Основным продуктом фотосинтеза является
Природа первичных продуктов фотосинтеза издавна интересует физиологов. Для решения этого вопроса исследователями были применены разные подходы. Прежде всего была сделана попытка определить фотосинтетический коэффициент. Под фотосинтетическим коэффициентом понимается отношение выделенного в процессе фотосинтеза кислорода к поглощенной СО2. Если в процессе фотосинтеза образуются углеводы, то, согласно приведенному суммарному уравнению, фотосинтетический коэффициент должен быть равен единице:
6О2
— — = 1.
6СО2
Если представить себе, что в процессе фотосинтеза образуются соединения более восстановленные (содержащие меньше кислорода) по сравнению с углеводами, то фотосинтетический коэффициент должен быть больше единицы. Так, расчеты показывают, что в случае образования белков фотосинтетический коэффициент равен 1,25, в случае жира — 1,44.
едняя величина фотосинтетического коэффициента для 27 видов растений оказалась равной 1,04. Расчеты показали, что такая величина фотосинтетического коэффициента указывает на образование наряду с углеводами некоторого количества белка (примерно 12%). Оказалось далее, что величина фотосинтетического коэффициента меняется в зависимости от условий. Так, с улучшением азотного питания фотосинтетический коэффициент повышается. Из этого можно сделать заключение, что в процессе фотосинтеза образуются азотсодержащие соединения.
В настоящее время признано, что наряду с углеводами продуктами фотосинтеза могут быть аминокислоты и, как следствие, белки. При этом в зависимости от условий соотношение продуктов фотосинтеза меняется. Так, преобладание синих лучей над красными приводит к увеличению доли образующихся белков, тогда как красный свет благоприятствует образованию углеводов. Усиление снабжения растений азотом, естественно, также приводит к увеличению первичного синтеза белка.
Фотосинтетическое образование углеводов
В результате двух циклов Кальвина образуется фруктозодифосфат. Из двух молекул фруктозодифосфата (Ф-1,6-диФ) образуются фруктозо-6-фосфат (Ф-6-Ф) и глюкозо-1-фосфат (Г-1-Ф). Глюкозо-1-фосфат, взаимодействуя с уридинтрифосфатом (УТФ) дает уридиндифосфоглюкозу (УДФГ). В свою очередь, УДФГ, реагируя с Ф-6-Ф, дает сахарозофосфат. Из сахарозофосфата путем дефосфорилирования образуется сахароза. Для образования одной молекулы сахарозы необходимо, чтобы прошли четыре цикла Кальвина. По-видимому, именно сахароза является первым свободным сахаром, образующимся в процессе фотосинтеза. Из сахарозы образуются нефосфорилированные моносахара (глюкоза и фруктоза). Крахмал образуется из аденозиндифосфоглюкозы (АДФГ) или уридиндифосфоглюкозы (УДФГ), процесс катализируется ферментом амилосинтетазой.
Фотосинтетическое образование аминокислот
Среди первых продуктов фотосинтеза обнаружены такие аминокислоты, как аланин, серин, глютаминовая кислота, глицин. По-видимому, ФГК, образовавшаяся на первом этапе цикла Кальвина, может превращаться в пировиноградную кислоту. Этот процесс идет особенно интенсивно при недостатке НАДФ. Н2, из-за чего задерживается преобразование ФГК в ФГА (обычный путь в цикле Кальвина). Пировиноградная кислота в присутствии NН3 дает аминокислоту аланин. Показано, что скорость включения 14СО2 в аланин в клетках хлореллы при некоторых условиях может даже превышать скорость ее включения в сахарозу.
При взаимодействии пировиноградной кислоты с глютаминовой образуется аминокислота серин (реакция переаминирования). Из пировиноградной кислоты может образоваться еще ряд органических кислот (в цикле Кребса). Образовавшиеся органические кислоты в процессе аминирования или переаминирования дают аминокислоты. Сам по себе синтез аминокислот еще не означает образование белков. Однако было показано, что между этими двумя процессами имеется прямая связь. Так, под влиянием освещения синими лучами (458—480 нм) усиливается фотосинтетическое образование как аминокислот, так и белков. В присутствии ингибиторов синтеза белка действие синего света не проявляется.
