Общая формула фотосинтеза
Фотосинтез представляет собой высокоэффективный процесс запасания энергии видимого света и трансформирования ее в энергию химических связей биологических молекул. Осуществляется данный процесс в клетках растений, водорослей и фототрофных бактерий, причем у большинства перечисленных организмов фотосинтез сопровождается использованием в качестве донора электронов воды и выделением молекулярного кислорода (уравнение 13.1). И только у аноксигенных бактерий, относящихся к классу Anoxyphotobacteria , вместо воды роль доноров электронов могут выполнять некоторые иные восстановленные соединения, например сероводород. При этом не происходит образования О2 (уравнение 13.2).

Данное наблюдение принадлежит Корнелису ван Нилю, который в 1931—1933 гг., будучи еще студентом-дипломником, исследовал особенности фотосинтеза у разных бактерий и сделал смелое предположение, перевернувшее взгляды исследователей на закономерности фотосинтеза. Исходя из выведенного им уравнения фотосинтеза для пурпурных серных бактерий (13.2), К. ван Ниль предположил, что не углекислота, а вода разлагается при фотосинтезе у растений, образуя молекулярный кислород, и предложил общее уравнение фотосинтеза (13.3):
В последнем уравнении H2A представляет собой донор электронов, которым у оксигенных организмов служит вода, а у аноксигенных бактерий—другие вещества (сероводород, молекулярный водород, изопропанол и др.). При этом выделяется дегидрированный донор электронов (А) и происходит восстановление углекислоты до органического соединения (СН2О). Эти превращения и составляют суть процесса фотосинтеза.
Интересно отметить, что вода является очень плохим донором электронов и ни один из окислителей, которыми располагают живые организмы, не является достаточно мощным, чтобы отщепить от молекулы Н2О атомы водорода. Это становится возможным лишь в результате фотохимических превращений особых молекул хлорофиллов, которые становятся сильными окислительными агентами и приобретают способность окислять воду.
Процесс фотосинтеза требует обязательного участия мембран. У прокариот эту роль выполняют впячивания плазматической мембраны, а у эукариот — мембраны тилакоидов, расположенные в хлоропластах. Тилакоиды находятся в строме хлоропластов, образуя стопки (граны). Внутреннее содержимое тилакоидов называют люменом.
В мембранах тилакоидов располагаются компоненты транспорта электронов, сгруппированные в 2 фотосистемы, и АТР-синтаза. Фотосистемы используются для переноса электронов и сопряженного с ним перевода протонов в люмен, в результате чего на тилакоидной мембране создается протонный градиент. Энергия протонного градиента запускает синтез АТР, подобно тому, как это имеет место при окислительном фосфорилировании. Однако, в отличие от дыхательной цепи, в фотосистемах электроны движутся от плохого донора (молекулы воды) к плохому акцептору (NADP+), т. е. в противоположном по сравнению с дыханием направлении. Соответственно и энергия при таком направленном «вверх» потоке электронов должна затрачиваться. Действительно, для восстановления NADP+ электроны должны дважды возбудиться светом.
Характеристика компонентов фотосистем. Компоненты фотосистем представляют собой белковые комплексы, содержащие фоточувствительные молекулы (пигменты) и переносчики восстановительных эквивалентов.
Пигменты представлены в основном хлорофиллами, каротиноидами и фикобилинами. Основную роль в процессе фотосинтеза играет зеленый, содержащий ионы магния тетрапиррольный пигмент — хлоро-филл a (рис. 13.1). Этот хромофор представляет собой магнийпорфирин и похож по структуре на гем (железопорфирин).
нако у хлорофилла a имеются следующие основные отличия: с одним из пиррольных колец слито пятое, циклопентановое кольцо; одно из пиррольных колец частично восстановлено; одна из кислотных боковых цепей представляет собой эфир фитола—двадцатиуглеродного высокогидрофобного спирта. Остаток фитола придает молекуле хлорофилла амфифильные свойства и служит «якорем», с помощью которого хромофор может удерживаться в липидном бислое тилакоидной мембраны. При этом гидрофильная голова (магнийпорфирин) находится на поверхности мембраны, обращенной к водной фазе стромы, и расположена параллельно плоскости мембраны. Таким образом, достигается ориентация хлорофилла в хлоропластах, способствующая наиболее эффективному улавливанию световой энергии.
