Характеристика профазы
Мейоз — один из типов клеточного деления, наряду с митозом. Мейоз включает два деления диплоидной клетки, результат которых — появление четырех гаплоидных половых клеток, у животных это гаметы, у растений и грибов споры. В любой гамете имеется половина первичного соматического набора хромосомы.
Иначе говоря, мейоз — это такое деление клетки, при котором происходит сокращение исходного количества хромосом вдвое: диплоидный набор (2n) превращается в гаплоидный (n).
Биологический смысл мейоза:
1) мейоз — залог постоянного образования половых клеток у животных, спор у грибов и растений;
2) в результате мейоза набор хромосом становится в два раза меньше, что способствует сохранению постоянства хромосомного набора в поколениях (диплоидный набор вновь восстанавливается после оплодотворения);
3) в процессе мейоза между гомологичными хромосомами происходит генетическая рекомбинация — кроссинговер, дающий новые «свежие» комбинации аллелей генов в половых клетках и новые комбинации признаков;
4) в мейозе идет независимое расхождение хромосом, в результате чего в половых клетках возникают новые сочетания хромосом, что также способствует появлению новых комбинаций признаков у отдельных особей.
Профаза 1 мейоза 1
Профаза мейоза 1 имеет пять последовательных стадий. Ниже мы рассмотрим подробности конъюгации и кроссинговера во всех пяти стадиях профазы мейоза 1: лептотене, зиготене, пахитене, диплотене и диакинезе.
1. Лептотена. Это так называемая стадия тонких нитей. Хромосомы тоненькие, удлиненные, «составлены» из двух сестринских хроматид, но они пока тесно сближены, отчего каждая хромосома кажется одиночной. Хромосомы конденсируются и видны в микроскоп. Они прикреплены концами к ядерной мембране. Итак, в ходе лептотены хромосомы «слипаются» в единое образование, становятся видимыми.
2. Зиготена. Гомологичные хромосомы объединяются. Вначале идет синапс — тесное сближение гомологов, это и обозначает переход от лептотены к зиготене. Концы хромосом-гомологов могут сближаться, а затем соединение от кончиков распространяется вдоль хромосом (впрочем, иногда бывает и наоборот). Образуется синаптонемальный комплекс.
1) Бивалент образуется при соединении двух гомологичных хромосом. Так как ДНК удваивалась в интерфазе, каждая из гомологичных хромосом будет состоять из пары хроматид.
2) Итак, бивалент — структура, содержащая четыре хроматиды, или (что аналогично) две гомологичные хромосомы. Используется и другое название — тетрада, при этом подчеркивается, что любая хромосома построена из пары сестринских хроматид.
3) Обратите внимание, что ниже на рисунках показан пример поведения в клетке лишь одной пары гомологичных хромосом (одного бивалента). Как вы понимаете, в разных клетках разное количество пар хромосом, значит, такие же процессы по аналогии будут идти с каждой парой хромосом.
4) На рисунке 1 две гомологичные хромосомы (бивалент) до сближения (очевидно, что состоят они из двух хроматид).
5) На рисунке 2 представлены биваленты при соединении двух гомологичных хромосом в профазе. Идет обмен участками хромосом. Проведем аналогию — на стадии зиготены хромосомы, как половые клетки при образовании зиготы в половом процессе, сближаются.
3. Пахитена. Стадия толстых нитей. Синапс завершен. Главное событие этой стадии — кроссинговер, или же перекрест между несестринскими хроматидами гомологичных хромосом. Перекресты проявляются в виде хиазм. Для запоминания можно применить «правило двух П»: пахитена и перекрест начинаются с буквы П.
1) В чем смысл кроссинговера? Материнские и отцовские хромосомы, построенные из пары хроматид, обмениваются участками.
2) Кроссинговер дает новые сочетания аллелей генов в хромосомах гамет. Помните, что в ходе кроссинговера не возникают новые аллели генов, он создает только их новые комбинации. Новые аллели возникают как результат генных мутаций.
3) Итак, при кроссинговере появляются хромосомы с новыми сочетаниями аллелей и, как следствие, новыми сочетаниями признаков, которые несут эти аллели.
4) Однако если в гомологичных хромосомах присутствуют две идентичные аллели генов, обмен ими не приведет к изменению признаков. Например, если идет перекрест между гомологичными хромосомами, в каждой из которых два одинаковых аллеля цвета глаз, то нового сочетания аллелей не образуется. Если же в одной хромосоме аллель А (карие глаза), а в другой а (голубые глаза), то кроссинговер приведет к обмену аллелями и образованию новых сочетаний аллелей в хромосомах.
4. Диплотена. Хромосомы в биваленте отталкиваются, они связаны только в местах хиазм. Идет окончание синапса, разрушение синаптонемального комплекса. У женщин на стадии диплотены хромосомы могут находиться в течение 10–15 лет, так как у них исходные клетки, из которых сформируются яйцеклетки, ооциты 1 порядка, начинают формироваться еще в ходе эмбрионального развития.
1) Итак, объединение хромосом заканчивается, они снова разделяются, и мы видим «ди» — две хромосомы, причем каждая имеет в своем составе две хроматиды.
2) В профазе 1 мейоза в отличие от профазы митоза, многие петли хромосом еще не конденсированы, в них идет транскрипция. К примеру, в ооцитах идет активный синтез РНК, синтез белков для питания будущего зародыша. Хромосомы с отходящими от них неконденсированными петлями хроматина называют хромосомами типа ламповых щеток (встречаются также у амфибий и других организмов).