Источник: StudFiles.net
История открытия фотосинтеза
В настоящее время школьники впервые знакомятся со сложными процессами фотосинтеза уже в 6 классе.
Но еще 300-400 лет назад ответ на вопрос «откуда растения берут питательные вещества для строительства своих клеток?» занимал умы ученых во всем мире.
Первым и очевидным ответом было предположение, что из земли. Однако, в далеком 1600 году фламандский ученый Ян Батист ван Гельмонт решил проверить влияние почвы на рост растений и провел уникальный в своей простоте опыт. Естествоиспытатель взял веточку ивы и бочку с почвой. Предварительно их взвесил. А затем посадил отросток ивы в бочку с почвой.
Долгие пять лет ван Гельмонт поливал молодое деревце лишь дождевой водой. А через пять лет выкопал деревце, и вновь взвесил отдельно деревце и отдельно почву. Каково же было его удивление, когда весы показали, что деревце увеличило свой вес практически в тридцать раз, и совсем не походило на тот скромный прутик, что был посажен в кадку. А вес почвы уменьшился всего на 56 граммов.
Ученый сделал вывод. что почва практически не дает строительного материала растениям, а все необходимые вещества растение получает из воды.
После ван Гельмонта различные ученые повторили его опыт, и сложилась так называемая «водная теория питания растений».
Одним из тех, кто попытался возразить этой теории был М.В. Ломоносов. И строил он свои возражения на том, что на пустых, скудных северных землях с редкими дождями растут высокие, мощные деревья. Михаил Васильевич предположил, что часть питательных веществ растения впитывают через листья, но доказать свою теорию экспериментально он не смог.
И как часто бывает в науке, помог его величество случай.
Однажды нерадивая мышь, решившая поживиться церковными запасами, случайно перевернула банку и оказалась в ловушке. И через некоторое время погибла. К нашей удаче, эту мышь в банке обнаружил Джозеф Пристли, который был не просто священником, а по совместительству ученым-химиком, и очень интересовался химией газов и способами очистки испорченного воздуха. И тут церковным мышам не повезло. Они стали участницами различных опытов английского ученого.
Джозеф Пристли ставил под одну банку горящую свечу, а в другую сажал мышь. Свеча тухла, грызун погибал.
В наше время его самого зоозащитники посадили бы в банку, но в далеком 1771 году ученому никто не помешал продолжить свои опыты. Пристли посадил мышь в банку, где до этого потухла свеча. Животное погибло еще быстрее.
И тогда Пристли сделал вывод, что раз все живое на Земле до сих пор не погибло, Бог (мы же помним, что Пристли был священником), придумал некий процесс, чтобы воздух вновь был пригоден для жизни. И скорее всего, основная роль в нем принадлежит растениям.
Чтобы доказать это, ученый взял воздух из банки где погибла мышь, и разделил его на две части. В одну банку он поставил мяту в горшочке. А другая банка ждала своего часа. Через 8 дней растение не только не погибло, а даже выпустило несколько новых побегов. И он опять посадил грызунов в банки. В той, где росла мята — мышь была бодра и закусывала листиками. А в той, где мяты не было — практически моментально лежала дохлая мышиная тушка.

Опыты Пристли вдохновили ученых, и во всем мире начали отлавливать мелких грызунов и пытаться повторить его эксперименты.
Но мы же помним, что Пристли был священником и весь день, до вечерней службы мог заниматься исследованиями.
А Карл Шееле, аптекарь из Швейцарии, экспериментировал в домашней лаборатории в свободное от работы время, т.е. по ночам, и мыши дохли у него независимо от присутствия мяты в банке. В результате его экспериментов получалось, что растения не улучшают воздух, а делают его непригодным для жизни. И Шееле обвинил Пристли в обмане научной общественности. Пристли не уступил, и в результате противостояния ученых было установлено, что для восстановления воздуха растениям необходим солнечный свет.
Именно эти опыты положили начало изучению фотосинтеза.
Исследование фотосинтеза стремительно продолжалось. Уже в 1782 году, спустя всего лишь 11 лет после исследований Пристли, швейцарский ботаник Жан Сенебье доказал, что органоиды растений разлагают углекислый газ в присутствии солнечного света. И практически еще сто лет провальных и удачных экспериментов понадобилась ученым разных специальностей, чтобы в 1864 году немецкий ученый Юлиус Сакс смог доказать, что растения потребляют углекислый газ и выделяют кислород в соотношении 1:1.
