Функции красного ядра
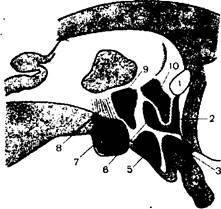
Средний мозг входит в состав ствола мозга. С вентральной стороны к нему примыкает задняя поверхность сосцевидных тел и передний край моста сзади (Атл., рис. 23, с. 133). В нем выделяют крышу и ножки. Полостью среднего мозга является водопровод мозга — узкий канал, длиной около 1,5 см, который снизу сообщается с четвертым желудочком, а сверху — с третьим.
Крыша среднего мозга представляет собой пластинку четверохолмия и расположена над водопроводом мозга. Крыша среднего мозга состоит из четырех возвышений — холмиков, которые отделены друг от друга двумя бороздками — продольной и поперечной.
В плоской канавке между верхними бугорками лежит шишковидное тело. Каждый холмик переходит в так называемую ручку холмика, направляющуюся латерально, кпереди и кверху, к промежуточному мозгу. Ручка верхнего холмика направляется к латеральному коленчатому телу; ручка нижнего холмика — к медиальному коленчатому телу.
Верхние два холмика крыши среднего мозга и латеральные коленчатые тела являются подкорковыми центрами зрения. Оба нижних холмика и медиальные коленчатые тела — подкорковыми центрами слуха.
От крыши среднего мозга берет начало тектоспинальный путь. Его волокна после перекреста в покрышке среднего мозга идут к двигательным ядрам головного и клеткам передних рогов спинного мозга. Путь проводит эфферентные импульсы в ответ на зрительные и слуховые раздражения.
Ножки мозга занимают переднюю часть среднего мозга, расположены под мостом и направляются к правому и левому полушариям переднего мозга. Углубления между правой и левой ножками получило название межножковой ямки. Ножки состоят из основания и покрышки, которые разделяются пигментированными клетками черной субстанции.
В основании ножек проходит пирамидный путь, направляющийся через мост в спинной мозг и корково-ядерный, волокна которого доходят до нейронов двигательных ядер черепных нервов, расположенных в области четвертого желудочка и водопровода, а также корково-мостовой путь оканчивающийся на клетках основания моста. Следовательно, основания ножек мозга целиком состоят из белого вещества, здесь проходят нисходящие проводящие пути. Покрышка ножек продолжает покрышку моста и продолговатого мозга. Верхняя ее поверхность служит дном водопровода мозга. В покрышке расположены ядра блокового (IV) и глазодвигательного (III) нервов, и проходят восходящие проводящие пути.
В области III пары нервов лежит парасимпатическое ядро; оно состоит из вставочных нейронов автономной нервной системы. В верхней части покрышки среднего мозга проходит дорсальный продольный пучок, связывающий таламус и гипоталамус с ядрами ствола мозга.
Среди ядер серого вещества выделяются черная субстанция и красное ядро. Черная субстанция разделяет основание и покрышку ножек мозга. Ее клетки содержат пигмент меланин. Этот пигмент существует только у человека и появляется в возрасте 3—4 лет. Черная субстанция получает импульсы от коры головного мозга, полосатого тела и мозжечка и передает их нейронам верхнего двухолмия и ядрам ствола, а далее — на мотонейроны спинного мозга. Черная субстанция играет существенную роль в интеграции всех движений и в регуляции пластического тонуса мышечной системы.
Красное ядроявляется самым крупным ядром покрышки и располагается несколько выше (дорсальнее) черного вещества. Оно имеет удлиненную форму и простирается от уровня нижних холмиков до таламуса. На уровне нижнего двухолмия совершается перекрест верхних ножек мозжечка. Большая их часть заканчивается на красных ядрах, а меньшая часть проходит сквозь красное ядро и продолжается к таламусу. В красном ядре оканчиваются волокна из больших полушарий. От его нейронов идут восходящие пути, в частности к таламусу. Основной нисходящий путь красный ядер — руброспинальный (красноядерно-спинно-мозговой). Его волокна сразу по выходе из ядра совершают перекрест, направляются вдоль покрышек ствола головного мозга и бокового канатика спинного мозга к мотонейронам передних рогов спинного мозга.
Латеральнее красного ядра в покрышке расположена медиальная петля. Между ней и серым веществом, окружающим водопровод, лежат нервные клетки и волокна ретикулярной формации (продолжение ретикулярной формации моста и продолговатого мозга) и проходят восходящие и нисходящие пути.
Функции среднего мозга. Средний мозг выполняет сенсорные функции, проводниковую, двигательную и рефлекторные функции.
Сенсорные функции осуществляются за счет поступления в средний мозг зрительной, слуховой информации. Верхние холмики четверохолмия являются первичными подкорковыми центрами зрительного анализатора (вместе с латеральными коленчатыми телами промежуточного мозга), нижние — слухового (вместе с медиальными коленчатыми телами промежуточного мозга). В них происходит первичное переключение зрительной и слуховой информации.
Проводниковая функция заключается в том, что через средний мозг проходят все восходящие пути к вышележащим отделам ЦНС: таламусу (медиальная петля, спинно-таламический путь), переднему мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу. К ним относятся пирамидный путь, корково-мостовые волокна, руброретикулоспинальный путь.
Двигательная функция реализуется за счет блокового нерва, ядер глазодвигательного нерва, красного ядра, черной субстанции. Красное ядро и окружающие его двигательные ядра имеют важное значение для осуществления всех движений, так как они рефлекторно регулируют тонус мускулатуры. Базальные ганглии головного мозга, мозжечок имеют свои окончания в красных ядрах. Нарушение связей красных ядер с ретикулярной формацией продолговатого мозга ведет к децеребрациальной ригидности. Это состояние характеризуется сильным напряжением мышц-разгибателей конечностей, шеи, спины. Основной причиной возникновения децеребрациальной ригидности служит выраженное активирующее влияние латерального вестибулярного ядра (ядро Дейтерса) на мотонейроны разгибателей. При перерезке мозга ниже ядра латерального вестибулярного нерва децеребрациальная ригидность исчезает.