Другие хлорофиллы зеленых растений (b, c, феофитин), а также хлорофиллы некоторых водорослей и бактерий (хлоробиум-хлорофилл, бактериохлорофиллы) отличаются от хлорофилла a структурой заместителей у пиррольных колец, отсутствием иона магния (феофитин), спектрами поглощения. Существование в клетке хлорофиллов с разными спектрами поглощения позволяет увеличить диапазон улавливаемого излучения.
Каротиноиды и фикобилины служат вспомогательными пигментами. Они еще больше увеличивают спектр поглощаемой организмом энергии, а кроме того, защищают хлорофиллы от избытка света и от окисления кислородом, который выделяется при фотосинтезе.
ротиноиды — это желтые, оранжевые, красные или коричневые пигменты, сильно поглощающие в фиолетовой области спектра. В хлоропластах присутствуют две группы каротиноидов—каротины(углеводороды, бульшую часть которых составляют тетратерпены, подробно рассматриваются в главе 17) и ксантофиллы (по химическому строению сходны с каротинами, но содержат кислород). Фикобилины характерны для цианобактерий и хлоропластов красных водорослей.
Для более полного улавливания световой энергии пигменты в фотосистемах собраны с помощью белков в антенные комплексы. В каждом антенном комплексе насчитывается по несколько сотен молекул пигментов, и их основная функция состоит в передаче поглощенной энергии хлорофиллу а. Таким образом, можно сравнить антенный комплекс с воронкой, «загоняющей» энергию к хлорофиллу реакционного центра фотосистемы. Энергия возбуждения (экситон) передается от молекулы к молекуле посредством электрического поля возбужденного электрона (очевидно, возбужденный электрон в молекуле, поглотившей квант света, передает свою энергию близлежащей молекуле таким образом, что в той тоже возбуждается электрон).
Транспортирующие электроны компоненты фотосистем представлены комплексом цитохрома b/f (агрегат интегральных мембранных белков, содержащий два цитохрома: b563 и f), феофитином, мембраносвязанными пластохинонами (QA и QB), а также мобильными переносчиками, функции которых выполняют пластохинон QP, пластоцианин и ферредоксин.
астохинон по структуре и функциям очень напоминает убихинон (рис. 12.1). Пластоцианин представляет собой белок с одним атомом меди, координационно связанным с боковыми цепями аминокислот: при переносе электрона Cu находится попеременно в степени окисленности +1 и +2. Ферредоксин является железосерным белком типа 4Fe, 4S2-, переносящим электроны. Цепь переносчиков замыкает фермент, транспортирующий электроны на NADP+.
Установлено, что фотосинтез требует участия двух фотосистем: II и I. Фотосистема II содержит в реакционном центре хлорофилл a, имеющий оптимум поглощения при 680 нм (Р680), а в реакционном центре фотосистемы I (Р700) содержится хлорофилл a с оптимумом поглощения 700 нм.
Световые реакции фотосинтеза. Процесс фотосинтеза условно можно разделить на 2 этапа: световые и темновые реакции. Световые реакции требуют участия видимого света и осуществляются в тилакоидных мембранах. Их итогом является восстановление NADP+ и синтез АТР. Темновые реакции могут осуществляться и в условиях отсутствия видимого света, в строме, и под ними обычно подразумевают восстановление СО2 при участии АТР и NADPH.
У растений фотосинтетический перенос электронов начинается с фотосистемы II. Энергия поглощенного света поступает от антенных комплексов к хлорофиллу a реакционного центра Р680 и переводит один из его электронов в возбужденное состояние. Возбужденный электрон сразу передается на расположенный рядом феофитин, а в молекуле хлорофилла появляется положительно заряженная «дырка» с очень высоким сродством к электрону, т.е. возникает положительно заряженный радикал Р680. «Дырка» очень быстро заполняется электроном, извлеченным из воды водорасщепляющим ферментом
при участии ионов марганца. Возбужденный электрон мигрирует по цепи переносчиков (рис. 13.2) и достигает второй «дырки» в Р700 фотосистемы I, которая, в свою очередь, образовалась при переносе «горячего» электрона на электронный акцептор (по-видимому, пластохинон). Здесь электрон возбуждается вторично и далее по переносчикам фотосистемы I передается на NADP+ (рис. 13.2). Такое движение электронов по цепи переносчиков называется нециклическим потоком, или Z-схемой. Важной отличительной особенностью этого процесса является то, что электрон дважды возбуждается в реакционных центрах двух фотосистем, и поэтому его энергии хватает для восстановления NADP+.