Значение фотосинтеза для жизни на Земле
И теперь становится понятна важность процесса фотосинтеза для жизни на земле. Именно благодаря этому сложному химическом процессу стало возможно зарождение жизни на земле и существование человека.
Кто-то может возразить, что на Земле есть места, где не растут ни деревья ни кустарники, например, пустыни или Арктические льды. Ученые доказали, что доля кислорода, выделяемого зеленой массой лесов, кустарников и трав — т. е. растений, что обитают на поверхности суши, составляет всего около 20% газообмена, а 80% кислорода приходится на мельчайшие морские и океанские водоросли, которые потоками воздуха переносятся по всей планете, позволяя дышать животным в экстремальных, практически лишенных растительности регионах нашей удивительной планеты.
Благодаря фотосинтезу вокруг нашей планеты сформировался защитный озоновый экран, защищающий все живое на земле от космической и солнечной радиации, и живые организмы смогли выйти на сушу из глубин океана.
Подробнее о «великой кислородной революции» можно прочитать в учебнике «Биология 10-11 классы» под редакцией А.А. Каменского на портале LECTA.
К сожалению, в настоящее время кислород потребляют не только живые существа, но и промышленность. Уничтожаются тропические леса, загрязняются океаны, что приводит к снижению газообмена и увеличению дефицита кислорода.
Определение и формула фотосинтеза
Определение и формула фотосинтеза
Слово фотосинтез состоит из двух частей: фото — «свет» и синтез — «соединение», «создание». Если подходить к определению упрощенно, то фотосинтез — это превращение энергии света в энергию сложных химических связей органических веществ при участии фотосинтетических пигментов. У зеленых растений фотосинтез происходит в хлоропластах.
Схема фотосинтеза, на первый взгляд, проста:
Вода + квант света + углекислый газ → кислород + углевод
или (на языке формул):
6СО2 + 6Н2О → С6Н12О6 + 6О2
Если копнуть поглубже и посмотреть на лист в электронный микроскоп, выяснится удивительная вещь: вода и углекислый газ ни в одной из структурных частей листа непосредственно друг с другом не взаимодействуют.
Фазы фотосинтеза
К фотосинтезу способны не только растения, но и многие одноклеточные животные благодаря специальным органоидам, которые называются хлоропласты.
Хлоропласты — это пластиды зеленого цвета фотосинтезирующих эукариот. В состав хлоропластов входят:
- две мембраны;
- стопки гранов;
- диски тилакоидов;
- строма — внутреннее вещество хлоропласта;
- люмен — внутреннее вещество тилакоида.
Сложный процесс фотосинтеза состоит из двух фаз: световой и темновой. Как понятно из названия, световая (светозависимая) фаза происходит с участием квантов света. Название темновая фаза вовсе не означает, что процесс происходит в темноте. Более точное определение — светонезависимая. Т.е. для реакций, происходящих в этой этой фазе, свет не нужен, а протекает она одновременно со световой, только в других отделах хлоропласта.
Многие делают ошибку, говоря, что в процессе фотосинтеза происходит производство растениями такого необходимого человечеству кислорода. На самом деле фотосинтез — это синтез углеводов (например, глюкозы), а кислород — лишь побочный продукт реакции.
Световая фаза фотосинтеза
Световая фаза фотосинтеза происходит на мембранах тилакоидов. Фотон света, попадая на хлорофилл, возбуждает его и происходит выделение электронов и скопление отрицательно заряженных электронов на мембране. После того, как хлорофилл потерял все свои электроны, квант света продолжает воздействовать на воду, вызывая фотолиз Н2О.
Н2О → Н+ + ОН-
Положительно заряженные протоны водорода накапливаются на внутренней мембране тилакоида.
Получается такой бутерброд: с одной стороны отрицательно заряженные электроны хлорофилла, с другой – положительно заряженные протоны водорода, а между ними – внутренняя мембрана тилакоида.
