Темная фаза фотосинтеза
Темновая фаза фотосинтеза
Темновая фаза фотосинтеза – это комплекс химических реакций, в результате которых происходит восстановление поглощенного листом СО2 за счет продуктов световой фазы (АТФ и НАДФ) и образуются органические вещества.
АТФ и НАДФ, образовавшиеся в световой фазе, и используемые для восстановления СО2, получили название ассимиляционной силы.

Цикл Кальвина (С-3 цикл)
3 РуБФ + 3 СО2 + 6 НАДФН + 9 АТФ ![]() 3 РуБФ + 3 ФГА + 3 Н2О + НАДФ+ 9 АДФ + 3Н3РО4
3 РуБФ + 3 ФГА + 3 Н2О + НАДФ+ 9 АДФ + 3Н3РО4
Происходит в строме хлоропластов
Этапы цикла Кальвина:
Карбоксилирующий – присоедниение 3 молекул СО2 к рибулозо-1,5-бифосфату (РуБФ).
(Фермент- рибулозодифосфаткарбоксилаза)
2. Восстановительный – восстановление ФГК с помощью ассимиляционной силы с образованием ФГА.
3. Регенерирующий – восстановление молекулы РуБФ.
РуБФ и ФГА – это конечные продукты цикла Кальвина. Все остальные вещества, участвующие в нем, называют промежуточными.
Т. к. в цикле Кальвина образуются пентозы, его называют еще восстановительным пентозофосфатным циклом (ВПФ-цикл).
3 РуБФ + 3 СО2 + 6 НАДФН + 9 АТФ ![]() 3 РуБФ + 3 ФГА + 3 Н2О + НАДФ+ 9 АДФ + 3Н3РО4
3 РуБФ + 3 ФГА + 3 Н2О + НАДФ+ 9 АДФ + 3Н3РО4
Цикл Хетча – Слэка и С4 – растения
Австралийские ученые и в 1967 г. обнаружили, что у кукурузы, сахарного тростника, сорго и некоторых др. тропических растений первичным продуктом темновой фазы не ФГК, а оксалоацетат.
Кроме того, эти растения имеют не только гранальные, но и агранальные хлоропласты.
В результате дальнейших исследований был открыт еще один цикл фотосинтеза, названный циклом Хетча – Слэка или С4 – циклом.
Первичный продукт темновой фазы – оксалоацетат – содержит в своей молекуле 4 атома углерода.
Акцептором СО2 в этом цикле является фосфоенолпируват (ФЕП)
В результате карбоксилирования ФЕП образуется оксалоацетат и ортофосфат.
ФЕП + СО2 + Н2О
![]() оксалоацетат + Н3РО4
оксалоацетат + Н3РО4
(фермент: ФЕП-карбоксилаза. Она локализована в цитозоле или на внешней мембране хлоропласта).
Образование оксалоацетата происходит вне гранального хлоропласта.
Образовавшийся оксалоацетат поступает в хлоропласт и здесь при участии НАДФН восстанавливается до малата:
Оксалоацетат + НАДФН + Н+ ![]() малат + НАДФ+
малат + НАДФ+
фермент: НАДФ+- зависимая малатдегидрогеназа
У некоторых растений оксалоацетат превращается в аспартат:
Оксалоацетат + NH3 + НАДФН ![]() аспартат + Н2О
аспартат + Н2О
фермент: аспартатаминотрансфераза
Малат или аспартат транспортируются из клеток мезофилла в хлоропласты обкладки проводящих пучков.
В зависимости от того, какая кислота — малат или аспартат – транспортируется в клетки обкладки, растения делят на два типа: малатный или аспартатный.
В клетках обкладки эти кислоты деркарбоксилируются.