Реакцию образования NADPH катализирует ферредоксин-NADP+-редуктаза, содержащая FAD в качестве простетической группы. Следует отметить, что для восстановления никотинамидных кофакторов требуются гидрид-ионы: 2 электрона от двух молекул восстановленного ферредоксина конвергируют, а протон поступает из стромы.
Кроме образования NADPH, нециклический поток электронов обусловливает перенос протонов в люмен и создание на тилакоидной мембране электрохимического градиента. Это происходит на этапе переноса водорода от восстановленного пластохинона QP на комплекс цитохромов b/f, которые, как известно (глава 12), являются переносчиками электронов. Когда протоны, согласно закономерностям облегченной диффузии, выходят из люмена в строму, они используют каналы АТР-синтазы, и происходит синтез АТР. Этот механизм запасания энергии носит название фотофосфорилирование, и его принцип сходен с механизмом окислительного фосфорилирования.
Кроме описанного выше нециклического потока, в тилакоидных мембранах могут осуществляться циклические потоки электронов. В них электрон возбуждается только единожды, в реакционном центре фотосистемы I, и не происходит образования NADPH. Поток становится циклическим, когда электроны от ферредоксина передаются не на NADP+ , а обратно, к пластохинону QP (рис. 13.2, 13.3, пунктирные стрелки). Такое движение электронов обеспечивает клетку только АТР, но не NADPH. Циклический поток электронов преобладает при низких концентрациях NADP+ , что обусловлено накоплением в клетке NADPH.
На рис. 13.3 показано расположение компонентов фотосистем, их субстратов, а также продуктов. Можно видеть, что основные продукты световых реакций фотосинтеза — NADPH и ATP — накапливаются в строме, т. е. там, где они должны расходоваться в темновых реакциях.
Темновые реакции фотосинтеза. К темновым реакциям фотосинтеза относят фиксацию, т. е. восстановление и включение в состав органических соединений, СО2. У большинства фотосинтезирующих организмов этот процесс осуществляется в цикле Кальвина (цикл носит имя Мелвина Кальвина, получившего за его открытие Нобелевскую премию).
Цикл Кальвина условно можно разделить на три этапа: 1) карбоксилирование рибулозодифосфата; 2) восстановление 3-фосфоглицерата до альдегида; 4) регенерация рибулозодифосфата — акцептора СО2.
Углекислота, восстанавливаемая в цикле Кальвина, поступает в строму хлоропластов зеленых растений через устьица в листьях и зеленых стеблях, а в клетки водорослей и цианобактерий — в растворенном виде. Ключевой реакцией восстановления СО2 является карбоксилирование рибулозодифосфата.
Данную реакцию катализирует необычный фермент—рибулозодифосфаткарбоксилаза/оксигеназа, обладающая двумя активностями: карбоксилазной и оксигеназной (рис. 13.4).
Оксигеназная активность реализуется в отсутствии СО2 и в присутствии О2, а продукты этой реакции участвуют в фотодыхании. Рибулозодифосфаткарбоксилаза/оксигеназа очень медленно работает: скорость ее катализа в сотни раз меньше, чем для большинства других ферментов. Поэтому в хлоропластах этот фермент может составлять до половины всего белка. Считается, что это самый распространенный на Земле белок, преобладающий в количественном отношении над другими белками. На уровне данного фермента осуществляется регуляция скорости цикла Кальвина.
Восстановление продуктов карбоксилазной реакции (2 молекул 3фосфоглицерата) происходит с участием фосфоглицераткиназы и глицеральдегид-3-фосфатдегидрогеназы. Обратные реакции, катализируемые этими ферментами, имеют место в гликолизе (рис. 9.3). В этом процессе затрачивается энергия АТР и восстановительные эквивалентны, поставляемые NADPH.
