При фотосинтезе используются вещества
Из истории изучения воздушного питания растений
Знания о питании растений накапливались постепенно. Около 350 лет назад голландский ученый Ян Гельмонт впервые поставил опыт по изучению питания растений. В глиняном горшке с почвой он выращивал иву, добавляя туда только воду. Опадавшие листья ученый тщательно взвешивал. Через пять лет масса ивы вместе с опавшими листьями увеличилась на 74,5 кг, а масса почвы уменьшилась всего на 57 г. На основании этого Гельмонт пришел к выводу, что все вещества в растении образуются не из почвы, а из воды. Мнение о том, что растение увеличивается в размерах только за счет воды, сохранялось до конца XVIII века.
В 1771 г. английский химик Джозеф Пристли изучал углекислый газ, или, как он его называл, «испорченный воздух» и сделал замечательное открытие. Если зажечь свечу и накрыть оо стеклянным колпаком, то, немного погорев, она погаснет. Мышь под таким колпаком начинает задыхаться. Однако если под колпак вместе с мышью поместить ветку мяты, то мышь не задыхается и продолжает жить. Значит, растения «исправляют» воздух, испорченный дыханием животных, то есть превращают углекислый газ в кислород.

В 1862 г. немецкий ботаник Юлиус Сакс с помощью опытов доказал, что зеленые растения не только выделяют кислород, но и создают органические вещества, служащие пищей всем другим организмам.
Фотосинтез
Главное отличие зеленых растений от других живых организмов — наличие в их клетках хлоропластов, содержащих хлорофилл. Хлорофилл обладает свойством улавливать солнечные лучи, энергия которых необходима для создания органических вещсств. Процесс образования органического вещества из углекислого газа и воды с помощью солнечной энергии называется фотосинтезом (греч. рЬо1оз свет). В процессе фотосинтеза образуются не только органические вещества — сахара, но и выделяется кислород.
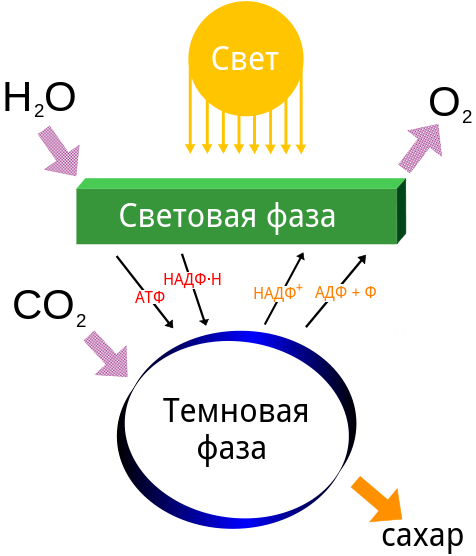
Схематически процесс фотосинтеза можно изобразить так:
![]()
Вода поглощается корнями и по проводящей системе корней и стебля передвигается к листьям. Углекислый газ — составная часть воздуха. Он поступает в листья через открытые устьица. Поглощению углекислого газа способствует строение листа: плоская поверхность листовых пластинок, увеличивающая площадь соприкосновения с воздухом, и наличие большого числа устьиц в кожице.
Образующиеся в результате фотосинтеза сахара превращаются в крахмал. Крахмал это органическое вещество, которое не растворяется в воде. Кго легко обнаружить с помощью раствора йода.
Доказательства образования крахмала в листьях на свету
Докажем, что в зеленых листьях растений из углекислого газа и воды образуется крахмал. Для этого рассмотрим опыт, который в свое время был поставлен Юлиусом Саксом.
Комнатное растение (герань или примулу) выдерживают двое суток в темноте, чтобы весь крахмал израсходовался на процессы жизнедеятельности. Затем несколько листьев закрывают с двух сторон черной бумагой так, чтобы была прикрыта только их часть. Днем растение выставляют на свет, а ночью его дополнительно освещают с помощью настольной лампы.