Красные ядра, получая информацию от двигательной зоны коры больших полушарий, подкорковых ядер и мозжечка о готовящемся движении, посылают корригирующие импульсы к мотонейронам спинного мозга по руброспинальному пути и тем самым регулируют тонус мускулатуры, подготавливая его уровень к произвольному движению.
Черная субстанция регулирует акты жевания, глотания (их последовательность), обеспечивает точные движения пальцев кисти рук, например, при письме. Нейроны этого ядра способны синтезировать медиатор дофамин, который по аксонам поступает к базальным ганглиям головного мозга. Поражение черного вещества приводит к нарушению пластического тонуса мышц и связано с невралгическим заболеванием — болезнью Паркинсона. Паркинсонизм проявляется в нарушении тонких содружественных движений, функции мимической мускулатуры и в проявлении непроизвольных мышечных сокращений, или тремора.
Тонкая регуляция пластического тонуса при игре на скрипке, письме, выполнении графических работ обеспечивается черным веществом. В то же время при длительном удержании определенной позы происходят пластические изменения в мышцах, чтообеспечивает наименьшие затраты энергии. Регуляция этого процесса обеспечивается клетками черной субстанции.
Нейроны ядер глазодвигательного и блокового нервов регулируют движения глаз вверх, вниз, к носу и вниз к углу носа. Нейроны добавочного ядра глазодвигательного нерва (ядро Якубовича) регулируют просвет зрачка и кривизну хрусталика.
Рефлекторные функции.Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Их основная функция заключается в организации реакций настараживания и так называемых старт-рефлексов на внезапные, еще не распознанные зрительные или звуковые сигналы. Активация среднего мозга в этих случаях через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию, к оборонительной реакции.
Четверохолмие организует ориентировочные зрительные и слуховые рефлексы. У человека этот рефлекс является сторожевым. В случаях повышенной возбудимости четверохолмий при внезапном звуковом или световом раздражении у человека возникает вздрагивание, иногда вскакивание на ноги, вскрикивание, максимально быстрое удаление от раздражителя, подчас безудержное бегство.
При нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое. Следовательно, четверохолмия принимают участие в организации произвольных движений.
Развитие среднего мозга.Рост и функциональное развитие среднего мозга связано с развитием других отделов ствола мозга и формированием его путей к мозжечку и коре больших полушарий головного мозга.
У новорожденного масса среднего мозга составляет 2,5 г. Его форма и строение не отличаются от таковых у взрослого. Водопровод мозга более широк, глазодвигательный нерв имеет миелинизированные волокна. Черное вещество и ретикулярная формация распространяются по длине среднего мозга до бледного шара. Их клетки хорошо дифференцированы, но не содержат пигмента, его появление приходится на шестой месяц жизни и иногда почти к периоду полового созревания. Максимального развития они достигают около 16 лет. Развитие пигментации находится в прямой связи с совершенствованием функции черной субстанции. Медиальная часть черного вещества начинает миелинизироваться в первые 2—3 месяца жизни.
Красное ядро хорошо выражено, его связи с другими отделами мозга формируются раньше, чем пирамидная система. У новорожденного пирамидные волокна миелинизированы, а пути, идущие к коре, не имеют к этому периоду миелиновой оболочки. Они миелинизируются с 4-го месяца жизни. Медиальная петля, а также волокна, связывающие красное ядро и черное вещество, миелинизированы.
Пигментация красного ядра начинается с 2-летнего возраста и заканчивается к 4 годам.
Функциональное развитие среднего мозга.Ряд рефлексов, осуществляющихся с участием среднего мозга, формируется в период внутриутробного развития. Уже на ранних этапах эмбриогенеза отмечены тонические и лабиринтные рефлексы, оборонительные и другие двигательные реакции в ответ на различные раздражения.
За 2—3 месяца до рождения у плода наблюдаются двигательные реакции в ответ на звуковые, температурные, вибрационные и другие раздражения.
В первые дни жизни ребенка появляется рефлекс Моро, который выражается в том, что в ответ на громкий внезапный звук у ребенка разгибаются руки в сторону под прямым углом к туловищу, разгибаются пальцы и туловище. Этот рефлекс исчезает к 4-му году жизни ребенка. Он сохраняется у умственно отсталых детей, и его считают связанным с незрелостью мозга.
Рефлекс Моро сменяется противоположной реакцией. Так, например, при таком же резком раздражении у ребенка возникает общая двигательная реакция с преобладанием сгибательных движений. Она нередко сопровождается движением головы и глаз, изменением дыхания или задержкой сосательного рефлекса. Эта реакция названа реакцией испуга или вздрагиванияи рассматривается как первое проявление ориентировочного рефлекса.
При повторных раздражениях этот рефлекс исчезает. С возрастом в ответ на раздражение он становится менее обобщенным, со 2-й недели жизни появляется сосредоточение на звуке, а на 3-м месяце возникает типичная ориентировочная реакция, проявляющаяся в повороте головы в сторону раздражителя. Начальные стадии этой реакции связаны с ранним формированием рецепторов внутреннего уха, проводящих путей и четверохолмий, ее совершенствование — с развитием коленчатых тел и коркового отдела слухового анализатора.
К моменту рождения у плода хорошо развиты структуры, лежащие в основе рефлексов, возникающих в ответ на зрительные раздражения. Первоначальной формой ответных реакций являются защитные рефлексы.
Так, например, у новорожденных детей прикосновение к ресницам, конъюнктиве, роговице или дуновение вызывает смыкание век. Зона этого рефлекса у новорожденного шире — у него закрываются глаза и при прикосновении к кончику носа и лбу. При освещении спящего ребенка веки его смыкаются сильнее. Рефлекторное мигание (ответ на быстрое приближение предмета к глазам) появляется к 1,6—2 месяцам жизни.