Окислительное декарбоксилирование малата в агранальных хлоропалстах клеток обкладки:
Малат + НАДФ+
![]() Пируват + СО2_+ НАДФН + Н+
Пируват + СО2_+ НАДФН + Н+
Окислительное декарбоксилирование малата в агранальных хлоропалстах клеток обкладки:
Малат + НАДФ+ ![]() Пируват + СО2_+ НАДФН + Н+
Пируват + СО2_+ НАДФН + Н+
СО2_используется для карбоксилирования РуБФ
(вторичное карбоксилирование), т. е. включается в С3 –цикл, идущий в агранальных хлоропластах клеток обкладки.
Пируват возвращается в хлоропласты клеток мезофилла, где он фосфорилируется за счет АТФ – продукта световой фазы, что приводит к регенерации ФЕП – акцептора СО2 и замыканию цикла:
ПВК + АТФ + ФН ![]() ФЕП + АМФ + ФФН
ФЕП + АМФ + ФФН
Представителями растений этой группы являются сахарный тростник, кукуруза, сорго
Особенности С4 – цикла
1. Из оксалоацетата – первичного продукта – образуются малат или аспартат;
2. Цикл делят на два этапа:
1 – карбоксилирование ФЕП в клетках мезофилла,
2 – декарбоксилирование малата или аспатрата в клетках обкладки.
3. Цикл разделен в пространстве: начинается и кончается в основных клетках мезофилла, а декарбоксилирование происходит в клетках обкладки.
С3 – и С4 – циклы действуют совместно. Такое совместное функционирование двух циклов получило название кооперативного фотосинтеза.
Основная функция С4 – цикла — концентрирование СО2 для С3 – цикла.
С4 – цикл является своеобразным насосом (углекислотной помпой) для С3 – цикла, с помощью которого СО2 атмосферы переносится в С3 – цикл.
С3 – растения – большинство сельскохозяйственных растений;
С4 – растения – преимущественно растения тропиков и субтропиков. Они очень урожайны.
В условиях засухи и высоких температур у них фотосинтез идет с высокой интенсивностью.
Большинство самых злостных сорняков на Земле — С4 – растения.
В неблагоприятных условиях у некоторых С3 – растений начинает работать и С4 – цикл.
Благодаря функционированию углекислотной помпы (С4 – цикла) концентрация СО2 в клетках обкладки, где идет С3 –цикл, в несколько раз больше, чем в окружающей среде.
Это очень важно, т. к. С4 – растения живут в условиях повышенных температур, когда растворимость СО2 значительно снижена.
В листьях С4 – растений карбоксилирование происходит дважды.
Особенности анатомического строения листьев
С4 – растений
1. Большое количество проводящих пучков, что помогает быстрому оттоку ассимилятов из листа.
2. В клетках мезофилла листа находятся гранальные хлоропласты.
В клетках обкладки – более крупные агранальные хлоропласты.
В гранальных хлоропластах основных клеток мезофилла происходит первичное карбоксилирование ФЕП и его регенерация из ПВК или аланина, т. е. здесь начинается и кончается С4 – цикл.
В агранальных хлоропластах клеток обкладки идет декабоксилирование С4 – кислот и С3 –цикл
Гранальные хлоропласты основных клеток мезофилла содержат Фотосистему I (ФС I) и Фотосистему II (ФС II), поэтому в них идет циклическое и нециклическое фотофосфорилирование с образованием АТФ и НАДФН.
Агранальные хлоропласты клеток обкладки содержат Фотосистему I (ФС I) и в них возможно только циклическое фотофосфорилирование. В этих клетках НАДФ не восстанавливается.
С4 – цикл поставляет в С3 –цикл РІРѕРґРѕСЂРѕРґ для восстановления СО2.
Для нормального течения темновой фазы фотосинтеза необходимы строгая координация скоростей С3 – и С4 – циклов и быстрый отток ассимилятов.
В процессе эволюции С3 – цикл возник раньше, чем С4 – цикл.
У древесных растений (более старая жизненная форма) С4 – цикл не обнаружен.
Кислотный метаболизм толстянковых
Crassulacean acid metabolism
(САМ-фотосинтез) Этот тип фотосинтеза впервые был обнаружен у растений из семейства толстянковых (Crassulaceaе).