Через сутки исследуемые листья срезают. Чтобы выяснить, в какой части листа образовался крахмал, листья кипятят в воле (чтобы набухли крахмальные зерна), а затем выдерживают в горячем спирте (хлорофилл при этом растворяется, и лист обесцвечивается). Затем листья промывают в воде и действуют на них слабым раствором йода. Тс участки листьев, которые были на свету, приобретают от действия йода синюю окраску. Это означает, что крахмал образовался в клетках освещенной части листа. Следовательно, фотосинтез происходит только на свету.
Доказательства необходимости углекислого газа для фотосинтеза
Чтобы доказать, что для образования крахмала в листьях необходим углекислый газ, комнатное растение также предварительно выдерживают в темноте. Затем один из листьев помещают в колбу с небольшим количеством известковой воды. Колбу закрывают ватным тампоном. Растение выставляют на свет. Углекислый газ поглощается известковой водой, поэтому его в колбе не будет. Лист срезается, и так же, как в предыдущем опыте, исследуется на наличие крахмала. Он выдерживается в горячей воде и спирте, обрабатывается раствором йода. Однако в этом случае результат опыта будет иным: лист не окрашивается в синий цвет, т.к. крахмал в нем не содержится. Следовательно, для образования крахмала, кроме света и воды, необходим углекислый газ.
Таким образом, мы ответили на вопрос, какую пищу получает растение из воздуха. Опыт показал, что это углекислый газ. Он необходим для образования органического вещества.
Организмы, самостоятельно создающие органические вещества для построения своего тела, называются автотрофамн (греч. autos — сам, trofe — пища).
Доказательства образования кислорода в процессе фотосинтеза
Чтобы доказать, что при фотосинтезе растения во внешнюю среду выделяют кислород, рассмотрим опыт с водным растением элодеей. Побеги элодеи опускают в сосуд с водой и сверху накрывают воронкой. На конец воронки надевают пробирку с водой. Растение выставляют на свет на двое-трое суток. На свету элодея выделяет пузырьки газа. Они скапливаются в верхней части пробирки, вытесняя воду. Для того чтобы выяснить, какой это газ, пробирку аккуратно снимают и вносят в нее тлеющую лучинку. Лучинка ярко вспыхивает. Это значит, что в колбе накопился газ, поддерживающий горение кислород.
Космическая роль растений
Растения, содержащие хлорофилл, способны усваивать солнечную энергию. Поэтому К.А. Тимирязев назвал их роль на Земле космической. Часть энергии Солнца, запасенная в органическом веществе, может долго сохраняться. Каменный уголь, торф, нефть образованы веществами, которые в далекие геологические времена были созданы зелеными растениями и вобрали в себя энергию Солнца. Сжигая природные горючие материалы, человек освобождает энергию, запасенную миллионы лет назад зелеными растениями.
Источник: ebiology.ru
Бактериальный фотосинтез и общее уравнение фотосинтеза
Наряду с фотосинтезом высших растений и водорослей, сопровождаемым выделением O2, в природе осуществляется бактериальный фотосинтез, в котором окисляемым субстратом является не вода, а другие соединения, обладающие более выраженными восстановительными свойствами, например H2S, SO2. Кислород при бактериальном фотосинтезе не выделяется, например:
Фотосинтезирующие бактерии способны использовать не только видимое, но и ближнее ИК излучение (до 1000 нм) в соответствии со спектрами поглощения преобладающих в них пигментов — бактериохлорофиллов. Бактериальный фотосинтез не имеет существенного значения в глобальном запасании солнечной энергии, но важен для понимания общих механизмов фотосинтеза Кроме того, локально бескислородный фотосинтез может вносить существенный вклад в суммарную продуктивность планктона. Так, в Черном море количество хлорофилла и бактериохлорофилла в столбе воды в ряде мест приблизительно одинаково.