У новорожденного хорошо развит зрачковый рефлекс. Этот рефлекс имеется даже у недоношенных детей. Расширение зрачков на звуковые и кожные раздражители появляется позже — с 10-й недели жизни ребенка.
В течение первого полугодия у большинства детей проявляется тонический рефлекс с глаз на мышцы шеи. Он проявляется в том, что в вертикальном положении тела ребенка (не поддерживая голову) при освещении глаз голова быстрым движением откидывается назад, тело при этом впадает в опистонус, то есть состояние, при котором тело выгибается назад вследствие повышения тонуса мышц-разгибателей. Реакция сохраняется до тех пор, пока глаза освещены. Этот рефлекс особенно хорошо выражен у новорожденных детей.
Лабиринтный, или установочный рефлекс, вследствие которого правильное положение в пространстве занимает сначала голова, а затем все тело, у новорожденного отсутствует. Этот рефлекс связан с формированием вестибулярного аппарата и красных ядер. Он хорошо выражен с 2—3месяцев жизни ребенка.
Лабиринтные рефлексы, возникающие при вращении (отклонении головы и глазных яблок в сторону, противоположную вращению), по данным большинства исследователей, имеют место сразу после рождения, они хорошо выражены с 7-го дня жизни ребенка. С первых дней жизни наблюдается и лифтная реакция, которая у ребенка выражается в поднимании рук вверх при быстром опускании тела (движение «падения»).
Рефлексы положения тела в пространстве зависят от правильного распределения тонуса мышц и суставов. Статические, установочные и выпрямительные рефлексы формируются после рождения. Их формирование связано с дальнейшим развитием головного мозга и коры больших полушарий. При этом происходит смена простейших рефлекторных актов на более сложные.
Так, например, врожденные предварительные локомоторные акты исчезают в 4—5 месяцев жизни ребенка. Первым исчезает рефлекс с глаз на шею (в 3 месяца), затем вестибулярная реакция на конечности (в 4—5 месяцев). Сокращение приводящих мышц противоположной ноги, сопровождающее коленный рефлекс, угасает к 7-ми месяцам, перекрестный сгибательный рефлекс ног — в 7—12месяцев, а ручной и ножной хватательный рефлекс переходит в произвольное хватание к концу первого года жизни. К этому времени почти полностью исчезает рефлекс Бабинского.
В течение первого года жизни ребенок учится переворачиваться на живот, ползать на животе и на четвереньках, сидеть, вставать и к концу года ходить.
Ретикулярная формация ствола мозга и ее влияние на активность различных отделов мозга.Ретикулярная формация (РФ) представлена сетью нейронов с многочисленными разветвлениями в разных направлениях. Нейроны расположены либо диффузно, либо образуют ядра.
Большинство нейронов РФ имеют длинные дендриты и короткий аксон. Имеются гигантские нейроны с длинным аксоном, которые образуют Т-образное ветвление: одна из ветвей аксона имеет нисходящее, а вторая восходящее направление. Так, например, в нисходящем направлении — ретикулоспинальный и руброспинальный пути. Аксоны нейронов РФ образуют большое число коллатералей и синапсов, которые оканчиваются на нейронах различных отделов мозга. Ретикулярная формация располагается в толще серого вещества продолговатого, среднего, промежуточного мозга (Атл., рис. 26, с. 135) и изначально связана с РФ спинного мозга. В связи с этим ее рассматривают как единую систему.
Ретикулярная формация имеет прямые и обратные связи с корой переднего мозга, базальными ганглиями, промежуточным мозгом, мозжечком, средним, продолговатым и спинным мозгом. Согласно современным представлениям, переход коры к активному состоянию связан с колебаниями количества восходящих сигналов от ретикулярной формации ствола мозга. Количество этих сигналов зависит от поступления в ретикулярную формацию сенсорных импульсов по коллатералям специфических афферентных восходящих путей. Практически к ретикулярной формации приходит информация от всех органов чувств по коллатералям от спинно-ретикулярного тракта, проприоспинальных путей, афферентных черепно-мозговых нервов, от таламуса и гипоталамуса, от моторных и сенсорных областей коры (рис. 9).
Большинство нейронов ретикулярной формации являются полисенсорными, то есть отвечают на раздражение различных модальностей (световых, звуковых, тактильных и т. д.). Ее нейроны имеют большие рецептивные поля, большой скрытый период и слабую воспроизводимость реакций. Эти свойства противоположны свойствам специфических ядер, и поэтому ретикулярные нейроны относят к неспецифическим.
| Ретикулярная формация |
| Моторная кора |
| Базальные ганглии |
| Лимбическая система |
| Сенсорные сигналы |
| Сенсорная кора |
| Таламус |
| Гипоталамус |
| Спинной мозг |
Рис. 10. Афферентные и эфферентные связи ретикулярной формации стволовой части мозга (по: Ноздрачев и др., 2004)
Однако исследования с раздражением РФ ствола мозга показали, что она может избирательно оказывать активирующее или тормозящее влияние на разные формы поведения, на сенсорные, моторные, висцеральные системы мозга.
Активность нейронов РФ различна и в принципе сходна с активностью нейронов других структур мозга, но среди нейронов РФ имеются такие, которые обладают устойчивой ритмической активностью, не зависящей от приходящих сигналов. В то же время в РФ среднего мозга и моста имеются нейроны, которые в покое «молчат», то есть не генерируют импульсы, но возбуждаются при стимуляции зрительных или слуховых рецепторов. Это так называемые специфические нейроны, обеспечивающие быструю реакцию на внезапные сигналы.
В ретикулярной формации продолговатого, среднего мозга и моста конвергируют сигналы различных модальностей. Сигналы от зрительной и слуховой сенсорных систем в основном приходят на нейроны среднего мозга.