Растения, имеющие такой тип фотосинтеза назвали растениями САМ-типа.
Растения САМ-типа – это суккуленты, растущие в сухих местах, у которых устьица открыты ночью и закрыты днем для уменьшения транспирации.
Поэтому эти растения поглощают СО2 ночью.
Особенности САМ-фотосинтеза
1. Темновая фаза фотосинтеза разделена во времени: СО2 поглощается ночью, а восстанавливается днем (суккуленты днем закрывают устьица для уменьшения транспирации).
2. Из оксалоацетата, как и у С4-растений, образуется малат.
3. Карбоксилирование в тканях происходит дважды: ночью карбоксилируется ФЕП, днем – РуБФ.
Значение САМ-фотосинтеза
Как и С4- цикл, водосберегающий САМ-тип фотосинтеза, является дополнительным, поставляющим СО2 в С3- цикл растений, приспособившихся к жизни в условиях повышенных температур или недостатка влаги.
Днем в жару САМ-растения запасают СО2 , образующийся в результате дыхания, а ночью, когда устьица открыты, они поглощают СО2 из воздуха.
САМ-фотосинтез помогает уменьшить транспирацию и запасать воду, но продуктивность этих растений намного ниже, чем у С3-растений.
Особенности клеток САМ-растений
Клетки мезофилла у САМ-растений в несколько раз крупнее, т. к. у них больше вакуоли.
Число устьиц намного больше, чем у С3 и С4-растений.
Гликолатный цикл фотосинтеза
В 1779 г. Я. Ингенхауз обнаружил, что иногда днем, при очень высокой освещенности и температуре, растения выделяют СО2 и поглощают О2 .
Через 150 лет об этом явлении вспомнили и назвали его фотодыханием (по типу газообмена, активирующегося на свету в зеленых клетках).
В 60-е годы ХХ века начали изучать фотодыхание. Оказалось, что большую роль в этом процессе играет РуБФ-карбоксилаза. Если мало СО2 и много О2, то она может присоединять к РуБФ не СО2, а О2, т. е. РуБФ-карбоксилаза может выполнять не только карбоксилирующую, но и оксигенирующую функции.
В хлоропласте:
РуБФ + О2 + АТФ ![]() ФГК + фосфогликолат + АДФ
ФГК + фосфогликолат + АДФ
Выходя из хлоропласта, фосфогликолат дефосфорилируется
фосфогликолат ![]() гликолат + Н3РО4
гликолат + Н3РО4
(фермент – фосфогликолатфосфатаза)
и превращается в гликолат, который поступает в пероксисому
В пероксисоме:
Гликолат + О2 ![]() глиоксилат + Н2О2
глиоксилат + Н2О2
(фермент – гликолатоксидаза)
Н2О2
![]() Н2О + О2
Н2О + О2
(фермент – каталаза)
Глиоксилат + NH3 ![]() глицин
глицин
(фермент – глиоксилатглицинаминотрансфераза)
Глицин транспортируется в митохондрию
В митохондрии:
2 Глицин ![]() серин + СО2 + NH3 + НАДН + Н+
серин + СО2 + NH3 + НАДН + Н+
(фермент – серинглиоксилатаминотрансфераза)
Серин транспортируется из митохондрии в пероксисому.
В пероксисоме:
серин ![]() NH3 + оксипируват
NH3 + оксипируват
Оксипируват глицерат
Глицерат транспортируется в хлоропласт
В хлоропласте:
Глицерат + АТФ ![]() ФГК + АДФ
ФГК + АДФ
ФГК является конечным продуктом этого цикла.
ФГК может поступить в С3 – цикл для регенерации РуБФ или использоваться для синтеза сахарозы или крахмала.