Учитывая данные о фотосинтезе высших растений, водорослей и фотосинтезирующих бактерий, обобщенное уравнение фотосинтеза можно записать в виде:
А — кислород в случае высших растений и водорослей, S либо другие элементы — в бактериальном фотосинтезе.
Молекулярны механизм фотосинтеза и структура фотосинтетического аппарата.
С использованием изотопных меток показано, что источником O2 в фотосинтезе является только вода:
Фотосинтез пространственно и во времени разделяется на два сравнительно обособленных процесса: световую стадию окисления воды и темновую стадию восстановления CO2 (рис. 1). Обе эти стадии осуществляются у высших растений и водорослей в специализированных органеллах клетки — хлоропластах. Исключение — синезеленые водоросли (цианобактерии), у которых нет аппарата фотосинтеза, обособленного от цитоплазматических мембран.
Схема световой и темновой стадий фотосинтеза
Хлоропласт, представляющий собой замкнутую структуру, отделенную от остальной части клетки оболочкой, заключает в себе весь фотосинтетический аппарат. Световая стадия реализуется в мембранных структурах хлоропласта (так называемых тилакоидах), тогда как темновая стадия происходит в жидком содержимом хлоропласта (строме) при участии водорастворимых ферментов. У фотосинтезирующих бактерий хлоропласты отсутствуют, но световая стадия также осуществляется в мембранных образованиях — в так называемых хроматофорах.
Световая стадия фотосинтеза
Минимальная функциональная единица, еще способная осуществлять световую стадию фотосинтеза,- тилакоид. Он представляет собой микроскопический плоский диск, образованный белковолипидными мембранами, в которых находятся пигменты. В эти мембраны встроены все компоненты, необходимые для окисления воды, восстановления кофермента никотинамиддинуклеотидфосфата (НАДФ) до НАДФН и синтеза АТФ из аденозиндифосфата. Световая стадия фотосинтеза инициируется поглощением кванта света пигментами, организованными в специальные светособирающие комплексы. Среди пигментов преобладает хлорофилл а. К вспомогательным пигментам относятся хлорофилл b, каротиноиды и др. Наличие светособирающей структуры из нескольких сотен или десятков молекул пигментов на каждый фотохимически активный (реакционный) центр на 2-3 порядка увеличивает сечение захвата излучения и обеспечивает возможность фотосинтеза при слабом освещении.
Часть вспомогательных пигментов, спектрально наиболее близких к фотохимически активному хлорофиллу, непосредственно окружает каждый из реакционных центров, образуя так называемые антенны.
Высокая эффективность переноса возбуждения от молекулы, поглотившей квант, к фотохимическому центру определяется спектральными свойствами и структурной организацией пигментов светособирающего комплекса и антенны, окружающей фотохимический центр. Эти пигменты обеспечивают передачу возбуждения за время менее 100 пс в пределах времени жизни синглетно возбужденного состояния хлорофилла.
В реакционном центре фотосинтеза, куда почти со 100%-ной вероятностью переносится возбуждение, происходит первичная реакция между фотохимически активной молекулой хлорофилла а (у бактерий — бактериохлорофилла) и первичным акцептором электрона (ПА). Дальнейшие реакции в тилакоидных мембранах происходят между молекулами в их основных состояниях и не требуют возбуждения светом. Эти реакции организованы в электронтранспортную цепь — последовательность фиксированных в мембране переносчиков электрона. В электронтранспортной цепи высших растений и водорослей содержится два фотохимических центра (фотосистемы), действующих последовательно (рис. 2), в бактериальной электронтранспортной цепи — один (рис. 3).
Рис. 2. Схема электронтранспортной цепи фотосинтеза растений и синезеленых водорослей
В фотосистеме II высших растений и водорослей синглетно возбужденный хлорофилл а в центре Р680 (число 680 обозначает, что максимум спектральных изменений системы при возбуждении светом находится вблизи 680 нм) отдает электрон через промежуточный акцептор к феофитину (ФЕО, безмагниевый аналог хлорофилла), образуя катион-радикал .