РФ контролирует передачу сенсорной информации, идущей через ядра таламуса, за счет затормаживания нейронов неспецифических ядер таламуса, тем самым облегчается передача сенсорной информации в кору больших полушарий. В ретикулярной формации моста, продолговатого, среднего мозга имеются нейроны, которые реагируют на болевые раздражения, идущие от мышц или внутренних органов, что создает общее диффузное дискомфортное, не всегда четко локализуемое, болевое ощущение («тупая боль»).
Ретикулярная формация ствола мозга имеет прямое отношение к регуляции мышечного тонуса, поскольку на РФ ствола мозга поступают сигналы от зрительного и вестибулярного анализаторов и мозжечка. От РФ к мотонейронам спинного мозга и ядрам черепных нервов поступают сигналы, организующие положение головы, туловища и т. д. Ретикулярная формация ствола мозга участвует в передаче информации от коры больших полушарий, спинного мозга к мозжечку и, наоборот, от мозжечка к этим же системам. Функция данных связей заключается в подготовке и реализации моторики, связанной с привыканием, ориентировочными реакциями, болевыми реакциями, организацией ходьбы, движениями глаз. Ретикулярная формация принимает участие в регуляции функционирования дыхательного и сердечно-сосудистых центров. Так, например, повреждение дыхательного центра, расположенного в РФ продолговатого мозга, приводит к остановке дыхания.
Другим жизненно важным центром РФ является сосудодвигательный центр, который регулирует изменения просвета сосудов вен и артерий, артериальное давление. В регуляции вегетативных функций большое значение имеют так называемые стартовые нейроны РФ. Они дают начало циркуляции возбуждения внутри группы нейронов, обеспечивая тонус регулируемых вегетативных систем. Влияния ретикулярной формации на все отделы мозга можно разделить на нисходящие и восходящие. В свою очередь каждое из этих влияний имеет тормозное и возбуждающее действие.
Нисходящие влияния РФ ствола мозга на регуляторную деятельность спинного мозга были установлены еще И. М. Сеченовым (1862). Им было показано, что при раздражении среднего мозга кристалликами соли у лягушки рефлексы отдергивания лапки возникают медленно, требуют более сильного раздражения или не появляются вообще, то есть тормозятся.
Г. Мэгун (1945—1950), нанося локальные раздражения на РФ продолговатого мозга, нашел, что при раздражении одних точек тормозятся, становятся вялыми рефлексы сгибания передней лапы, коленный, роговичный. При раздражении РФ в других точках продолговатого мозга эти же рефлексы вызывались легче, были сильнее, то есть их реализация облегчалась. По мнению Мэгуна, тормозные влияния на рефлексы спинного мозга может оказывать только РФ продолговатого мозга, а облегчающие влияния регулируются всей РФ ствола и спинного мозга.
Восходящие влияния РФ на кору больших полушарий повышают ее тонус, регулируют возбудимость ее нейронов, не изменяя специфику ответов на адекватные раздражения. РФ влияет на функциональное состояние всех сенсорных областей мозга, следовательно, она имеет значение в интеграции сенсорной информации от разных анализаторов.
Ретикулярная формация имеет прямое отношение к регуляции цикла бодрствование — сон. Стимуляция одних структур РФприводит к развитию сна, стимуляция других вызывает пробуждение. Г. Мэгун и Дж. Моруцци выдвинули концепцию, согласно которой все виды сигналов, идущих от периферических рецепторов, достигают по коллатералям РФ продолговатого мозга и моста, где переключаются на нейроны, дающие восходящие пути в таламус и затем в кору больших полушарий.
Возбуждение РФ продолговатого мозга или моста вызывает синхронизацию активности коры больших полушарий, появление медленных ритмов в электроэнцефалограмме, сонное торможение. Такое же состояние головного мозга (спящий мозг) наблюдается при повреждении восходящих путей ретикулярной формации.
Возбуждение РФ среднего мозга вызывает противоположный эффект пробуждения; десинхронизацию электрической активности коры, появление быстрых низкоамплитудных (b-ритма) в электроэнцефалограмме. Следовательно, важнейшей функцией восходящей РФ является регуляция цикла сон — бодрствование.
Реакция активации коры головного мозга наблюдается при раздражении РФ продолговатого, среднего, промежуточного мозга. В то же время раздражение некоторых ядер таламуса приводит к возникновению ограниченных локальных участков возбуждения, а не к общему ее возбуждению, как это бывает при раздражении других отделов РФ.
Ретикулярная формация ствола мозга может оказывать не только возбуждающее, но и тормозное влияние на активность коры мозга. Тормозное влияние ретикулярной формации на передний мозг изучено значительно хуже. В. Гесс (1929), Дж. Моруцци (1941) установили, что раздражение некоторых точек ретикулярной формации ствола мозга может перевести животное из бодрствующего состояния в сонное. При этом возникает реакция синхронизации ритмов электроэнцефалограммы.
| <== предыдущая лекция | | | следующая лекция ==> |
| Продолговатый мозг, строение, функции и развитие | | | Строение, функции и развитие промежуточного мозга |
Источник: helpiks.org
Через средний мозг, являющийся продолжением ствола мозга, проходят восходящие пути от спинного и продолговатого мозга к таламусу, коре больших полушарий и мозжечку.
В состав среднего мозга входят четверохолмия, черная субстанция и красные ядра. Срединную его часть занимает ретикулярная формация (см. § 6 этой главы), нейроны которой оказывают мощное активирующее влияние на всю кору больших полушарий, а также на спинной мозг.
Передние бугры четверохолмия представляют собой первичные зрительные центры, а задние бугры—первичные слуховые центры. Ими осуществляют также ряд реакций, являющихся компонентами ориентировочного рефлекса при появлении неожиданных раздражителей. В ответ на внезапное раздражение происходит поворот головы и глаз в сторону раздражителя, а у животных—настораживания ушей. Этот рефлекс (по И. П. Павлову, рефлекс «Что такое?») необходим для подготовки организма к своевременной реакции на любое новое воздействие. Он сопровождается усилением тонуса мыщц-сгибателей (подготовка к двигательной реакции) и изменениями вегетативных функций (дыхание, сердцебиения).