Гликолат считается первичным продуктом этого цикла, поэтому цикл превращениия его в ФГК (триозофосфат) назвали гликолатным
Особенности гликолатного цикла фотосинтеза
СО2 образуется во время превращения двух молекул глицина в серин;
О2 расходуется для синтеза гликолата и глиоксилата;
Во время этого цикла, как и при дыхании, поглощается О2 и выделяется СО2;
В течение цикла образуется свободный аммиак NH3, который используется для аминирования оксиглутарата, в результате образуется глутамат;
ФГК может использоваться для синтеза сахарозы или крахмала.
Было доказано, что гликолатный цикл фотосинтеза есть у С3 – растений.
У САМ – растений устьица днем закрыты, поэтому СО2 не выделяется в атмосферу, и этот цикл трудно обнаружить.
У С4 – растений гликолатный цикл есть только в клетках обкладки и идет с небольшой интенсивностью. В клетках мезофилла листа нет РуБФ-карбоксилазы.
В клетках обкладки не происходит фотолиз воды из-за отсутствия в их хлоропластах ФС II.
У С4 – растений очень высокая продуктивность.
Функции гликолатного цикла фотосинтеза
1. Гликолатный цикл, как и другие циклы фотосинтеза, источником промежуточных веществ для других синтезов.
2. Гликолатный цикл играет важную роль в образовании аминокслот – серина и глицина, т. е. связан с азотным обменом растения.
3. При образовании серина из глицина восстанавливантся митохондриальный НАД, при окислении которого образуется АТФ.
4. Избыток продуктов С3 – цикла увеличивает скорость гликолатного цикла, а их недостаток усиливает интенсивность С3 – цикла.
Следовательно, соотношение гликолатного цикла и С3 – цикла помогает поддерживать гомеостаз в растительном организме
Общая характеристика фотосинтеза
Суть фотосинтеза состоит в фотоокислении воды с выделением О2 и использованием Н+ на восстановление СО2 с образованием органических веществ;
Фотосинтез состоит из двух фаз: световой и темновой;
Энергия поглощенного света используется для возбуждения молекул П700 и П680 и транспорта электронов;
Транспорт электронов электронов сопряжен с синтезом АТФ;
Световая фаза проходит в тилакоидах стромы и гран, а темновая — в строме хлоропласта.
Источник: pandia.ru
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + H2O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6H2O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: biology.su
Темновая фаза фотосинтеза связана с реакциями фиксации углерода, которые проходят в строме хлоропласта и продолжаются в цитоплазме без непосредственного поглощения света. В процессе световой фазы фотосинтеза накапливается достаточно высокий уровень АТФ и НАДФ·Н. Однако сами по себе эти макроэргические соединения не способны синтезировать углеводы из CO2. Становится очевидным, что и темновая фаза фотосинтеза – сложный процесс, включающий большое количество последовательно идущих реакций, возможных только после осуществления световой фазы.
Существует несколько разных путей связывания CO2 в углеводы, встречающихся у растений разных экологических и систематических групп, но основным, характерным для всех растений, является так называемый C3-путь фотосинтеза, или цикл Кальвина.
Способ ассимиляции СО2 в углеводы, присущий всем растениям, был расшифрован только в середине XX века американским биохимиком Мэлвином Кальвином и его коллегами на примере одноклеточных зеленых водорослей (хлореллы и др.) и зеленых листьев шпината. Исследование этой проблемы продолжалось 10 лет – с 1946 по 1966 год. Вначале ученые вели поиск первичного акцептора CO2. После ряда экспериментов они установили, что первичную фиксацию CO2 осуществляет пятиуглеродный сахар – рибулозо-1,5-дифосфат РуДФ). Фиксация осуществляется следующим образом: сначала происходит присоединение CO2 к молекуле РуДФ. При этом образуется промежуточный продукт – очень неустойчивое шестиуглеродное соединение, из которого в присутствии воды образуются две молекулы трехуглеродного соединения – 3-фосфоглицериновой кислоты (3-ФГК). В этой реакции для связывания одной молекулы CO2 затрачивается три молекулы АТФ и две молекулы НАДФ·Н.
