Рецепторная функция мембраны
Клеточная мембрана также называется плазматической (или цитоплазматической) мембраной и плазмалеммой. Данная структура не только отделяет внутреннее содержимое клетки от внешней среды, но также входит с состав большинства клеточных органелл и ядра, в свою очередь отделяя их от гиалоплазмы (цитозоля) — вязко-жидкой части цитоплазмы. Договоримся называть цитоплазматической мембраной ту, которая отделяет содержимое клетки от внешней среды. Остальными терминами обозначать все мембраны.
Строение клеточной мембраны
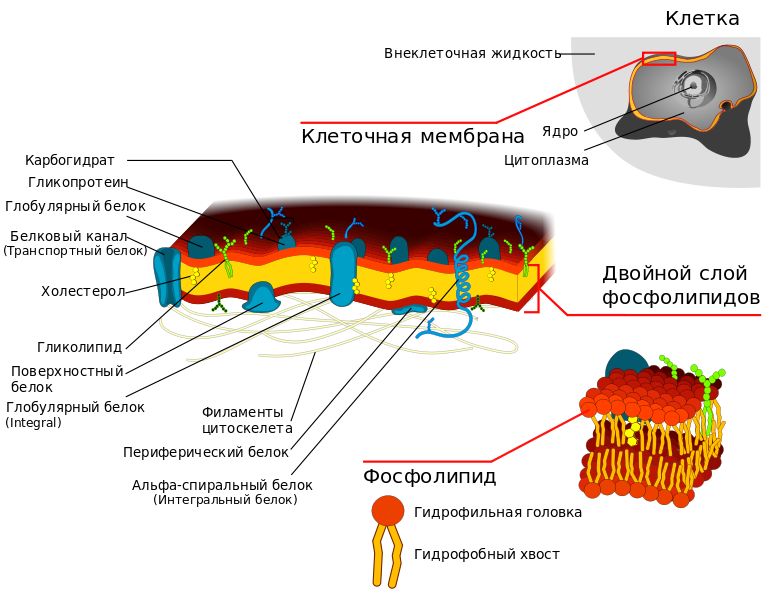
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.

В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков. Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.
Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Большинство погруженных в клеточную мембрану белков выполняют ферментативную функцию (являются ферментами). Часто (особенно в мембранах органоидов клетки) ферменты располагаются в определенной последовательности так, что продукты реакции, катализируемые одним ферментом, переходят ко второму, затем третьему и т. д. Образуется конвейер, который стабилизируют поверхностные белки, т. к. не дают ферментам плавать вдоль липидного бислоя.
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции. Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).
При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO2, O2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану. Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример — натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами). Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз — это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа. При этом могут открываться каналы, и в клетку могут начать поступать определенные вещества или выводиться из нее.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) — синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
Источник: biology.su
Транспортная функция
Плазматическая мембрана обладает свойствами полупроницаемого барьера, который пропускает и не пропускает только определенные вещества и молекулы. Выделяют несколько разновидностей транспорта через мембрану: пассивный транспорт, активный транспорт, ионный транспорт, транспорт в мембранной упаковке (эндо и экзоцитоз).
Пассивный транспорт – это транспорт через мембрану веществ из зоны высокой концентрации в зону низкой концентрации. Он осуществляется в двух формах: в форме простой диффузии и в форме «облегченной» диффузии. Оба этих процесса не нуждаются в энергии, идут относительно медленно и останавливаются, когда концентрация веществ по обе стороны мембраны уравняется. Скорость диффузии и сама возможность транспорта веществ через мембрану зависит (помимо концентрации) от ряда факторов: температуры, размера молекул, способности растворяться в липидах. Жирорастворимые вещества проходят через липидные слои легко, водорастворимые – с трудом. В мембране существуют специальные липидные и белковые «поры», через которые и проходит диффузия. Простая диффузия относительно медленный процесс и природа приспособила для ускорения транспорта специальные мембранные белки – переносчики. Они соединяются с транспортируемым веществом и переносят его с одной стороны мембраны на другую. Для каждой группы веществ в мембране должны быть свои переносчики. Такой процесс и называется «облегченная диффузия», протекая в десятки раз быстрее простой диффузии.