Гидроксильные ионы идут на производство кислорода:
4ОН → О2 + 2Н2О
Когда количество протонов водорода и электронов достигает максимума, запускается специальный переносчик — АТФ-синтаза. АТФ-синтаза выталкивает протоны водорода в строму, где их подхватывает специальный переносчик никотинамиддинуклеотидфосфат или сокращенно НАДФ. НАДФ — специфический переносчик протонов водорода в реакциях углеводов.
Прохождение протонов водорода через АТФ-синтазу сопровождается синтезом молекул АТФ из АДФ и фосфата или фотофосфорилированием, в отличие от окислительного фосфорилирования.
На этом световая фаза фотосинтеза заканчивается, а НАДФН+ и АТФ переходят в темновую фазу.
Повторим ключевые процессы световой фазы фотосинтеза:
- Фотон попадает на хлорофилл с выделением электронов.
- Фотолиз воды.
- Выделение кислорода.
- Накопление НАДФН+.
- Накопление АТФ.
У некоторых растений фотосинтез идет по упрощенному варианту, который называется «циклическое фосфорилирование» и разбирается этот процесс в учебнике «Биология 10-11 классы» под редакцией А. А. Каменского на портале LECTA.
Темновая фаза фотосинтеза
Темновая фаза фотосинтеза — совокупность ферментативных реакций, которые происходят в строме хлоропласта. Результатом таких реакций является восстановление поглощенного СО2 при помощи НАДФН+ и АТФ из световой фазы, а еще – синтез сложных органических веществ.
В настоящее время учеными открыто три различных варианта реакций, протекающих в темновую фазу фотосинтеза.
В зависимости от метаболизма, СО2 растения делят на:
- С3-растения — большинство сельскохозяйственных культур, произрастающих в умеренном климате, у которых в результате реакций СО2 превращается в фосфоглицериновую кислоту.
- С4-растения — растения тропиков и субтропиков, наиболее живучие сорняки. У этих растений в результате реакций СО2 превращается в оксалоацетат.
- САМ-растения — особый тип С4-фотосинтеза у растений, испытывающих дефицит влаги.
Более подробно остановимся на реакциях С3-фотосинтеза, присущих большинству растений и носящих название цикл Калвина.
Мелвин Калвин, американский химик, в 1961 году за определение последовательности реакций при усвоении СО2 был удостоен Нобелевской премии в области химии.
В ходе реакций цикла образуется глюкоза. Чтобы получилась всего лишь одну молекулу глюкозы, последовательные реакции цикла Кальвина одна за другой происходят целых шесть раз и на ее построение тратится шесть молекул СО2, восемнадцать молекул АТФ, двенадцать НАДФН+ и двадцать четыре протона.
В ходе дальнейших исследований с меченым радиоактивным углеродом было установлено, что у некоторых тропических и субтропических растений синтез углеводов идет другим путем. И в 1966 году австралийские ученые М. Хетч и К. Слэк описали С4-фотосинтез, который в их честь называется циклом Хетча-Слэка.
Главное отличие этих путей фотосинтеза в том, что у С3-растений процесс фотосинтеза протекает лишь в клетках мезофилла, а у С4-растений как в клетках мезофилла, так и в клетках обкладки сосудистых пучков.
На первый взгляд, увеличение количества реакций может показаться лишенным смысла. Однако в природе не существует ничего бессмысленного или излишнего. И путь С4-фотосинтеза — эволюционное приспособление растений к более сухому и жаркому климату. Произрастание в условиях ограниченного водоснабжения привело к снижению транспирации для уменьшения потерь воды, что в свою очередь привело к дефициту диоксида углерода и необходимости его концентрации в клетках обкладки.
Также существует еще один уникальный механизм фотосинтеза, характерный для суккулентов. Он носит название САМ (crassulaceae acid metabolism)— «путь фотосинтеза». Химические реакции напоминают путь метаболизма С4, однако здесь химические реакции разделены не в пространстве, а во времени. Диоксид углерода накапливается в темное время суток.
Протекание фотосинтетических реакций в таком варианте позволяет растениям осуществлять процесс фотосинтеза в условиях значительного дефицита влаги. Считается, что данный путь фотосинтеза сформировался самым последним в ходе эволюции.
Изучая пути фотосинтеза, Вы могли заметить, что в ходе эволюции вырабатываются уникальные приспособительные механизмы к различным условиям существования: от засушливых пустынь до морских глубин.