ион-радикал восстановленного феофитина служит далее донором электрона для связанного пластохинона (ПХ*; отличается от убихинонов заместителями в хиноидном кольце), координированного с ионом Fe3+ (в бактериях имеется аналогичный Fе3+-убихинонный комплекс). Далее электрон переносится по цепи, включающей свободный пластохинон (ПХ), присутствующий в избытке по отношению к остальным компонентам цепи, затем цитохромы (Ц) b6 и f, образующие комплекс с железо-серным центром, через медьсодержащий белок пластоцианин (ПЦ; мол. м. 10400) к реакционному центру фотосистемы I.
Центры быстро восстанавливаются, принимая электрон через ряд промежуточных переносчиков от воды. Образование O2 требует последовательного четырехкратного возбуждения реакционного центра фотосистемы П и катализируется мембранным комплексом, содержащим Mn.
Хлорофилл a в фотосистеме I, имеющий максимум поглощения вблизи 700 нм (центр Р700), является первичным фотовозбуждаемым донором электрона, который он отдает первичному акцептору (ПА; его природа однозначно не установлена), а затем, через ряд промежуточных переносчиков (Ai) — растворимому белку ферредоксину (ФД), восстанавливающему с помощью фермента ферредоксин-НАДФ-редуктазы (ФНР) НАДФ до НАДФН. Катион-радикал окисленного пигмента
восстанавливается пластоцианином.
В зрелых хлоропластах имеются граны (стопки тилакоидов), в мембранах которых присутствуют все компоненты злектронтранспортной цепи, и так называемые агранальные тилакоиды, не содержащие фотосистемы II.
Благодаря асимметрическому расположению компонентов электронтранспортной цепи относительно плоскости мембраны при разделении зарядов между хлорофиллом в каждом из двух фотосинтетических центров и акцептором электрона на тилакоидной мембране создается разность электрических потенциалов (плюс — на внутренней, минус — на внешней ее стороне). Перенос электрона пластохиноном сопровождается транспортом протонов, которые захватываются снаружи тилакоида при восстановлении пластохинона и освобождаются внутрь тилакоида при окислении пластогидрохинона. Перенос электронов сопряжен с синтезом АТФ из аденозиндифосфата (АДФ) и неорганического фосфата. Предполагают, что обратный транспорт протонов из тилакоидов в строму через белковый сопрягающий фактор (Н+-АТФ-синтетазу) сопровождается образованием АТФ.
Фотосистема I может действовать автономно без контакта с системой II. В этом случае циклический перенос электрона (на схеме показан пунктиром) сопровождается синтезом АТФ, а не НАДФН. Образующиеся в световой стадии коферменты НАДФН и АТФ используются в темновой стадии фотосинтез, в ходе которой снова образуется НАДФ и АДФ.
Электронтранспортные цепи фотосинтезирующих бактерий в основных своих чертах аналогичны отдельным фрагментам таковых в хлорогпастах высших растений. На рис. 3 показана электронтранспортная цепь пурпурных бактерий.
Рис. 3. Схема электронтранспортной цепи пурпурных бактерий
Темновая стадия фотосинтеза
Все фотосинтезирующие организмы, выделяющие O2, а также некоторые фотосинтезирующие бактерии сначала восстанавливают CO2 до фосфатов Сахаров в так называемом Калвина. У фотосинтезирующих бактерий встречаются, по-видимому, и другие механизмы. Большинство ферментов цикла Калвина находится в растворимом состоянии в строме хлоропластов.