Ионный транспорт – это разновидность пассивного транспорта для заряженных ионов, но имеет свои особенности. Белки в цитоплазме клетки несут на своей поверхности, как правило, отрицательный заряд, создавая определенный электрический фон в клетке. Если в клетку направляются потоки положительных (катионы) или отрицательных (анионы) ионов, то положительных ионов войдет больше, поскольку часть катионов будет связываться белками цитоплазмы, нейтрализоваться и создавать дополнительную разницу концентраций разнозаряженных ионов внутри и снаружи клетки. Для транспорта ионов в мембране есть либо специальные ионные поры, либо переносчики.
Активный транспорт – наиболее важная форма транспорта для клетки. Диффузия, если она продолжается достаточно долго, приводит к тому, что по обе стороны мембраны устанавливается равновесие, что недопустимо для клетки – это для нее смерть. Состав цитоплазмы и межклеточной жидкости сильно различаются. Поэтому в клетке существует система «активного транспорта». В этом случае транспорт молекул осуществляется против градиента концентрации (из зоны низкой концентрации в зону высокой). Для этого существуют специальные белковые мембранные комплексы (ионные и молекулярные каналы), работающие с затратой энергии. До 40% всей энергии, вырабатываемой клеткой, идет на транспортные расходы!
Транспорт в мембранной упаковке (эндо – и экзоцитоз). Во многих случаях клетка должна транспортировать через мембрану не отдельные молекулы или ионы, а целые молекулярные комплексы и даже частицы. Они через мембранные поры не проходят. В связи с этим, в клетке существует специальная разновидность транспорта – транспорт в мембранной упаковке. В одних случаях происходит выпячивание наружной плазматической мембраны, охватывающей инородную частицу (например: частицу пищи или бактерию), замыкание ее в мембранное кольцо и погружение вглубь цитоплазмы клетки. Такой процесс был открыт знаменитым Российским ученым, лауреатом Нобелевской премии И.И.Мечниковым и назван фагоцитозом. Это явление было обнаружено при изучении защитных свойств клеток крови, которые захватывали бактерии и нейтрализовали их. Более мелкие частицы и капельки жидкости клетка захватывает путем образования впячиваний мембраны – этот процесс получил наименование пиноцитоз (греч. pinos- жидкость).
На самом деле фагоцитоз и пиноцитоз – это разновидности общего явления, характерного для большинства клеток – эндоцитоза. Процесс обратный эндоцитозу – выведение из клеток каких либо веществ и продуктов получил название экзоцитоза. На базе мембранного транспорта построен весь процесс выделения и поглощения секретов и гормонов клетками. Важно подчеркнуть, что эндо- и экзоцитоз являются энергозатратными процессами и, таким образом, относятся к разновидностям активного транспорта.
Рецепторная функция мембраны
Принципы работы мембранных рецепторов.
Клетка постоянно получает сигналы из внешней среды о наличии там разнообразных сигнальных молекул и должна адекватно отвечать на эти сигналы, передавая информацию от них внутрь клетки. Для этого в плазматической мембране встроены специальные рецепторные комплексы. Как правило, это сложные образования из нескольких белковых молекул (в состав рецепторов могут входить также мембранные липиды и углеводы).
Все разнообразные рецепторы клеточных мембран имеют ряд общих особенностей:
1. Рецепторы специфичны – т.е. связываются только с определенными веществами. Специфичность рецептора определяется структурой «активного центра» в его молекуле и возможностью других молекул связываться с этим активным центром.
2. Процесс рецепции и передачи сигнала на мембрану или вглубь клетки проходит со значительной затратой энергии.
3. По принципу работы все рецепторы можно разделить на три группы: рецепторы прямого действия; рецепторы непрямого действия и каталитические рецепторы. В первом случае молекула связывается с рецепторной частью комплекса и передает сигнал непосредственно на ионный канал. Во втором варианте рецепторная часть комплекса передает сигнал на ионный канал в мембране или вглубь клетки через систему вспомогательных белков, называемых «вторичными посредниками». При третьем варианте рецепторная часть комплекса после взаимодействия с сигнальной молекулой активируется и выполняет функции фермента, влияя, таким образом, на работу клетки.