Схема первичной фиксации CO2
Реакцию фиксации углерода, открытую в 1948 году, катализирует очень крупный фермент из стромы хлоропласта – рибулозобисфосфаткарбоксилазаоксигеназа (сокращенно – РУБИСКО). Так как фермент РУБИСКО работает весьма медленно, необходимо, чтобы его молекул в хлоропластах было много. Действительно, этот фермент обычно составляет более 50 % общего количества белков хлоропластов. Многие исследователи утверждают, что это самый распространенный белок в живой природе.
Дальнейшие исследования лаборатории Кальвина способствовали установлению всех последующих реакций C3-пути фотосинтеза, обеспечивающих синтез углеводов. За расшифровку механизма фиксации CO2 в процессе фотосинтеза М. Кальвин в 1961 году стал лауреатом Нобелевской премии по химии.
Цикл Кальвина состоит из трех стадий:
- карбоксилирования,
- восстановления,
- превращения.
Упрощенная схема цикла Кальвина – пути фиксации углерода при фотосинтезе
На первой стадии (карбоксилирование) фиксация углерода идет с участием ферментов и АТФ, полученной на световой фазе фотосинтеза; при этом образуются молекулы 3-фосфоглицериновой кислоты (3-ФГК). На второй стадии (восстановление) помимо АТФ используется НАДФ·Н. Здесь 3-ФГК восстанавливается до 3-фосфоглицеринового альдегида (З-ФГА), часть молекул которого идет на синтез 6-углеродного моносахарида (глюкозы или фруктозы). На третьей стадии (превращение) при повторении цикла другая часть молекул 3-ФГА используется для синтеза шестиатомного фосфорилированного моносахарида – фруктозо-1,5-дифосфата. Трехуглеродные фосфосахара вместе с множеством других продуктов метаболизма хлоропластов транспортируются в цитоплазму клетки, где образуют ди- и полисахариды (сахара, крахмал, целлюлозу или другие соединения).
В процессах темновой фазы фотосинтеза образуются углеводы – первичные органические вещества. На определенном этапе темновой фазы фотосинтеза судьба трехуглеродных молекул 3-фосфоглицериновой кислоты может оказаться различной. Одни из них соединяются друг с другом и образуют шестиуглеродные сахара, которые, в свою очередь, могут полимеризоваться в крахмал, целлюлозу и др. Некоторые могут использоваться для синтеза аминокислот, карбоновых кислот, спиртов и пр. Но целый ряд молекул ФГК вовлекается в длинный ряд реакций, приводящих к превращению трехуглеродных молекул в молекулы пятиуглеродного сахара (РуДФ), которые могут снова ассимилировать углекислый газ и многократно повторять этот цикл до тех пор, пока растение живет и получает световую энергию. Все процессы темновой фазы фотосинтеза идут без непосредственного потребления света, но в них большую роль играют макроэргические соединения (АТФ и НАДФ·Н), образующиеся во время световой фазы фотосинтеза. Доказано, что для синтеза одной молекулы глюкозы в цикле Кальвина необходимы 12 молекул НАДФ·Н и 18 молекул АТФ, которые поставляются с тилакоидных мембран в результате фотохимических реакций световой фазы фотосинтеза.
Схема синтеза 3-фосфоглицеринового альдегида (ФГА) в хлоропласте
В процессе темновой фазы энергия макроэргических связей АТФ преобразуется в химическую энергию органических соединений – молекул углеводов. Это означает, что энергия солнечного света как бы консервируется в химических связях между атомами органических веществ, что имеет огромное значение для энергетики биосферы и жизнедеятельности всего населения нашей планеты.
В настоящее время известны и другие пути ассимиляции углекислого газа наряду с системой его фиксации в цикле Кальвина (C3-пути фотосинтеза). Существует так называемый C4-путь ассимиляции углерода в фотосинтезе. Он может протекать при низких концентрациях CO2. Этот тип фиксации углекислого газа в фотосинтезе выработался в процессе эволюции у растений жарких, засушливых мест и наблюдается у кукурузы, сахарного тростника, проса, сорго, амаранта, лебеды, баклажанов и др., а также у растений, устойчивых к засолению почвы.