Регенерация рибулозодифосфата осуществляется в результате межмолекулярных перестроек с участием трансальдолаз и транскетолаз (принимают также участие в пентозофосфатных путях, описаны в главе 9). В превращения вступают 3 соединения: глицеральдегид-3-фосфат, образующийся из него в ходе изомеризации дигидроксиацетонфосфат, а также фруктозо-1,6-дифосфат, который образуется в реакции альдольной конденсации из двух триозофосфатов (см. Гликолиз, глава 9).
В результате описанных превращений из 6 молекул СО2 в ходе 6 оборотов цикла Кальвина синтезируется 1 молекула гексозы. При этом затрачивается 18 молекул АТР и 12 молекул NADPH (рис. 13.5). Таким образом, фиксация углекислоты обходится клетке очень дорого: на включение лишь одной молекулы СО2 в органическое соединение расходуется 3 молекулы АТР и 2 молекулы NADPH.
Цикл Кальвина изображают замкнутым, однако, как и в ЦТК, многие его промежуточные продукты используются в качестве предшественников для биосинтеза клеточных соединений. Так, 3-фосфоглицерат мо- жет превращаться в пируват (реакции гликолиза); эритрозо-4-фосфат —
в ароматические аминокислоты; рибозо-5-фосфат — в нуклеотиды; гексозофосфаты — в полисахариды (последние три промежуточных соединения формируются на стадии межмолекулярных перестроек). Так же, как и в других циклах, существуют анаплеротические реакции, не позволяющие циклу прерываться.
Скорость цикла Кальвина строго регулируется, поскольку на фиксацию СО2 не должно расходоваться чересчур много клеточной энергии. Основной принцип регуляции цикла состоит в том, чтобы «подогнать» скорость фиксации СО2 к скорости световых реакций фотосинтеза, где образуются необходимые для фиксации АТР и NADPH. Поэтому, не смотря на то что цикл Кальвина не требует непосредственного участия видимого света, наиболее интенсивно он все же протекает в условиях освещения. Основной стадией, лимитирующей скорость фиксации углекислоты, является карбоксилирование рибулозодифосфата. Активность карбоксилазы значительно увеличивается при освещении, чему способствуют следующие причины:
1) карбоксилаза аллостерически активируется фруктозо-6-фосфатом и ингибируется фруктозо-1,6-дифосфатом. В свою очередь, содержание этих продуктов контролирует фермент фруктозо-1,6-дифосфатаза, которая активируется светом и катализирует расщепление фруктозодифосфата;
2) еще одним аллостерическим активатором карбоксилазы является NADPH, а его количество возрастает на свету при интенсивном нециклическом потоке электронов;
3) скорость рассматриваемой ферментативной реакции возрастает при повышении рН от 7 до 9. Фермент работает в строме, а ее защелачивание является следствием закисления люмена при создании протонного градиента на тилакоидной мембране;
4) карбоксилаза активируется ионами марганца, которые высвобождаются в строму при переносе протонов в люмен в процессе транспорта электронов.
Кроме этого, свет активирует еще один фермент цикла Кальвина — глицеральдегид-3-фосфат-дегидрогеназу, который, очевидно, меняет свою специфичность (от NADH к NADPH) под действием света.
С4 -путь фиксации СО2. Описанный выше процесс включения СО2 в состав 3-фосфоглицерата (результат карбоксилазной реакции рибулозодифосфат-карбоксилазы/оксигеназы) называется С3-путем, а растения, в которых он осуществляется — С3-растениями. В то же время существует альтернативный С4-путь (цикл Хэтча—Слэка), отличающийся присутствием дополнительной начальной стадии — фиксации СО2 в составе четырехуглеродного соединения (оксалоацетата). Эту реакцию катализирует фосфоенолпируваткарбоксилаза — фермент, работающий гораздо быстрее, чем рибулозодифосфат-карбоксилаза/оксигеназа. В результате у С4-растений углекислота включается в состав оксалоацетата намного эффективнее, чем у С3-растений в состав 3-фосфоглицерата. Однако С4-путь требует дополнительных стадий и затрат энергии: на фиксацию одной молекулы СО2 здесь затрачивается 5 молекул АТР вместо 3 в С3-пути (рис. 13.6). Тем не менее, С4-растения (кукуруза, сахарный тростник, сорго, многие сорняки, произрастающие в умеренной зоне и др.) растут го-
раздо быстрее, чем С3-растения (пшеница, рожь, овес, рис и др.). Причиной такого несоответствия служит фотодыхание — расточительный процесс, характерный для всех С3-растений и практически отсутствующий у С4 — растений.