Изолирующая функция мембраны. Межклеточные контакты
В многоклеточном организме клетки находятся в постоянном контакте друг с другом, который иногда приобретает черты тесного взаимодействия. Особенно это проявляется в тканях, которые граничат с наружной средой (покровы, кишечник, дыхательные пути) или где возникает необходимость тесного контакта клеток для согласованной работы (гладкие и сердечные мышцы, нервные клетки и волокна).
Встречаются четыре разновидности межклеточных контактов: изолирующие контакты (tigth junction), плотные контакты (gap junction), десмосомы (desmosome) и адгезионные контакты (adhesion junction).
Изолирующие контакты оправдывают свое название – белковые комплексы в мембранах соседних клеток соединяются таким образом, что через этот контакт не проходят никакие молекулы и ионы. Плотные контакты пропускают мелкие молекулы, ионы и через них может происходить обмен между клетками. В частности, плотные контакты приспособлены нервными и мышечными клетками для передачи электрических сигналов. Десмосомы предназначены для скрепления группы клеток друг с другом или с каким-либо другим субстратом.
Они могут быстро исчезать и снова образовываться. Адгезионные контакты также предназначены для прикрепления клеток друг к другу. Они более лабильны и могут быстро образовываться и исчезать по мере необходимости.
Источник: letopisi.ru
<<Назад / Далее>>
В основе рецепции лежит свойство биоструктур клеточного уровня переходить от состояния покоя в возбужденное состояние под действием внешних раздражителей и трансформировать энергию раздражения в информационный импульс (нервный, гуморальный, электромагнитный, акустический). Главенствующую роль в формировании возбуждения клетки играют структура и функциональное состояние ее цитоплазматической мембраны. Как известно, цитоплазматические мембраны представляют собой двойной слой фосфолипидных молекул с встроенными или примыкающими к нему белками и комплексами небелкового происхождения, включающими стероидные гормоны, простагландины, ионы металлов и т. п. (рис. 1).
Не останавливаясь на описанных в многочисленной специальной литературе биофизических и физико-химических свойствах этих мембран, выделим из них лишь наиболее важные с точки зрения понимания механизмов клеточной рецепции.
Для нормально функционирующей цитоплазматической мембраны необходима строго определенная среда по ионному составу и температуре. Известно, что мембрана обладает избирательностью по проницаемости для различных ионов и периодически ее меняет при возбуждении. С изменением температуры в биомембранах обнаружены генерализованные скачкообразные фазовые переходы, резко изменяющие их свойства, такие как вязкость, активность ферментативных реакций и др. [12]. Природа этих явлений остается недостаточно изученной.
Молекулы фосфолипидов и белков, составляющих структуру биомембраны, при физиологических температурах обладают высокой подвижностью в плоскости мембраны. Это явление называют латеральной диффузией и природу его относят к хаотическому тепловому перемещению. Следует отметить, что относительно размеров клетки скорость так называемой хаотической диффузии весьма высока. По данным, приведенным Ю. А. Владимировым с соавт. [13, с. 120-121], в отдельных видах липосом молекула фосфолипида в среднем перемещается за 1 секунду на 5 мкм, а для антигенов на поверхности лимфоцита скорость перемещения составляет примерно 0,2 мкмЧс-1. Биомолекулы в мембранах также совершают достаточно интенсивные вращательные движения: поворот на 1 радиан происходит для различных молекул за период 10-9ё10-4 секунды.
Известно, что встроенные в фосфолипидный бислой сложные по структуре молекулы белка обладают конформационной динамикой под действием факторов как внутриклеточной, так и внеклеточной (внешней) сред. Так, по А. Б. Рубину [14, с. 192], «флуктуации молекулярного объема у молекул с молекулярной массой до 14000 могут составить при 300 К 0,03-0,04 нм3, а флуктуации поверхности — до 0,01 нм2, то есть 0,1 % всей площади поверхности молекулы. Длина водородной связи — основного фактора, стабилизирующего вторичную структуру, может меняться при тепловых флуктуациях(300 К) на 0,01 нм, а углы — на 20-25о«. Конформационные состояния создаются балансом сил ковалентных, ион-дипольных, водородных и других взаимодействий, каждое из которых обладает определенной энергией. Наиболее слабые гидрофобные и водородные связи обладают энергией взаимодействия 4ё25 кДжЧмоль-1, ковалентные связи, определяющие первичную структуру белка — 150-700 кДжЧмоль-1. При этом каждое конформационное состояние молекулы белка обладает специфичным только для данного его состояния набором физико-химических проявлений, в частности, энергией активации реакций обмена.