Средний мозг играет важную роль в регуляции движений глаз. Управление глазодвигательным аппаратом осуществляют расположенные в среднем мозгу ядра блокового (IV) нерва, иннервирующего верхнюю косую мышцу глаза, и глазодвигательного (III) нерва иннервирующего верхнюю, нижнюю и внутреннюю прямые мышцы нижнюю косую мышцу и мышцу, поднимающую веко, а также расположенное в заднем мозгу ядро отводящего (VI) нерва, иннервирующего наружную прямую мышцу глаза. С участием этих ядер осуществляются поворот глаза в любом направлении, аккомодация глаза, фиксация взгляда на близких предметах путем сведения зрительных осей, зрачковый рефлекс (расширение зрачков в темноте и сужение их на свету).
У человека при ориентации во внешней среде ведущим является зрительный анализатор, поэтому особое развитие получили передние бугры четверохолмия (зрительные подкорковые центры). У животных с преобладанием слуховой ориентации (собака, летучая мышь), наоборот, в большей степени развиты задние бугры (слуховые подкорковые центры).
Черная субстанция среднего мозга имеет отношение к рефлексам жевания и глотания, участвует в регуляции тонуса мышц (особенно при выполнении мелких движений пальцами рук).
В среднем мозгу важные функции осуществляет красное ядро. О возрастании роли этого ядра в процессе эволюции свидетельствует резкое увеличение его размеров по отношению к остальному объему среднего мозга. Красное ядро тесно связано с корой больших полушарий, ретикулярной формацией ствола, мозжечком и спинным мозгом.
От красного ядра начинается руброспинальный путь к мотонейронам спинного мозга. С его помощью осуществляется регуляция тонуса скелетных мышц, происходит усиление тонуса мышц-сгибателей. Это имеет большое значение как при поддержании позы в состоянии покоя, так и при осуществлении движений. Импульсы, приходящие в средний мозг от рецепторов сетчатки глаза и от проприорецепторов глазодвигательного аппарата, участвуют в осуществлении глазодвигательных реакций, необходимых для ориентации в пространстве, выполнении точностных движений.
Источник: scorcher.ru
В среднем мозге замыкается ряд рефлексов. Так, при участии нейронов бугров четверохолмия осуществляются ориентировочные зрительные и слуховые рефлексы. У животных они проявляются в повороте головы и тела по направлению к свету и звуку, настораживании ушей.
Ядра четверохолмия участвуют в осуществлении сторожевого рефлекса, существенным компонентом которого является усиление тонуса сгибателей.
Черная субстанция участвует в сложной координации движений. В ней сосредоточены содержащие дофамин нейроны, многие из которых посылают аксоны в передний мозг. Они принимают участие в регуляции эмоционального поведения. Другая часть дофами- нергических нейронов черной субстанции посылает аксоны к ядрам полосатого тела, где дофамин играет важную роль в контроле сложных двигательных актов. Повреждения черной субстанции, приводящие к дегенерации дофаминергических волокон, проецирующихся в полосатое тело, приводят к нарушению тонких движений пальцев рук, развитию мышечной ригидности и тремору (болезнь Паркинсона).
После перерезки ствола мозга ниже уровня красного ядра у животных наблюдается выраженное изменение тонуса мускулатуры туловища и конечностей — децеребрацион-
ная ригидность, т. е. резкое повышение тонуса мышц-разгибателей. Конечности при этом сильно вытянуты, голова запрокинута назад, хвост приподнят (рис. 89).
Развитие децеребрационной ригидности связано с выключением импульсации, поступающей в спинной мозг по кортико-спиналь- ному и руброспинальному трактам, которые активируют в первую очередь мотонейроны мышц-сгибателей. При этом начинает преобладать активность вестибулоспинальной системы, повышающей тонус преимущественно мотонейронов мышц-разгибателей.
| 5* |
МОЗЖЕЧОК
Мозжечок представляет собой образование, расположенное позади больших полушарий мозга над продолговатым мозгом и мостом мозга. В эволюционном плане он представляет собой очень древнюю структуру. Впервые мозжечок появляется у круглоротых и сохраняет принципиально сходные функции во всем филогенетическом ряду позвоночных (от рыб до человека), будучи связан в первую очередь с моторной координацией. У млекопитающих и человека физиологическое значение мозжечка особенно возрастает, а его нейронная организация и структура отдельных нервных элементов достигает наибольшей сложности.
Анатомически в мозжечке можно выделить среднюю часть — червь, расположенные по обе стороны от нею полушария и боковые флоккулонодулярные доли. Последние филогенетически представляют самую древнюю часть мозжечка — аршоцеребеллум. Полушария мозжечка в свою очередь делятся на переднюю и заднюю доли. Передние доли полушарий и задняя часть червя мозжечка составляют старый мозжечок — палеоцеребеллум. Наконец, филогенетически самая молодая часть мозжечка, связанная двусторонними связями с наиболее новыми образованиями мозга — новой корой,— неоцеребеллум, включает переднюю часть задних долей полушарий мозжечка.
В полушариях мозжечка выделяют верхнюю поверхность, образующую кору мозжечка и скопления нервных клеток — ядра мозжечка. Мозжечок связан с другими отделами ЦНС тремя парами ножек, образованных пучками нервных волокон.
Нейронная организация. Нейронная организация мозжечка отличается исключительной упорядоченностью. Кора мозжечка построена по единому принципу и состоит из 3 слоев. В поверхностном, или молекулярном, слое находятся дендритные разветвления грушевидных клеток (клетки Пуркинье), представляющих собой одни из наиболее сложно устроенных нейронов мозга. Грушевидные клетки имеют чрезвычайно разветвленное дендритное дерево. Дендритные отростки, в особенности в дистальных частях, обильно покрыты шипиками. Многочисленные разветвления дендритов резко увеличивают площадь поверхностной мембраны. Это создает условия для размещения огромного числа синапсов. Подсчитано, что один грушевидный нейрон имеет до 200 ООО синапсов. Кроме дендритов грушевидных клеток, в поверхностном слое располагаются так называемые параллельные волокна, представляющие собой аксоны многочисленных вставочных нейронов.