Рис. 4. Упрощенная схема цикла Калвина
Упрощенная схема цикла показана на рис. 4. Первая стадия — карбоксилирование рибулозо-1,5-дифосфата и гидролиз продукта с ооразованием двух молекул 3-фосфоглицериновой кислоты. Эта С3-кислота фосфорилируется АТФ с образованием 3-фосфоглицероилфосфата, который затем восстанавливается НАДФН до глицеральдегид-3-фосфата. Полученный триозофосфат затем вступает в ряд реакций изомеризации, конденсации и перегруппировок, дающих 3 молекулы рибулозо-5-фосфата. Последний фосфорилируется при участии АТФ с образованием рибулозо-1,5-дифосфата и, таким образом, цикл замыкается. Одна из 6 образующихся молекул глицеральдегид-3-фосфата превращается в глюкозо-6-фосфат и используется затем для синтеза крахмала либо выделяется из хлоропласта в цитоплазму. Глицеральдегид-3-фосфат может также превращаться в 3-глицерофосфат и затем в липиды. Триозофосфаты, поступающие из хлоропласта, превращаются в основном в сахарозу, которая переносится из листа в другие части растения.
В одном полном обороте цикла Калвина расходуется 9 молекул АТФ и 6 молекул НАДФН для образования одной молекулы 3-фосфоглицериновой кислоты. Энергетическая эффективность цикла (отношение энергии фотонов, необходимых для фотосинтеза АТФ и НАДФН, к ΔG0 образования углевода из CO2) с учетом действующих в строме хлоропласта концентраций субстратов составляет 83%. В самом цикле Калвина нет фотохимических стадий, но световые стадии могут косвенно влиять на него (в том числе и на реакции, не требующие АТФ или НАДФН) через изменения концентраций ионов Mg2+ и H+, а также уровня восстановленности ферредоксина.
Некоторые высшие растения, приспособившиеся к высокой интенсивности света и к теплому климату (например, сахарный тростник, кукуруза), способны предварительно фиксировать CO2 в дополнительном С4-цикле. При этом CO2 сначала включается в обмен четырехуглеродных дикарбоновых кислот, которые затем декарбоксилируются там, где локализован цикл Калвина. С4-цикл характерен для растений с особым анатомическим строением листа и разделением функций между двумя типами клеток: мезофильных, где сосредоточено карбоксилирование фосфоенолпировиноградной кислоты, и клеток обкладки сосудистого пучка, где функционирует цикл Калвина. Образующаяся в С4-цикле щавелевоуксусная кислота восстанавливается НАДФН до яблочной, которая перемещается в клетки сосудистой обкладки и здесь подвергается окислительному декарбоксилированию, образуя пировиноградную кислоту, CO2 и НАДФН. Два последних используются в цикле Калвина, а пировиноградная кислота возвращается в С4-цикл (рис. 5). Физиологический смысл С4-цикла состоит в запасании CO2 и повышении, таким образом, общей эффективности процесса.
Рис. 5. С4-цикл фиксации CO2
Для кактусов, молочая и других засухоустойчивых растений характерно частичное разделение фиксации CO2 и фотосинтеза во времени (CAM-обмен, или обмен по типу толстянковых; CAM сокр. от англ. Crassulaceae acid metabolism). Днем устьица (каналы, через которые осуществляется газообмен с атмосферой) закрываются, чтобы уменьшить испарение воды. При этом поступление CO2 также затруднено. Ночью устьица открываются, происходит фиксация CO2 в виде фосфоенол-пировиноградной кислоты с образованием С4-кислот, которые днем декарбоксилируются, а освобождаемый при этом CO2 включается в цикл Калвина (рис. 6).
Рис. 6. CAM-тип углеродного обмена
Фотосинтез галобактерий
Единственный известный в природе нехлорофилльный способ запасания энергии света осуществляют бактерии Halobacterium halobium. Ha ярком свету при пониженной концентрации O2 они образуют в своих мембранах пурпурный белок бактериородопсин. В результате индуцированной светом цис-транс-изомеризации ретиналя (хромофора этого пигмента) происходит поглощение H+ и синтез АТФ. Последний используется для частичного обеспечения энергетических потребностей клетки.