Тайны живой природы помогут открыть электронные учебники по биологии на портале LECTA.
#ADVERTISING_INSERT#
Источник: rosuchebnik.ru
Фотосинтез
Фотосинтез — синтез органических веществ из углекислого газа и воды с обязательным использованием энергии света:
6СО2 + 6Н2О + Qсвета → С6Н12О6 + 6О2.
У высших растений органом фотосинтеза является лист, органоидами фотосинтеза — хлоропласты (строение хлоропластов — лекция №7). В мембраны тилакоидов хлоропластов встроены фотосинтетические пигменты: хлорофиллы и каротиноиды. Существует несколько разных типов хлорофилла (a, b, c, d), главным является хлорофилл a. В молекуле хлорофилла можно выделить порфириновую «головку» с атомом магния в центре и фитольный «хвост». Порфириновая «головка» представляет собой плоскую структуру, является гидрофильной и поэтому лежит на той поверхности мембраны, которая обращена к водной среде стромы. Фитольный «хвост» — гидрофобный и за счет этого удерживает молекулу хлорофилла в мембране.
Хлорофиллы поглощают красный и сине-фиолетовый свет, отражают зеленый и поэтому придают растениям характерную зеленую окраску. Молекулы хлорофилла в мембранах тилакоидов организованы в фотосистемы. У растений и синезеленых водорослей имеются фотосистема-1 и фотосистема-2, у фотосинтезирующих бактерий — фотосистема-1. Только фотосистема-2 может разлагать воду с выделением кислорода и отбирать электроны у водорода воды.
Фотосинтез — сложный многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы: реакции световой фазы и реакции темновой фазы.
Световая фаза
Эта фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это приводит к распаду или фотолизу воды:
Н2О + Qсвета → Н+ + ОН—.
Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы •ОН:
ОН— → •ОН + е—.
Радикалы •ОН объединяются, образуя воду и свободный кислород:
4НО• → 2Н2О + О2.
Кислород при этом удаляется во внешнюю среду, а протоны накапливаются внутри тилакоида в «протонном резервуаре». В результате мембрана тилакоида с одной стороны за счет Н+ заряжается положительно, с другой за счет электронов — отрицательно. Когда разность потенциалов между наружной и внутренней сторонами мембраны тилакоида достигает 200 мВ, протоны проталкиваются через каналы АТФ-синтетазы и происходит фосфорилирование АДФ до АТФ; атомарный водород идет на восстановление специфического переносчика НАДФ+ (никотинамидадениндинуклеотидфосфат) до НАДФ·Н2:
2Н+ + 2е— + НАДФ → НАДФ·Н2.
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2 транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.
1 — строма хлоропласта; 2 — тилакоид граны.
Темновая фаза
Эта фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
Первая реакция в этой цепочке — фиксация углекислого газа; акцептором углекислого газа является пятиуглеродный сахар рибулозобифосфат (РиБФ); катализирует реакцию фермент рибулозобифосфат-карбоксилаза (РиБФ-карбоксилаза). В результате карбоксилирования рибулозобисфосфата образуется неустойчивое шестиуглеродное соединение, которое сразу же распадается на две молекулы фосфоглицериновой кислоты (ФГК). Затем происходит цикл реакций, в которых через ряд промежуточных продуктов фосфоглицериновая кислота преобразуется в глюкозу. В этих реакциях используются энергии АТФ и НАДФ·Н2, образованных в световую фазу; цикл этих реакций получил название «цикл Кальвина»:
6СО2 + 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды. В настоящее время различают два типа фотосинтеза: С3— и С4-фотосинтез.
С3-фотосинтез
Это тип фотосинтеза, при котором первым продуктом являются трехуглеродные (С3) соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза (М. Кальвин). Именно С3-фотосинтез описан выше, в рубрике «Темновая фаза». Характерные особенности С3-фотосинтеза: 1) акцептором углекислого газа является РиБФ, 2) реакцию карбоксилирования РиБФ катализирует РиБФ-карбоксилаза, 3) в результате карбоксилирования РиБФ образуется шестиуглеродное соединение, которое распадается на две ФГК. ФГК восстанавливается до триозофосфатов (ТФ). Часть ТФ идет на регенерацию РиБФ, часть превращается в глюкозу.