Фотодыхание. Этот процесс обязан существованию оксигеназной активности рибулозодифосфат-карбоксилазы/оксигеназы. Молекулярный кислород конкурирует с СО2 за активный центр данного фермента, и часть рибулозо дифосфата превращается в фосфогликолат (рис.13.4). Этого не происходит у С4-растений, поскольку большинство их имеет своей родиной тропические страны, где высокое испарение влаги заставило растения выработать механизм закрывания устьиц, через которые ткани вентилируются газами, в самые жаркие часы дня (при максимальном солнечном освещении). Чтобы такой механизм не привел к снижению скорости фиксации СО2, растения запасают углекислоту в составе малата или аспартата (образуются из оксалоацетата в цикле Хэтча—Слэка), а затем расходуют по мере надобности (реакции декарбоксилирования). В этом случае концентрация СО2 в зеленых частях растений всегда находится на высоком уровне и отсутствует конкуренция О2 за активный центр фермента, поскольку молекулярный кислород поступает в клетки в ограниченных количествах при закрывании устьиц.
Субстратом фотодыхания служит гликолат, который образуется при дефосфорилировании фосфогликолата. При фотодыхании (в отличие от митохондриального дыхания, тоже характерного для растений в темноте) потребление О2 и выделение СО2 не связано с запасанием энергии, наоборот, здесь расходуются восстановительные эквиваленты и ATP, а фиксированный в цикле Кальвина углерод бесполезно теряется в виде СО2. Показано, что фотодыхание может приводить к реокислению и выделению до 50% углерода, фиксированного в цикле Кальвина. До сих пор не разгадана целесообразность этого столь неэкономного и ограничивающего эффективность роста растений процесса.
Последовательность событий фотодыхания изображена на рис.13.7. Можно видеть, что отдельные стадии этого процесса осуществляются в разных органеллах. Итогом фотодыхания является образование одной
молекулы фосфоглицерата (3 атома «С») из двух молекул гликолата (4 атома «С»), а участие кислорода приводит к окислению четвертого атома углерода в углекислоту. По-видимому, назначение сложной последовательности превращений в фотодыхании состоит в том, чтобы вернуть в цикл хотя бы часть углерода из гликолата, который накапливается в избытке.
При фотодыхании происходит потеря одного атома углерода из каждых четырех, причем следует учитывать, что на фиксацию этого атома углерода уже затрачена энергия в цикле Кальвина. Кроме этого, при окислении глицина и дезаминировании серина выделяется аммиак, на включение которого в состав аминокислот снова придется затрачивать энергию. В результате фотодыхание снижает потенциальную урожайность С3-растений на 30—40%.
Источник: studopedia.ru
История открытия фотосинтеза
В настоящее время школьники впервые знакомятся со сложными процессами фотосинтеза уже в 6 классе.
Но еще 300-400 лет назад ответ на вопрос «откуда растения берут питательные вещества для строительства своих клеток?» занимал умы ученых во всем мире.
Первым и очевидным ответом было предположение, что из земли. Однако, в далеком 1600 году фламандский ученый Ян Батист ван Гельмонт решил проверить влияние почвы на рост растений и провел уникальный в своей простоте опыт. Естествоиспытатель взял веточку ивы и бочку с почвой. Предварительно их взвесил. А затем посадил отросток ивы в бочку с почвой.
Долгие пять лет ван Гельмонт поливал молодое деревце лишь дождевой водой. А через пять лет выкопал деревце, и вновь взвесил отдельно деревце и отдельно почву. Каково же было его удивление, когда весы показали, что деревце увеличило свой вес практически в тридцать раз, и совсем не походило на тот скромный прутик, что был посажен в кадку. А вес почвы уменьшился всего на 56 граммов.