Фотосинтез: историческая справка
Ок. 1770 Дж.Пристли обнаружил, что растения выделяют O2. В 1779 Я.Ингенхауз установил, что для этого необходим свет и что O2 выделяют только зеленые части растений. Ж.Сенебье в 1782 показал, что для питания растений требуется CO2; в начале 19 в. H.Соссюр, исходя из закона сохранения массы, подтвердил, что большая часть массы растений создается из CO2 и воды. В 1817 П.Пельтье и Ж.Каванту выделили зеленый пигмент хлорофилл. Позже К.А.Тимирязев показал близость спектра действия фотосинтеза и спектра поглощения хлорофилла. Ю.Сакс в середине 19 в., по-видимому, первым осознал, что этот продукт накапливается в хлоропластах, а Т.В.Энгельман доказал, что именно там же выделяется и O2.
В работах Ф.Блэкмана (1905), P.Эмерсона и У.Арнолда (1932), а также P.Хилла (1936-41) показано наличие световой и темновой стадий фотосинтеза и экспериментально реализована световая стадия в отсутствие CO2 с использованием искусственных акцепторов электрона. Тем самым были получены подтверждения представлений об образовании O2 путем окисления воды. Окончательно это было доказано масс-спектрометрическим методом (С.Рубен, M.Камен, а также А.П.Виноградов и Р.В.Тейс, 1941).
В 1935-41 К.Ван Ниль обобщил данные по фотосинтезу высших растений и бактерий и предложил общее уравнение, охватывающее все типы фотосинтеза X.Гаффрон и К.Воль, а также Л.Дёйсенс в 1936-52 на основе количественных измерений выхода продуктов фотосинтеза поглощенного света и содержания хлорофилла сформулировали представление о «фотосинтетической единице» — ансамбле молекул пигмента, осуществляющих светосбор и обслуживающих фотохимический центр.
В 40-50-х гг. M.Калвин, используя изотоп 14C, выявил механизм фиксации CO2. Д.Арнон (1954) открыл фотофосфорилирование (инициируемый светом синтез АТФ из АДФ и H3PO4) и сформулировал концепцию электронного транспорта в мембранах хлоропластов. P.Эмерсон и Ч.M.Льюис (1942-43) обнаружили резкое снижение эффективности фотосинтеза при 700 нм (красное падение, или первый эффект Эмерсона), а в 1957 Эмерсон наблюдал неаддитивное усиление фотосинтеза при добавлении света низкой интенсивности с 650 нм к дальнему красному свету (эффект усиления, или второй эффект Эмерсона). На этом основании в 60-х гг. сформулировано представление о последовательно действующих фотосистемах в электронтранспортной цепи фотосинтеза с максимумами в спектрах действия вблизи 680 и 700 нм.
Основные закономерности образования O2 при окислении воды в фотосинтезе установлены в работах Б.Кока и П.Жолио (1969-70). Близится к завершению выяснение молекулярной организации мембранного комплекса, катализирующего этот процесс. В 80-х гг. методом рентгеновского структурного анализа детально изучена структура отдельных компонентов фотосинтетического аппарата, включая реакционные центры и светособирающие комплексы (И.Дайзенхофер, X.Михель, P.Хубер).
Лит.: Клейтон Р., Фотосинтез. Физические механизмы и химические модели, пер. с англ., M., 1984; «Ж. Всес. хим. об-ва им. Д.И.Менделеева», 1986, т. 31, № 6; Фотосинтез, под ред. Говинджи, пер. с англ., т. 1-2, M., 1987; Итоги науки и техники, сер. Биофизика, т. 20-22, M., 1987.
© М.Г.Гольдфельд.
Источник: chemport.ru
Фотосинтез — это процесс синтеза органических веществ из неорганических за счет энергии света. В подавляющем большинстве случаев фотосинтез осуществляют растения с помощью таких клеточных органелл как хлоропласты, содержащих зеленый пигмент хлорофилл.