 Рис. 80. Децеребрацнонная ригидность. Рис. 80. Децеребрацнонная ригидность. |
В нижней части молекулярного слоя находятся тела корзинчатых клеток, аксоны которых образуют синапгические контакты с телами грушевидных клеток. В молекулярном слое также имеется некоторое число звездчатых клеток. Далее следует ганглиозный слой, в котором находятся тела грушевидных клеток. В следующем гранулярном слое коры мозжечка находятся тела вставочных нейронов (клеток-зерен, или гранулярных клеток). Аксоны гранулярных клеток поднимаются в молекулярный слой, где они Т-образно разветвляются. В гранулярном слое находятся также клетки Гольджи, аксоны которых направляются в молекулярный слой.
| Рис. 90. Синаптические сняли нейронов мозжечка |3kk.ic Дж, 1009[. |
Схематически показаны шпбуждаюшие синапсы, образуемые на грушевидных нейронах (клетки ПуркинмЧ лазающими нолокнамн (JIB I. аксонами клеток-зерен (КЗ). которые в слою очередь активируются мшистыми волокнами (МВ). и тормозные синапсы обр л.юна мы аксон;] ми эпеддчатых [ЗвЮ н корзиичатык клеток; Т — торможение; В — возбуждение
В кору мозжечка поступают только два типа афферентных волокон: лазающие и мшистые (или моховидные). По этим каналам в мозжечок доставляются все сенсорные влияния. Лазающие волокна, являющиеся аксонами нейронов нижних олив, образуют синапсы с основаниями дендритов грушевидных клеток. Каждое лазающее волокно контактирует обычно с одной грушевидной клеткой. Однако возбуждающее действие лазающего волокна столь эффективно, что грушевидная клетка отвечает на одиночный импульс в лазающем волокне ритмическим разрядом потенциалов действия.
В противоположность лазающим волокнам, моховидные волокна характеризуются значительной дивергенцией. Разветвление одного моховидного волокна образуют синапсы примерно на 20 вставочных нейронах, но не контактируют непосредственно с грушевидными клетками. Число вставочных нейронов примерно в 2300 раз превышает число клеток Пуркинье, аксоны их, разветвляясь в молекулярном слое, образуют систему параллельных волокон, оканчивающихся синапсами на более дистальных, покрытых много численными шипами дендритах грушевидных клеток. Указанные синапсы, как и синапсы, образуемые лазающими волокнами, являются возбуждающими.
Мшистые волокна, кроме того, образуют синапсы с корзинчатыми клетками. Аксоны корзинчатых клеток образуют густые сплетения типа корзинок вокруг тел грушевидных клеток, обеспечивающие значительную площадь синаптических контактов. Синапсы между аксонами корзинчатых клеток и телами грушевидных нейронов являются тормозными. Они обеспечивают эффективное торможение возбуждающих влияний, оказываемых на грушевидные клетки через аксодендритные синапсы, образуемые лазающими волокнами и аксонами вставочных нейронов.
Наконец, мшистые волокна образуют синаптические контакты также с клетками Гольджи и звездчатыми клетками. Как и корзинчатые клетки, клетки Гольджи и звездчатые клетки являются тормозными нейронами. Однако аксоны клеток Гольджи заканчиваются не на грушевидных клетках, а на многих тысячах вставочных нейронов.
Схема синаптической организации нейронов коры мозжечка показана на рис. 90.
| |
| |
Рис. 91. Синаптические эффекты, вызываемые в поясничном мотонейроне обезьяны раздражением промежуточного ндра мозжечка (а), красного ядра (б) и обеих структур имеете (а+б). Верхняя кривая — нанесение раздражении, нижняя кривая — ответы мотонейрона.
Если в кору мозжечка входит два типа афферентных волокон: лазающие и мшистые, то покидает ее всего лишь один тип эфферентных волокон, являющихся аксонами грушевидных нейронов (нейроны Пуркинье). Таким образом, грушевидные клетки образуют единственный выход всей сложно организованной нейронной системы, составляющей кору мозжечка. Другим отличительным свойством грушевидных клеток является то, что все они являются тормозными нейронами, т. е. образуют тормозящие синапсы со всеми клетками, с которыми они контактируют. Тормозные постсинаптические потенциалы при активации грушевидных клеток возникают в нейронах собственных ядер мозжечка и в нейронах вестибулярных ядер.
В мозжечок поступает информация из различных сенсорных систем. Афферентные сигналы достигают мозжечка по различным путям, которые можно подразделить на следующие группы: восходящие от спинного мозга (по спинно-мозжечковым трактам), от вестибулярных рецепторов, от нижней оливы и от ретикулярной формации заднего мозга.
Волокна дорсального и вентрального спинно-мозжечковых трактов доставляют в мозжечок информацию о состоянии мышечного аппарата.-
Спинно-ретикуломозжечковый тракт имеет переключение в латеральном ретикулярном ядре продолговатого мозга. По этому пути в мозжечок поступает информация от кожи и более глубоких тканей.
Важный афферентный вход кора мозжечка получает из нижней оливы, где происходит переключение импульсов, поступающих по нескольким путям, берущим начало как в спинном мозге, так и в структурах головного мозга. Значительную роль при этом играют сигналы, поступающие в его кору из больших полушарий по мшистым и лазающим волокнам.
Наконец, в коруфлоккулонодулярной доли приходят первичные и вторичные вестибулярные афферентные влияния.
Из красного ядра к мозжечку подходят коллатерали руброспинальных аксонов. Нейроны промежуточного ядра мозжечка посылают волокна к клеткам красного ядра. Синапсы, образуемые этими волокнами на руброспинальных нейронах, являются возбуждающими и характеризуются высокой эффективностью. Поэтому раздражение промежуточного ядра мозжечка вызывает в спинальных мотонейронах ответы, сходные с теми, которые возникают при стимуляции красного ядра (рис. 91).