Ученый сделал вывод. что почва практически не дает строительного материала растениям, а все необходимые вещества растение получает из воды.
После ван Гельмонта различные ученые повторили его опыт, и сложилась так называемая «водная теория питания растений».
Одним из тех, кто попытался возразить этой теории был М.В. Ломоносов. И строил он свои возражения на том, что на пустых, скудных северных землях с редкими дождями растут высокие, мощные деревья. Михаил Васильевич предположил, что часть питательных веществ растения впитывают через листья, но доказать свою теорию экспериментально он не смог.
И как часто бывает в науке, помог его величество случай.
Однажды нерадивая мышь, решившая поживиться церковными запасами, случайно перевернула банку и оказалась в ловушке. И через некоторое время погибла. К нашей удаче, эту мышь в банке обнаружил Джозеф Пристли, который был не просто священником, а по совместительству ученым-химиком, и очень интересовался химией газов и способами очистки испорченного воздуха. И тут церковным мышам не повезло. Они стали участницами различных опытов английского ученого.
Джозеф Пристли ставил под одну банку горящую свечу, а в другую сажал мышь. Свеча тухла, грызун погибал.
В наше время его самого зоозащитники посадили бы в банку, но в далеком 1771 году ученому никто не помешал продолжить свои опыты. Пристли посадил мышь в банку, где до этого потухла свеча. Животное погибло еще быстрее.
И тогда Пристли сделал вывод, что раз все живое на Земле до сих пор не погибло, Бог (мы же помним, что Пристли был священником), придумал некий процесс, чтобы воздух вновь был пригоден для жизни. И скорее всего, основная роль в нем принадлежит растениям.
Чтобы доказать это, ученый взял воздух из банки где погибла мышь, и разделил его на две части. В одну банку он поставил мяту в горшочке. А другая банка ждала своего часа. Через 8 дней растение не только не погибло, а даже выпустило несколько новых побегов. И он опять посадил грызунов в банки. В той, где росла мята — мышь была бодра и закусывала листиками. А в той, где мяты не было — практически моментально лежала дохлая мышиная тушка.
Опыты Пристли вдохновили ученых, и во всем мире начали отлавливать мелких грызунов и пытаться повторить его эксперименты.
Но мы же помним, что Пристли был священником и весь день, до вечерней службы мог заниматься исследованиями.
А Карл Шееле, аптекарь из Швейцарии, экспериментировал в домашней лаборатории в свободное от работы время, т.е. по ночам, и мыши дохли у него независимо от присутствия мяты в банке. В результате его экспериментов получалось, что растения не улучшают воздух, а делают его непригодным для жизни. И Шееле обвинил Пристли в обмане научной общественности. Пристли не уступил, и в результате противостояния ученых было установлено, что для восстановления воздуха растениям необходим солнечный свет.
Именно эти опыты положили начало изучению фотосинтеза.
Исследование фотосинтеза стремительно продолжалось. Уже в 1782 году, спустя всего лишь 11 лет после исследований Пристли, швейцарский ботаник Жан Сенебье доказал, что органоиды растений разлагают углекислый газ в присутствии солнечного света. И практически еще сто лет провальных и удачных экспериментов понадобилась ученым разных специальностей, чтобы в 1864 году немецкий ученый Юлиус Сакс смог доказать, что растения потребляют углекислый газ и выделяют кислород в соотношении 1:1.
Значение фотосинтеза для жизни на Земле
И теперь становится понятна важность процесса фотосинтеза для жизни на земле. Именно благодаря этому сложному химическом процессу стало возможно зарождение жизни на земле и существование человека.
Кто-то может возразить, что на Земле есть места, где не растут ни деревья ни кустарники, например, пустыни или Арктические льды. Ученые доказали, что доля кислорода, выделяемого зеленой массой лесов, кустарников и трав — т. е. растений, что обитают на поверхности суши, составляет всего около 20% газообмена, а 80% кислорода приходится на мельчайшие морские и океанские водоросли, которые потоками воздуха переносятся по всей планете, позволяя дышать животным в экстремальных, практически лишенных растительности регионах нашей удивительной планеты.