Характерным свойством любой живой клетки является наличие разности электрического потенциала между внешней и внутренней сторонами цитоплазматической мембраны, при этом внутри клетки потенциал отрицательный по отношению к наружной стороне. Обычно трансмембранная разность потенциалов (D) в состоянии покоя для нервных и мышечных клеток составляет 60ё90 мВ. Здесь следует обратить внимание на то, что если применить расчет для плоского конденсатора, то при толщине (d) мембраны 4ё9 нм напряженность (Е) электрического поля составит весьма внушительную величину. Например,
если принять D=70 мВ=70*10-3 В, а d=7 нм=7*10-9 м, то:
Прежде чем рассматривать механизмы рецепторной функции клеток, обратим внимание на соотношение размеров объектов взаимодействия в процессе рецепции: клетка — кванты света (фотоны), молекулы вещества, ионы. Поперечный размер клеток составляет от долей до одного-двух десятков микрометров (10-6 м). Для фотона примем размерность электрона -10-15 м, для молекул-10-9 м. Теперь, чтобы упростить наше ассоциативное представление увеличим перечисленные размерности, например, в 1010 раз. Размерность клетки станет 104 м, тогда участок длиной в 1 мкм превращается в 10 км, а площадь поверхности цитоплазматической мембраны в 1 мкм2 составит 108 м2 или 10 тысяч гектаров. Фотон будет иметь размерность 10-5 м или 0,01 мм, а молекула — 10 м. Таким образом, возникает наглядное соотношение: на поверхность 10 тыс. га могут воздействовать как частицы с размером много меньшим макового зерна, так и десятиметровые «глыбы». Но, что такое даже десяти метровое в диаметре тело по отношению к 10 тыс. га поверхности — это меньше одной миллионной ее части.
При таком соотношении размерностей фотоны или молекулы вещества могут вызвать возбуждение клеточных процессов за счет скорости движения, энергетического уровня, резонансных, электрохимических и других явлений взаимодействия со специализированными в сенсорном отношении структурами — рецептивными центрами клетки. Такими центрами (структурами) являются, прежде всего, белки и их ассоциации с лигандами, встроенные в фосфолипидный бислой и специфически реагирующие своей структурной организацией на тот или иной вид воздействия, иначе говоря, способные «узнавать» воздействующий сигнал, создавать возмущение мембраны в зоне своей локализации и тем самым выполнять функцию рецепторного аппарата клеток. Эти белки называют белками-рецепторами. Комплексы небелкового происхождения, обладающие определенной структурой, также могут исполнять роль рецептивных центров [15]. В состав рецептивных центров входят стероидные гормоны, простагландины и ионы металлов, связанные в специфические структуры посредством водородных, ионных, гидрофобных и других взаимодействий.
Источник: www.medinfo.ru
Рецепторная функция мембран обеспечивает взаимодействие клетки с микроокружением; участие клетки в реакциях ткани, органа; участие ядра, органелл в формировании реакции клетки на воздействии.
Информационные сигналы, которые воздействуют на цитоплазматическую мембрану и вызывают значимые изменения в деятельности клетки, можно сгруппировать в три группы:
1. Изменение потенциала мембраны.
2. Изменение напряжение билипидного слоя мембраны или цитоскелета клетки.
3. Сигнальные молекулы (лиганды).
Классификация мембранных рецепторов
По локализации делятся на цитоплазматические и ядерные.
По механизму развития событий рецепторы делятся на ионотропные и метаботропные.
Ионотропные рецепторы относят к быстроотвечающим рецепторам, ответ в течение миллисекунд.
Формируются интегральными белками, имеют несколько субъединиц. Содержат субъединицу, имеющую центр связывания для сигнальной молекулы.
Центры связывания для сигнальной молекулы у ионотропных рецепторов делятся на:
- потенциалзависимые сенсоры;
- механозависимые сенсоры;
- сенсоры для внеклеточных и внутриклеточных лигандов.
Метаботропные рецепторы — медленноотвечающие (секунды, минуты, часы).
Метаботропные рецепторы делятся на две большие группы:
- рецепторы, связанные с ионными каналами. Изменение проницаемости ионных каналов реализуется через вторые посредники;
- рецепторы, не связанные непосредственно с мембранными каналами.