Нейроны других мозжечковых ядер образуют возбуждающие синапсы на ретикулоспинальных нейронах продолговатого мозга и моста. Итак, вся информация, приходящая в мозжечок, передается грушевидным клеткам или клеткам Пуркинье, а те в свою очередь оказывают тормозящее влияние на ядра мозжечка (а через них тормозят активность ретикуло- и руброспинальных нейронов) и на нейроны преддверного латерального ядра (ядро Дейтерса), дающие начало вестибулоспинальному тракту. Таким образом, мозжечок может эффективно контролировать значительную часть команд, поступающих в спинной мозг по основным нисходящим трактам. Действительно, после удаления мозжечка ритмика вестибуло-, ретикуло- и руброспинальных нейронов, которая в норме изменяется в соответствии с выполнением определенной части двигательного акта, перестает зависеть от двигательных циклов.
Функции мозжечка
Несмотря на исключительную упорядоченность нейронной организации мозжечка, благодаря которой его сравнивают со своего рода нейронной машиной, его функциональная роль раскрыта еще не полностью.
Клинические проявления, развивающиеся при поражении мозжечка, а также эффекты, наблюдаемые при его раздражении или экстирпации, свидетельствуют о важной роли мозжечка в осуществлении статических, статокинетических рефлексов и других процессов управления двигательной активностью, автоматически регулирующих работу двигательного аппарата.
Экспериментальные и клинические наблюдения показывают, что при поражениях мозжечка развиваются разнообразные нарушения двигательной активности и мышечного тонуса, а также вегетативные расстройства. Основные проявления расстройств двигательной системы включают нарушения равновесия и мышечного тонуса: тремор, атаксию и асинергию движений.
Полное удаление мозжечка или его передней доли у животных приводит к повышению тонуса мышц-разгибателей, в то время как раздражение передней доли — к снижению этого тонуса (торможение децеребрационной ригидности).
Через несколько суток после удаления мозжечка тонус разгибателей ослабляется, сменяясь гипотонией, лежащей в основе двигательных нарушений. В этот период животные с удаленным мозжечком не могут не только ходить, но и стоять и в то же время способны хорошо плавать.
Многие клинические проявления, свидетельствующие о мозжечковой недостаточности, связаны с мышечной атонией и неспособностью поддерживать позу. После исследования коленного рефлекса или смещения пассивно висящей конечности нога не возвращается в исходное положение, а раскачивается подобно маятнику.
Одно из наиболее характерных проявлений мозжечковой недостаточности — возникновение тремора. Тремор покоя характеризуется небольшой амплитудой, колебания протекают синхронно в разных сегментах тела. Для мозжечковых повреждений характерна также атаксия: нарушение величины, скорости и направления движений, что приводит к утрате плавности и стабильности двигательных реакций. Целенаправленные движения, например попытка взять предмет, выполняются порывисто, рывками, промахами мимо цели. Атаксический тремор наблюдается при выполнении произвольных движений, будучи наиболее выражен в начале и в конце движения, а также при перемене его направления.
Асинергия проявляется также нарушением взаимодействия между двигательными центрами различных мышц. Так, у больных при выполнении движений не происходит одновременного сокращения мышц, компенсирующих смещение центра тяжести. Делая шаг, больной выносит ногу, не сгибая, впереди туловища, что приводит к падению. В результате асинергии сложные движения как бы распадаются на ряд выполняемых последовательно более простых движений.
Разновидностью асинергии можно считать характерный для поражения мозжечка симптом — адиадохокинез — нарушение правильного чередования противоположных движений, например сгибания и разгибания пальцев.
Итак, расстройство равновесия и тонуса скелетной мускулатуры приводит к характерным нарушениям в осуществлении произвольных движений. Особенно сильно затрудняется выполнение задач, связанных с необходимостью точно коснуться какого-либо предмета. Нарушение двигательной координации — астазия — характеризуется появлением качательньгх и дрожательных движений. Нарушение локомоции — атаксия — проявляется расстройством походки, которая становится неровной, зигзагообразной.
Наступает неадекватное перераспределение мышечного тонуса — дистония. Локальные повреждения небольших участков коры передней доли мозжечка позволили выявить локализацию в ней представительства различных участков скелетной мускулатуры.
Нарушение двигательной координации при поражениях мозжечка объясняется тесными связями его с основными структурами ствола мозга (дающими начало трактам, передающим импульсы в спинной мозг), а также с таламусом и сенсомоторной областью коры больших полушарий.
Одна из главных проекций мозжечка направлена к нейронам ретикулярной формации ствола, особенно к тем из ретикулоспинальных клеток, аксоны которых отличаются наиболее высокой скоростью проведения возбуждения. Учитывая, что последние устанавливают контакты с а- и у-мотонейронами, можно заключить, что через ретикулоспиналь- ный путь обеспечивается срочная передача мозжечковых команд непосредственно к этим ключевым элементам спинального управления движениями.
Ретикулоспинальные нейроны находятся под мозжечковым контролем, после устранения которого участие их в центральной регуляции движений резко нарушается.
Связи между мозжечкам и преддверным латеральным ядром (ядро Дейтерса), дающим начало вестибулоспинальному тракту, настолько тесны и характерны, что это вестибулярное ядро функционально можно рассматривать как ядро мозжечка, вынесенное в продолговатый мозг. Значительная часть мозжечкового возбуждающего и тормозящего контроля спинальных центров обеспечивается с помощью вестибулоспинальных нейронов.
Команды из промежуточной коры и промежуточного ядра мозжечка передаются к спинному мозгу через нейроны красного ядра.