Благодаря фотосинтезу вокруг нашей планеты сформировался защитный озоновый экран, защищающий все живое на земле от космической и солнечной радиации, и живые организмы смогли выйти на сушу из глубин океана.
Подробнее о «великой кислородной революции» можно прочитать в учебнике «Биология 10-11 классы» под редакцией А.А. Каменского на портале LECTA.
К сожалению, в настоящее время кислород потребляют не только живые существа, но и промышленность. Уничтожаются тропические леса, загрязняются океаны, что приводит к снижению газообмена и увеличению дефицита кислорода.
Определение и формула фотосинтеза
Определение и формула фотосинтеза
Слово фотосинтез состоит из двух частей: фото — «свет» и синтез — «соединение», «создание». Если подходить к определению упрощенно, то фотосинтез — это превращение энергии света в энергию сложных химических связей органических веществ при участии фотосинтетических пигментов. У зеленых растений фотосинтез происходит в хлоропластах.
Схема фотосинтеза, на первый взгляд, проста:
Вода + квант света + углекислый газ → кислород + углевод
или (на языке формул):
6СО2 + 6Н2О → С6Н12О6 + 6О2
Если копнуть поглубже и посмотреть на лист в электронный микроскоп, выяснится удивительная вещь: вода и углекислый газ ни в одной из структурных частей листа непосредственно друг с другом не взаимодействуют.
Фазы фотосинтеза
К фотосинтезу способны не только растения, но и многие одноклеточные животные благодаря специальным органоидам, которые называются хлоропласты.
Хлоропласты — это пластиды зеленого цвета фотосинтезирующих эукариот. В состав хлоропластов входят:
- две мембраны;
- стопки гранов;
- диски тилакоидов;
- строма — внутреннее вещество хлоропласта;
- люмен — внутреннее вещество тилакоида.
Сложный процесс фотосинтеза состоит из двух фаз: световой и темновой. Как понятно из названия, световая (светозависимая) фаза происходит с участием квантов света. Название темновая фаза вовсе не означает, что процесс происходит в темноте. Более точное определение — светонезависимая. Т.е. для реакций, происходящих в этой этой фазе, свет не нужен, а протекает она одновременно со световой, только в других отделах хлоропласта.
Многие делают ошибку, говоря, что в процессе фотосинтеза происходит производство растениями такого необходимого человечеству кислорода. На самом деле фотосинтез — это синтез углеводов (например, глюкозы), а кислород — лишь побочный продукт реакции.
Световая фаза фотосинтеза
Световая фаза фотосинтеза происходит на мембранах тилакоидов. Фотон света, попадая на хлорофилл, возбуждает его и происходит выделение электронов и скопление отрицательно заряженных электронов на мембране. После того, как хлорофилл потерял все свои электроны, квант света продолжает воздействовать на воду, вызывая фотолиз Н2О.
Н2О → Н+ + ОН-
Положительно заряженные протоны водорода накапливаются на внутренней мембране тилакоида.
Получается такой бутерброд: с одной стороны отрицательно заряженные электроны хлорофилла, с другой – положительно заряженные протоны водорода, а между ними – внутренняя мембрана тилакоида.
Гидроксильные ионы идут на производство кислорода:
4ОН → О2 + 2Н2О
Когда количество протонов водорода и электронов достигает максимума, запускается специальный переносчик — АТФ-синтаза. АТФ-синтаза выталкивает протоны водорода в строму, где их подхватывает специальный переносчик никотинамиддинуклеотидфосфат или сокращенно НАДФ. НАДФ — специфический переносчик протонов водорода в реакциях углеводов.
Прохождение протонов водорода через АТФ-синтазу сопровождается синтезом молекул АТФ из АДФ и фосфата или фотофосфорилированием, в отличие от окислительного фосфорилирования.
На этом световая фаза фотосинтеза заканчивается, а НАДФН+ и АТФ переходят в темновую фазу.
Повторим ключевые процессы световой фазы фотосинтеза:
- Фотон попадает на хлорофилл с выделением электронов.
- Фотолиз воды.
- Выделение кислорода.
- Накопление НАДФН+.
- Накопление АТФ.
У некоторых растений фотосинтез идет по упрощенному варианту, который называется «циклическое фосфорилирование» и разбирается этот процесс в учебнике «Биология 10-11 классы» под редакцией А. А. Каменского на портале LECTA.