Если бы растения не были способны к синтезу органики, то почти всем остальным организмам на Земле нечем было бы питаться, так как животные, грибы и многие бактерии не могут синтезировать органические вещества из неорганических. Они лишь поглощают готовые, расщепляют их на более простые, из которых снова собирают сложные, но уже характерные для своего тела.
Так обстоит дело, если говорить о фотосинтезе и его роли совсем кратко. Чтобы понять фотосинтез, нужно сказать больше: какие конкретно неорганические вещества используются, как происходит синтез?
Для фотосинтеза нужны два неорганических вещества — углекислый газ (CO2) и вода (H2O). Первый поглощается из воздуха надземными частями растений в основном через устьица. Вода — из почвы, откуда доставляется в фотосинтезирующие клетки проводящей системой растений. Также для фотосинтеза нужна энергия фотонов (hν), но их нельзя отнести к веществу.
В общей сложности в результате фотосинтеза образуется органическое вещество и кислород (O2). Обычно под органическим веществом чаще всего имеют в виду глюкозу (C6H12O6).
Органические соединения большей частью состоят из атомов углерода, водорода и кислорода. Именно они содержатся в углекислом газе и воде. Однако при фотосинтезе происходит выделение кислорода. Его атомы берутся из воды.
Кратко и обобщенно уравнение реакции фотосинтеза принято записывать так:
6CO2 + 6H2O → C6H12O6 + 6O2
Но это уравнение не отражает сути фотосинтеза, не делает его понятным. Посмотрите, хотя уравнение сбалансированно, в нем общее количество атомов в свободном кислороде 12. Но мы сказали, что они берутся из воды, а там их только 6.
На самом деле фотосинтез протекает в две фазы. Первая называется световой, вторая — темновой. Такие названия обусловлены тем, что свет нужен только для световой фазы, темновая фаза независима от его наличия, но это не значит, что она идет в темноте. Световая фаза протекает на мембранах тилакоидов хлоропласта, темновая — в строме хлоропласта.
В световую фазу связывания CO2 не происходит. Происходит лишь улавливание солнечной энергии хлорофилльными комплексами, запасание ее в АТФ, использование энергии на восстановление НАДФ до НАДФ*H2. Поток энергии от возбужденного светом хлорофилла обеспечивается электронами, передающимися по электрон-транспортной цепи ферментов, встроенных в мембраны тилакоидов.
Водород для НАДФ берется из воды, которая под действием солнечного света разлагается на атомы кислорода, протоны водорода и электроны. Этот процесс называется фотолизом. Кислород из воды для фотосинтеза не нужен. Атомы кислорода из двух молекул воды соединяются с образованием молекулярного кислорода. Уравнение реакции световой фазы фотосинтеза кратко выглядит так:
H2O + (АДФ+Ф) + НАДФ → АТФ + НАДФ*H2 + ½O2
Таким образом, выделение кислорода происходит в световую фазу фотосинтеза. Количество молекул АТФ, синтезированных из АДФ и фосфорной кислоты, приходящихся на фотолиз одной молекулы воды, может быть различным: одна или две.
Итак, из световой фазы в темновую поступают АТФ и НАДФ*H2. Здесь энергия первого и восстановительная сила второго тратятся на связывание углекислого газа. Этот этап фотосинтеза невозможно объяснить просто и кратко, потому что он протекает не так, что шесть молекул CO2 объединяются с водородом, высвобождаемым из молекул НАДФ*H2, и образуется глюкоза:
6CO2 + 6НАДФ*H2 →С6H12O6 + 6НАДФ
(реакция идет с затратой энергии АТФ, которая распадается на АДФ и фосфорную кислоту).
Приведенная реакция – лишь упрощение для облегчения понимания. На самом деле молекулы углекислого газа связываются по одной, присоединяются к уже готовому пятиуглеродному органическому веществу. Образуется неустойчивое шестиуглеродное органическое вещество, которое распадается на трехуглеродные молекулы углевода. Часть этих молекул используется на ресинтез исходного пятиуглеродного вещества для связывания CO2. Такой ресинтез обеспечивается циклом Кальвина. Меньшая часть молекул углевода, включающего три атома углерода, выходит из цикла. Уже из них и других веществ синтезируются все остальные органические вещества (углеводы, жиры, белки).