Существует и особый тип фотосинтеза у таких растений, как кактусы, молочаи, крассулы, каланхое, седумы и другие суккуленты, произрастающие в засушливых, безводных условиях. Эти растения запасают CO2 в виде органических кислот ночью, так как он поступает в клетки только тогда, когда открыты их устьица (днем они закрыты для предотвращения потери воды).
Источник: vseobiology.ru
С3-путь фотосинтеза (цикл кальвина)
Этот способ ассимиляции СО2, присущий всем растениям, был расшифрован американскими исследователями во главе с Мэльвином Кальвиным. Схема последовательности реакций получила название цикла Кальвина. Характерной особенностью фотосинтетического восстановления СО2 являются цикличность и разветвленность этого процесса. Цикличность обеспечивает высокую производительность, саморегуляцию и непрерывность образования углеводов. Разветвленность — образование разнообразных продуктов, дублирование путей регенерации акцептора СО2. Цикл состоит из трех этапов: карбоксилирования, восстановления и регенерации акцептора СО2 (рис.).
Карбоксилирование. Первой реакцией, вводящей СО2 в цикл Кальвина, является карбоксилирование рибулезо-1.5-дифосфата (1,5-РДФ) с участием фермента рибулезодифосфаткарбоксилазы (РДФ-карбоксилазы). Образующееся при этом нестойкое щестиутлеродное соединение быстро распадается на триозы — две молекулы 3-фосфоглицериновой кислоты (З-ФГК). Поэтому З-ФГК можно считать первичным продуктом фотосинтеза.
Фаза восстановления. Восстановление 3-фосфоглицериновой кислоты (З-ФГК) до 3-фосфоглицеринового альдегида (3-ФГА) происходит в два этапа. Сначала при участии АТФ и фосфоглицераткиназы З-ФГК присоединяет остаток фосфорной кислоты с образованием 1,3-дифосфоглицериновой кислоты (1,3-ДФГК.) Этим достигается повышение реакционной способности соединения и возможность его восстановления с помощью НАДФ-Н до З-ФГА. Это единственная восстановительная реакция цикла. Фаза восстановления является центральным звеном цикла. Именно здесь скрещиваются световая и темновая фазы фотосинтеза. Все остальные превращения идут на уровне сахаров, одинаковых по степени восстановленности.
Фаза регенерации первичного акцептора СО2 и синтеза конечных продуктов фотосинтеза. В результате рассмотренных ранее реакций при фиксации трех молекул СО2 образуются 6 молекул восстановленных 3-фосфотриоз, пять из них используются затем для регенерации РДФ, а 1- для синтеза глюкозы. Это достигается следующей последовательностью реакций. Часть молекул З-ФГА под действиеч триозофосфатизомеразы превращается в фосфодиоксиацетон (ФДА). Затем два изомера (ФГА и ФДА) полвергаются конденсации при участии фермента альдолазы, образуя фруктоза-1,6-дифосфат, у которого затем отщепляется один фосфат.
В дальнейших реакциях, связанных с регенерацией акцептора СО2, возникает цепь фосфорных эфиров сахаров, содержащих в своем составе 4, 5 или 7 атомов углерода. В этих реакциях последовательно принимают участие транскетолазы и трансальдолазы. Транскетолаза катализирует перенос 2-углеродной группировки от фруктоза-6-фосфата на З-ФГА, при этом образуются эритрозо-4-фосфат (С4) и ксилулозо-5-фосфат (С5). Затем альдолаза осуществляет перенос 3-углеродного остатка ФДА на эритрозо-4-фосфат, в результате чего синтезируется седогептулозо-1.7-дифосфат (С7). От последнего отщепляется один остаток фосфорной кислоты и под действием транскетолазы из него и З-ФГА образуются ксилулозо-5-фосфат (С5) и рибозо-5-фосфат (С5). Две молекулы ксилулозо-5-фосфата путем эпимеризации и одна молекула рибозо-5-фосфата за счет изомеризации превращаются в три молекулы рибулозо-5-фосфата (С5). Это соединение подвергается фосфорилированию за счет АТФ, образующейся в световую фазу.