Темновая фаза фотосинтеза
Темновая фаза фотосинтеза — совокупность ферментативных реакций, которые происходят в строме хлоропласта. Результатом таких реакций является восстановление поглощенного СО2 при помощи НАДФН+ и АТФ из световой фазы, а еще – синтез сложных органических веществ.
В настоящее время учеными открыто три различных варианта реакций, протекающих в темновую фазу фотосинтеза.
В зависимости от метаболизма, СО2 растения делят на:
- С3-растения — большинство сельскохозяйственных культур, произрастающих в умеренном климате, у которых в результате реакций СО2 превращается в фосфоглицериновую кислоту.
- С4-растения — растения тропиков и субтропиков, наиболее живучие сорняки. У этих растений в результате реакций СО2 превращается в оксалоацетат.
- САМ-растения — особый тип С4-фотосинтеза у растений, испытывающих дефицит влаги.
Более подробно остановимся на реакциях С3-фотосинтеза, присущих большинству растений и носящих название цикл Калвина.
Мелвин Калвин, американский химик, в 1961 году за определение последовательности реакций при усвоении СО2 был удостоен Нобелевской премии в области химии.
В ходе реакций цикла образуется глюкоза. Чтобы получилась всего лишь одну молекулу глюкозы, последовательные реакции цикла Кальвина одна за другой происходят целых шесть раз и на ее построение тратится шесть молекул СО2, восемнадцать молекул АТФ, двенадцать НАДФН+ и двадцать четыре протона.
В ходе дальнейших исследований с меченым радиоактивным углеродом было установлено, что у некоторых тропических и субтропических растений синтез углеводов идет другим путем. И в 1966 году австралийские ученые М. Хетч и К. Слэк описали С4-фотосинтез, который в их честь называется циклом Хетча-Слэка.
Главное отличие этих путей фотосинтеза в том, что у С3-растений процесс фотосинтеза протекает лишь в клетках мезофилла, а у С4-растений как в клетках мезофилла, так и в клетках обкладки сосудистых пучков.
На первый взгляд, увеличение количества реакций может показаться лишенным смысла. Однако в природе не существует ничего бессмысленного или излишнего. И путь С4-фотосинтеза — эволюционное приспособление растений к более сухому и жаркому климату. Произрастание в условиях ограниченного водоснабжения привело к снижению транспирации для уменьшения потерь воды, что в свою очередь привело к дефициту диоксида углерода и необходимости его концентрации в клетках обкладки.
Также существует еще один уникальный механизм фотосинтеза, характерный для суккулентов. Он носит название САМ (crassulaceae acid metabolism)— «путь фотосинтеза». Химические реакции напоминают путь метаболизма С4, однако здесь химические реакции разделены не в пространстве, а во времени. Диоксид углерода накапливается в темное время суток.
Протекание фотосинтетических реакций в таком варианте позволяет растениям осуществлять процесс фотосинтеза в условиях значительного дефицита влаги. Считается, что данный путь фотосинтеза сформировался самым последним в ходе эволюции.
Изучая пути фотосинтеза, Вы могли заметить, что в ходе эволюции вырабатываются уникальные приспособительные механизмы к различным условиям существования: от засушливых пустынь до морских глубин.
Тайны живой природы помогут открыть электронные учебники по биологии на портале LECTA.
#ADVERTISING_INSERT#
Источник: rosuchebnik.ru
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + H2O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6H2O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
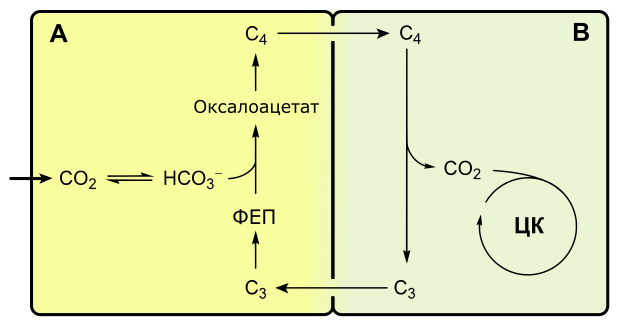
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: biology.su