, после охлаждения ею оборудования электростанций. Повышение температуры воды в результате таких сбросов ускоряет рост все тех же водорослей и других растений. Если температура будет слишком велика, то животный мир может вообще погибнуть или произойти сбой в его репродуктивной системе.
Еще одна форма использования человеком экосистемы озера – это привнесение в нее живых организмов, ей несвойственных. Иногда это может произойти случайно. Но бывает, что это делается преднамеренно, с целью разведения полезных для человека видов рыб, моллюсков, беспозвоночных и тому подобное.
Эти организмы ведут себя агрессивно по отношению к местным видам флоры и фауны. А с учетом стимулирования их роста и развития человеком, то природная биосистема начинает подвергаться существенным изменениям. Происходит дисбаланс, который может привести к ее полной гибели. Примером могут служить Великие озера в Америке.
Вам будет интересно посмотреть фото и картинки экосистемы озера.
Посмотрите видео: Красивые фото озер, рек и морей.
Источник: ecology-of.ru