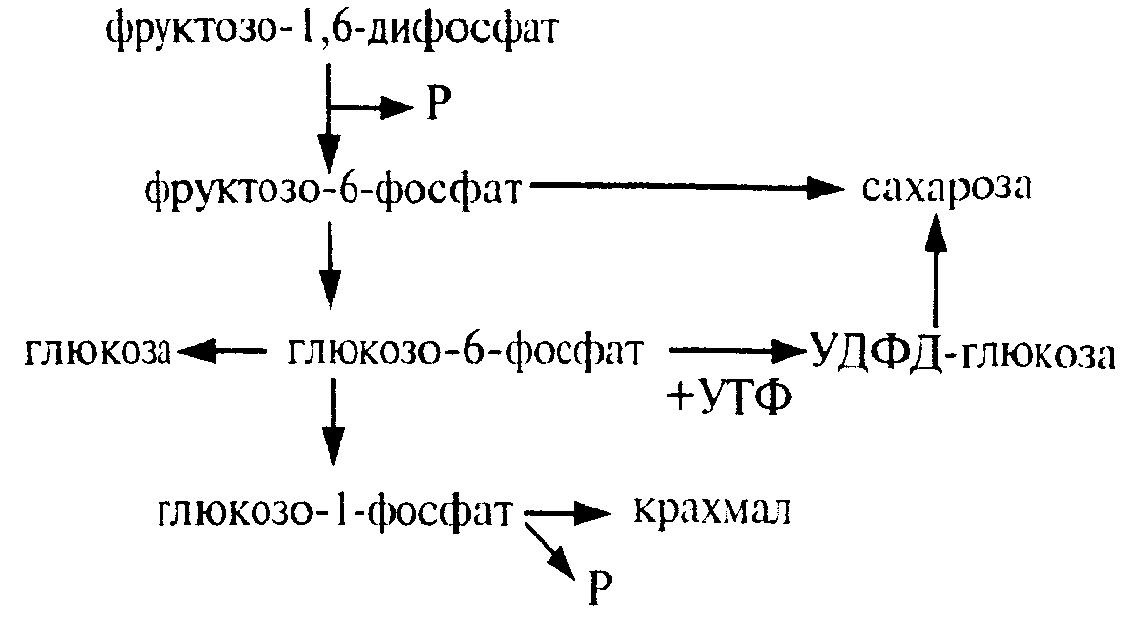
Из оставшейся неиспользованной молекулы ФГА при повторении цикла образуется фруктоза-1,6-дифосфат, из которой могут образовываться глюкоза, сахароза и крахмал.
Таким образом, для синтеза одной молекулы ГЛЮКОЗЫ (С6) должно произойти шесть оборотов цикла. В каждом обороте используются 3 мол. АТФ (две для активирования двух молекул ФГК и одна при регенерации акцептора СО2 1,5-РДФ) и две молекулы НАДФН для восстановления ФГК до ФГА. Поэтому для синтеза одной молекулы глюкозы в цикле Кальвина необходимо 12 НАДФН и 18 АТФ. Таков энергетический вклад световой фазы в темновую фиксацию СО2.
В цикле Кальвина первичными продуктами включення СО2 в органические вещества н восстановления являются трехуглеродные соединения (З-ФГК, З-ФГА, ФДА). Поэтому этот способ фиксации СО2 носит название С3-пути фотосинтеза. Большинство растений, особенно произрастающих в умеренной зоне, используют именно этот путь восстановления СО2 и называются С3-растениями.
Источник: StudFiles.net