Фотодыхание
Это светозависимое поглощение кислорода и выделение углекислого газа. Еще в начале прошлого века было установлено, что кислород подавляет фотосинтез. Как оказалось, для РиБФ-карбоксилазы субстратом может быть не только углекислый газ, но и кислород:
О2 + РиБФ → фосфогликолат (2С) + ФГК (3С).
Фермент при этом называется РиБФ-оксигеназой. Кислород является конкурентным ингибитором фиксации углекислого газа. Фосфатная группа отщепляется, и фосфогликолат становится гликолатом, который растение должно утилизировать. Он поступает в пероксисомы, где окисляется до глицина. Глицин поступает в митохондрии, где окисляется до серина, при этом происходит потеря уже фиксированного углерода в виде СО2. В итоге две молекулы гликолата (2С + 2С) превращаются в одну ФГК (3С) и СО2. Фотодыхание приводит к понижению урожайности С3-растений на 30–40% (С3-растения — растения, для которых характерен С3-фотосинтез).
С4-фотосинтез
С4-фотосинтез — фотосинтез, при котором первым продуктом являются четырехуглеродные (С4) соединения. В 1965 году было установлено, что у некоторых растений (сахарный тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются четырехуглеродные кислоты. Такие растения назвали С4-растениями. В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый газ. Путь превращений углерода в С4-растениях стали называть путем Хэтча-Слэка.
Для С4-растений характерно особое анатомическое строение листа. Все проводящие пучки окружены двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки. Углекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват (ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С). Процесс катализируется ФЕП-карбоксилазой. В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2 и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла много гран, где активно идут реакции световой фазы. В хлоропластах клеток обкладки идут реакции темновой фазы.
Оксалоацетат (4С) превращается в малат, который через плазмодесмы транспортируется в клетки обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата, СО2 и НАДФ·Н2.
Пируват возвращается в клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
|
Строение С4-растений: |
|
|
С4-фотосинтез: |
Значение фотосинтеза
Благодаря фотосинтезу, ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются миллиарды тонн кислорода; фотосинтез является основным источником образования органических веществ. Из кислорода образуется озоновый слой, защищающий живые организмы от коротковолновой ультрафиолетовой радиации.
При фотосинтезе зеленый лист использует лишь около 1% падающей на него солнечной энергии, продуктивность составляет около 1 г органического вещества на 1 м2 поверхности в час.
Хемосинтез
Синтез органических соединений из углекислого газа и воды, осуществляемый не за счет энергии света, а за счет энергии окисления неорганических веществ, называется хемосинтезом. К хемосинтезирующим организмам относятся некоторые виды бактерий.
Нитрифицирующие бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3 → HNO2 → HNO3).
Железобактерии превращают закисное железо в окисное (Fe2+ → Fe3+).
Серобактерии окисляют сероводород до серы или серной кислоты (H2S + ½O2 → S + H2O, H2S + 2O2 → H2SO4).
В результате реакций окисления неорганических веществ выделяется энергия, которая запасается бактериями в форме макроэргических связей АТФ. АТФ используется для синтеза органических веществ, который проходит аналогично реакциям темновой фазы фотосинтеза.
Хемосинтезирующие бактерии способствуют накоплению в почве минеральных веществ, улучшают плодородие почвы, способствуют очистке сточных вод и др.
Источник: licey.net
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + H2O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6H2O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
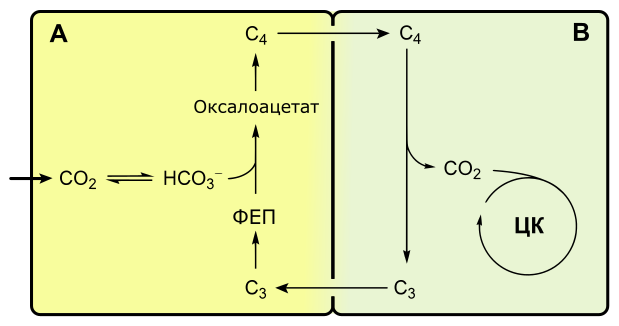
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: biology.su