То есть на самом деле из темновой фазы фотосинтеза выходят трехуглеродные сахара, а не глюкоза.
Источник: scienceland.info
Роль хлоропластов в фотосинтезе
В клетках растений имеются микроскопические образования — хлоропласты. Это органоиды, в которых происходит поглощение энергии и света и превращение ее в энергию АТФ и иных молекул — носителей энергии. В гранах хлоропластов содержится хлорофилл — сложное органическое вещество. Хлорофилл улавливает энергию света для использования ее в процессах биосинтеза глюкозы и других органических веществ. Ферменты, необходимые для синтеза глюкозы, расположены также в хлоропластах.
Световая фаза фотосинтеза
Квант красного света, поглощенный хлорофиллом, переводит электрон в возбужденное состояние. Возбужденный светом электрон приобретает большой запас энергии, вследствие чего перемещается на более высокий энергетический уровень. Возбужденный светом электрон можно сравнить с камнем, поднятым на высоту, который также приобретает потенциальную энергию. Он теряет ее, падая с высоты. Возбужденный электрон, как по ступеням, перемещается по цепи сложных органических соединений, встроенных в хлоропласт. Перемещаясь с одной ступени на другую, электрон теряет энергию, которая используется для синтеза АТФ. Растративший энергию электрон возвращается к хлорофиллу. Новая порция световой энергии вновь возбуждает электрон хлорофилла. Он снова проходит по тому же пути, расходуя энергию на образования молекул АТФ. Ионы водорода и электроны, необходимые для восстановления молекул-носителей энергии, образуются при расщеплении молекул воды. Расщепление молекул воды в хлоропластах осуществляется специальным белком под воздействием света. Называется этот процесс фотолизом воды. Таким образом, энергия солнечного света непосредственно используется растительной клеткой для: 1. возбуждения электронов хлорофилла, энергия которых далее расходуется на образование АТФ и других молекул-носителей энергии; 2. фотолиза воды, поставляющего ионы водорода и электроны в световую фазу фотосинтеза. При этом выделяется кислород как побочный продукт реакций фотолиза. Этап, в течение которого за счет энергии света образуются богатые энергией соединения — АТФ и молекулы-носители энергии, называют световой фазой фотосинтеза.
Темновая фаза фотосинтеза
В хлоропластах есть пятиуглеродные сахара, один из которых рибулозодифосфат, является акцептором углекислого газа. Особый фермент связывает пятиуглеродный сахар с углекислым газом воздуха. При этом образуется соединения, которые ща счет энергии АТФ и иных молекул-носителей энергии восстанавливаются до шестиуглеродной молекулы глюкозы. Таким образом, энергия света, преобразованная в течение световой фазы в энергию АТФ и иных молекул-носителей энергии, используется для синтеза глюкозы. Эти процессы могут идти в темноте. Из растительных клеток удалось выделить хлоропласты, которые в пробирке под действием света осуществляли фотосинтез — образовывали новые молекулы глюкозы, при этом поглощали углекислый газ. Если прекращали освещать хлоропласты, то приостанавливался и синтез глюкозы. Однако если к хлоропластам добавляли АТФ и восстановленные молекулы-носители энергии, то синтез глюкозы возобновлялся и мог идти в темноте. Это означает, что свет действительно нужен только для синтеза АТФ и зарядки молекул-носителей энергии. Поглощение углекислого газа и образование глюкозы в растениях называют темновой фазой фотосинтеза, поскольку она может идти в темноте. Интенсивное освещение, повышенное содержание углекислого газа в воздухе приводят к повышению активности фотосинтеза.
Источник: edu.glavsprav.ru