Рецепторы, не связанные непосредственно с мембранными каналами делятся на:
1. Рецепторы, связанные с G-белком. К этой группе относится большая часть рецепторов.
2. Каталитические рецепторы:
— с собственной гуанилитциклазной активностью. К ним относятся рецепторы, обладающие способностью реализовывать сигнал через цГМФ опосредованный путь;
— с собственной тирозинкиназной активностью. К ним относятся рецепторы к инсулину, активация которых вызывает фосфорилирование различных групп внутриклеточных белков, которые, меняя свою биологическую активность, вызывают широкий спектр
реакций, присущих инсулину.
3. Рецепторы, освобождающие факторы транскрипции.
Находятся в мембранах цитоплазмы и эндоплазматического ретикулума. При активации от них протеолитическими ферментами цитозоля отщепляется пептидный фрагмент, который, попадая в ядро клетки, запускает транскрипцию соответствующего гена.
4. Ядерные рецепторы.
Белки-рецепторы стероидных гормонов — факторы транскрипции. Каждый рецептор имеет область для связывания лиганда и участок, взаимодействующий с ДНК.
Вторые посредники (мессенджеры) передачи сигнала в клетке.
В настоящее время ко вторым посредникам относят цАМФ, цГМФ, ДАГ, ИФ3, ионы Са++.
Вторые посредники:
• оказывают воздействие на несколько групп протеинкиназ;
• изменяют активность нескольких групп фосфодиэстераз;
• способны непосредственно влиять на активность некоторых ионных каналов.
цАМФ:
• активируют протеинкиназу А (цАМФ-зависимую протеинкиназу);
• активирует фосфодиэстеразу, катализирующую цГМФ. Уровень цАМФ определяется соотношением активности протеинкиназы А и фосфосфодиэстеразы, гидролизующей цАМФ.
Значительное влияние на активность цАМФ оказывают производные арахидоновой кислоты.
цГТФ:
• активируют протеинкиназу G (цГМФ-зависимую протеинкиназу);
• активируют фосфодиэстеразу, катализирующую цАТФ;
• изменяют проницаемость ионных каналов (Na+ каналы и др.).
Инозитол-1, 4, 5-трифосфат (ИФ3).
Инозитол-1, 4, 5-трифосфат (ИФ3) или (ИТФ) способен связываться с кальциевыми каналами мембран цитоплазмы, эндоплазматического ретикулума и повышать их проницаемость. По градиенту концентрации Са++ входит в клетку через эти каналы, концентрация кальция в цитоплазме возрастает.
Диацилглицерол (ДАГ).
Диацилглицерол (ДАГ) за счет латеральной диффузии активирует мембранносвязанный фермент — протеинкиназу С (ПК-С).
Кальций (Са++).
Кальций, находясь в ионизированном состоянии:
• активирует фосфолипазу С;
• наряду с ДАГ, Са++ является активатором протеинкиназы С;
• связывает с кальмодулином;
активирует кальмодулинзависимые протеинкиназы.
11. Трансмембранный обмен…
Осуществляется за счет 2-х процессов:
—диффузии и осмоса.
Осмос — когда через мембрану движется растворитель из зоны с меньшей концентрацией в зону с большей концентрацией. Осмос поддерживает объем и форму клетки.
Диффузия— процесс проникновения веществ, растворимых в воде, по градиенту концентрации. Движущая сила при этом — разность концентраций.
Простая диффузия осуществляется либо через поры, которые есть в гидрофильных участках мембраны (фенестры, окна), либо через кинки — постоянно образующиеся временные дефекты мембраны. Простая диффузия не требует энергетических затрат, происходит за счет разности концентраций и осуществляется периодически, когда возникает разная концентрация.
Облегченная диффузия– ускоряет и усиливает перенос из зоны с большей концентрацией в зону с меньшей концентрацией по сравнению с простой диффузией. Она широко распространена в организме, так как хотя и сопровождается дополнительными энергетическими затратами, но не приводит к серьезным затратам энергии.
Облегченная диффузия — осуществляется за счет специфических переносчиков, создает условия для транспорта натрия, калия, хлора, моносахаридов, т.е. для некрупных молекул.