Таким образом, нейронная организация мозжечка обеспечивает поступление в его кору разнообразной афферентной информации, в том числе от различных компонентов двигательного аппарата, сложную обработку этой информации в нейронах и синапсах мозжечка и эффективную передачу корригирующих влияний к нейронам стволовых и спинальных центров моторного контроля.
Как было показано JI. А. Орбели, мозжечок играет также важную роль в регуляции вегетативных функций за счет многочисленных синаптических связей с ретикулярной формацией ствола мозга.
ПРОМЕЖУТОЧНЫЙ МОЗГ
Промежуточный мозг образует стенки III желудочка. В процессе эмбриогенеза он формируется вместе с большими полушариями из переднего мозгового пузыря.
Главными структурами промежуточного мозга являются таламус, или зрительный бугор, и гипоталамус, или подбугроеая область. Ядра таламуса расположены главным образом в области боковой стенки III желудочка; ядра гипоталамуса образуют его нижнюю и нижнебоковую стенки. Верхняя часть III желудочка образована сводом и эпифизом (эпиталаму с).
В глубине мозговой ткани промежуточного мозга расположены ядра наружных и внутренних коленчатых тел. Наружная граница промежуточного мозга проходит лате- ральнее коленчатых тел и образована белым веществом внутренней капсулы, отделяющей промежуточный мозг от подкорковых ядер конечного мозга.
ТАЛАМУС
Нейронная организация. Нервные клетки таламуса, группируясь, образуют большое количество ядер: всего различают до 40 таких образований. Топографически все они могут быть подразделены на несколько основных групп: передние, интраламинарньге, срединные и задние. В каждой из этих основных групп различают более мелкие ядра, отличающиеся друг от друга как нейронной организацией, так и особенностями афферентных и эфферентных проекций. С функциональной точки зрения принято различать неспецифические и специфические ядра таламуса. Нейроны неспецифических ядер посылают аксоны диффузно ко всей новой коре, в то время как нейроны специфических ядер образуют связи только с клетками определенных корковых полей (рис. 92). Неспецифические ядра являются по своему происхождению более древними и включают срединные и интраламинарные ядра, а также медиальную часть переднего вентрального ядра. Нейроны неспецифических ядер сначала передают сигналы в подкорковые структуры, от которых импульсация поступает параллельно в разные отделы коры. Не специфические ядра являются продолжением ретикулярной формации среднего мозга, представляя собой ретикулярную формацию таламуса.
На нейронах специфических ядер заканчиваются волокна различных восходящих трактов. Аксоны этих нейронов образуют прямые моносинаптические связи с нейронами сенсорной и ассоциативной коры. К клеткам ядер латеральной группы таламуса, включающих заднее вентральное ядро, поступают импульсы от кожных рецепторов, двигательного аппарата, а также мозжечково-таламического пути.
Другая часть специфических ядер таламуса входит в состав задней группы и образует медиальное и латеральное коленчатые тела. На нейронах латерального коленчатого тела заканчиваются афферентные пути зрительной системы. Нейроны медиального коленчатого тела воспринимают сигналы от нейронов слуховых ядер продолговатого мозга и задних бугров четверохолмия.
Нейроны специфического комплекса ядер посылают по направлению к коре аксоны, почти не имеющие коллатералей. В отличие от него нейроны неспецифической системы посылают аксоны, дающие множество коллатералей. Вместе с тем и волокна, приходящие из коры к нейронам специфических ядер, характеризуются топографической локали- зованностью своих окончаний в противоположность широко разветвленной системе диф- фузно оканчивающихся волокон в неспецифических ядрах.
Функции таламуса
Все сенсорные сигналы, за исключением возникающих в обонятельном тракте, достигают коры больших полушарий только через таламокортикальные проекции. Таламус представляет собой своего рода ворота, через которые в кору поступает и достигает сознания основная информация об окружающем нас мире и о состоянии нашего тела.
Тот факт, что афферентные сигналы на пути к коре мозга переключаются на нейронах таламуса, имеет важное значение. Тормозные влияния, приходящие в таламус из коры, других образований и соседних таламических ядер, позволяют обеспечить лучшую передачу в кору мозга наиболее важной информации. Торможение подавляет слабые возбуждающие влияния, благодаря чему выделяется наиболее важная информация, приходящая в таламус от различных рецепторов.
Через неспецифические ядра таламуса в кору мозга поступают восходящие активирующие влияния от ретикулярной формации мозгового ствола. Система неспецифических ядер таламуса осуществляет контроль ритмической активности коры больших полушарий и выполняет функции внутриталамической интегрирующей системы.
Электрическое раздражение неспецифических ядер таламуса вызывает в коре больших полушарий периодические колебания потенциалов, синхронные с ритмом активности таламических структур. Реакция в коре возникает с большим скрытым периодом и значительно усиливается при повторении. Таким образом, нейроны коры больших полушарий вовлекаются в процесс активности как бы постепенно. Такая реакция вовлечения коры мозга отличается от специфических ее ответов своей’ генерализованностью, охватом обширных областей коры. Активацию нейронов неспецифических ядер таламуса особенно эффективно вызывают болевые сигналы (таламус является высшим центром болевой чувствительности). Импульсы, идущие по путям болевой чувствительности, формируются при раздражении различных областей тела и внутренних органов. Скрытые периоды ответов в таламусе отличаются большой длительностью и вариабельностью.
Наблюдения, проведенные на людях во время хирургических операций, показывают, что повреждения неспецифических ядер таламуса приводят к нарушениям сознания. Это
| Рис. 93. Схематическое изображение основных ядер гипоталамуса. I — передняя комиссура; 2 — прсоптиче* с кое ядро; 3 — супраоптическое ядро; 4 — ножка гипофиза; 5 — йентромедиальное гипоталамическое ядро; 6 — до рсо медиальное гипоталамическое ядро; 7 медиальное ядро мамиллнрнош тела; 8 — ма- миллярно-галамический лучок: 9- заднее гипоталамическое ядро; 10 — паравентри- кулярное ядро. |
Источник: megaobuchalka.ru