Различают 2 вида переносчиков:
1.Переносчики — белки, которые тем или иным способом переносят вещества через мембрану – за счет конформации (пространственного преобразования) молекул переносчика (сальтообразно).
2.Белки, которые образуют постоянные каналы, диаметр 0,3-0,6 нм, (так переносятся ионы натрия, калия, хлора).
Активный транспорт — транспорт веществ через мембрану, который осуществляется против градиента концентрации и требует значительных затрат энергии. Одна треть основного обмена тратиться на активный транспорт.
Активный транспорт бывает:
1. Первично-активый — такой транспорт, для обеспечения которого используется энергия макроэргов — АТФ, ГТФ, креатинфосфат. Например: Калиево-натриевый насос — важная роль в процессах возбудимости в клетке. Он вмонтирован в мембрану.
Калиево-натриевый насос — фермент калий-натриевая АТФаза. Этот фермент — белок. Он существует в мембране в виде 2-х форм:
-Е 1, Е 2
В ферментах существует активный участок, который взаимодействует с калием и с натрием. Когда фермент находится в форме Е 1, его активный участок обращен внутрь клетки и обладает высоким сродством к натрию, а значит способствует его присоединению (3 атома Na). Как только натрий присоединяется, происходит конформация этого белка, которая перемещает 3 атома натрия через мембрану и с наружной поверхности мембраны натрий отсоединяется. При этом происходит переход ферментаиз формы Е 1 в Е 2.
Е 2имеет активный участок, обращенный к наружной поверхности клетки, обладает высоким сродством к калию. При этом 2 атома К присоединяется к активному участку фермента, изменяется конформация белка и калий перемещается внутрь клетки. Это происходит с большой затратой энергии, так как фермент АТФаза постоянно расщепляет энергию АТФ.
2. Вторично-активный — это транспорт, который осуществляется тоже против градиента концентрации, но на это перемещение тратится не энергия макроэргов, а энергия электрохимических процессов, которая возникает при движении каких-либо веществ через мембрану при первично-активном транспорте.
Например: Сопряженный транспорт натрия и глюкозы, энергия — за счет перемещения натрия в калиево-натриевом насосе.
Классическим примером вторично-активного транспорта выступает натрий – Н (аш)-обменник — когда обмениваются натрий и водород (это тоже вторично-активный транспорт).
Способы транспортировки через мембрану:
1. Унипорт — это такой вид транспорта веществ через мембрану, когда переносчиком или каналом транспортируется одно вещество (Na-каналы)
2. Симпорт— это такой вид транспорта, когда 2 или более веществ в своем транспорте через мембрану взаимосвязаны и транспортируются вместе в одном направлении. (Na и глюкоза — в клетку) Это вид сопряженного транспорта
3. Антипорт— такой сопряжённый вид транспорта, когда его участники друг без друга не могут транспортироваться, но потоки идут навстречу друг другу (К-Na-насос-активный вид транспорта).
Эндоцитоз, экзоцитоз — как формы транспорта веществ через мембрану.
12. Ионные каналы…
Ионный канал состоит из нескольких субъединиц, их количество в отдельном ионном канале составляет от 3 до 12 субъединиц. По своей организации субъединицы, входящие в канал, могут быть гомологичными (однотипными), ряд каналов сформирован разнотипными субъединицами.
Каждая из субъединиц состоит из нескольких (три и более) трансмембранных сегментов (неполярные части, закрученные в α-спирали), из вне- и внутриклеточных петель и концевых участков доменов (представлены полярными областями молекул, формирующих домен и выступающих за пределы билипидного слоя мембраны).
Каждый из трансмембранных сегментов, вне- и внутриклеточных петель и концевых участков доменов выполняет свою функцию.
Так, трансмембранный сегмент 2, организованный в виде α-спирали, определяет селективность канала.
Концевые участки домена выступают в качестве сенсоров к вне- и внутриклеточным лигандам, а один из трансмембранных сегментов играет роль потенциалзависимого сенсора.
Третьи трансмембранные сегменты в субъединице ответственны за работу воротной системы каналов и т.д.
Ионные каналы работают по механизму облегченной диффузии. Движение по ним ионов при активации каналов идет по градиенту концентрации. Скорость перемещения через мембрану составляет 10 ионов в секунду.
Источник: cyberpedia.su