5. Диакинез. Заканчивается конденсация хромосом. Они утолщены, отделены от ядерной мембраны. Бивалент явно состоит их двух гомологичных хромосом. Каждая из них — из двух хроматид. Набор хромосом и количество ДНК — 2n4c.
Источник: EgeVideo.ru
Митоз, его фазы, биологическое значение
Важнейшим компонентом клеточного цикла является митотический (пролиферативный) цикл.
представляет собой комплекс взаимосвязанных и согласованных явлений во время деления клетки, а также до и после него. Митотический цикл — это совокупность процессов, происходящих в клетке от одного деления до следующего и заканчивающихся образованием двух клеток следующей генерации. Кроме этого, в понятие жизненного цикла входят также период выполнения клеткой своих функций и периоды покоя. В это время дальнейшая клеточная судьба неопределенна: клетка может начать делиться (вступает в митоз) либо начать готовиться к выполнению специфических функций.
Основные стадии митоза.
1.Редупликация (самоудвоение) генетической информации материнской клетки и равномерное распределение ее между дочерними клетками. Это сопровождается изменениями структуры и морфологии хромосом, в которых сосредоточено более 90% информации эукариотической клетки.
2.Митотический цикл состоит из четырех последовательных периодов: пресинтетического (или постмитотического) G1, синтетического S, постсинтетического (или премитотического) G2 и собственно митоза. Они составляют автокаталитическую интерфазу (подготовительный период).
Фазы клеточного цикла:
1) пресинтетическая (G1). Идет сразу после деления клетки. Синтеза ДНК еще не происходит. Клетка активно растет в размерах, запасает вещества, необходимые для деления: белки (гистоны, структурные белки, ферменты), РНК, молекулы АТФ. Происходит деление митохондрий и хлоропластов (т. е. структур, способных к ауторепродукции). Восстанавливаются черты организации интерфазной клетки после предшествующего деления;
2) синтетическая (S). Происходит удвоение генетического материала путем репликации ДНК. Она происходит полуконсервативным способом, когда двойная спираль молекулы ДНК расходится на две цепи и на каждой из них синтезируется комплементарная цепочка.
В итоге образуются две идентичные двойные спирали ДНК, каждая из которых состоит из одной новой и старой цепи ДНК. Количество наследственного материала удваивается. Кроме этого, продолжается синтез РНК и белков. Также репликации подвергается небольшая часть митохонд-риальной ДНК (основная же ее часть реплицируется в G2 период);
3) постсинтетическая (G2). ДНК уже не синтезируется, но происходит исправление недочетов, допущенных при синтезе ее в S период (репарация). Также накапливаются энергия и питательные вещества, продолжается синтез РНК и белков (преимущественно ядерных).
S и G2 непосредственно связаны с митозом, поэтому их иногда выделяют в отдельный период — препрофазу.
После этого наступает собственно митоз, который состоит из четырех фаз. Процесс деления включает в себя несколько последовательных фаз и представляет собой цикл. Его продолжительность различна и составляет у большинства клеток от 10 до 50 ч. При этом у клеток тела человека продолжительность самого митоза составляет 1—1,5 ч, G2-периода интерфазы — 2—3 ч, S-периода интерфазы — 6—10 ч.
Стадии митоза.
Процесс митоза принято подразделять на четыре основные фазы: профазу, метафазу, анафазу и телофазу (рис. 1–3). Так как он непрерывен, смена фаз осуществляется плавно — одна незаметно переходит в другую.
В профазе увеличивается объем ядра, и вследствие спирализации хроматина формируются хромосомы. К концу профазы видно, что каждая хромосома состоит из двух хроматид. Постепенно растворяются ядрышки и ядерная оболочка, и хромосомы оказываются беспорядочно расположенными в цитоплазме клетки. Центриоли расходятся к полюсам клетки. Формируется ахроматиновое веретено деления, часть нитей которого идет от полюса к полюсу, а часть — прикрепляется к центромерам хромосом. Содержание генетического материала в клетке остается неизменным (2n2хр).

Профаза
К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе является характерной, но не обязательной для всех клеток особенностью.
Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК.
к следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе.
Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено — одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца, микротрубочки и кинетохоры хромосом.
С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут до 15 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы.
Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления.
Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме.
Прометафаза
Прометафаза
Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение.
В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом. В результате такого взаимодействия кинетохорные микротрубочки стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки.
В клетках млекопитающих прометафаза протекает, как правило, в течение 10—20 минут. В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона — около 30 минут.
Метафаза
Метафаза
В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи — к периферии клетки. В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка.
Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки.
В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума.
К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель.
Анафаза
Анафаза
Анафаза — самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид в направлении противоположных полюсов клетки. Хроматиды расходятся с равномерной скоростью достигающей 0,5—2 мкм/мин., при этом они часто принимают V-образную форму. Их движение обусловлено воздействием значительных сил, оценочно 10 дин на хромосому, что в 10 000 раз превышает усилие, необходимое для простого продвижения хромосомы через цитоплазму с наблюдаемой скоростью.
Как правило, расхождение хромосом в анафазе состоит из двух относительно независимых процессов называемых анафазой А и анафазой В.
Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За их движение при этом отвечают те же силы, что ранее удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения хроматид сопровождается сокращением длины деполимеризующихся кинетохорных микротрубочек. Причем их распад наблюдается преимущественно в области кинетохоров, со стороны плюс-концов. Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения сестринских хроматид, так как их движение прекращается при добавлении таксола или тяжёлой воды, оказывающих стабилизирующее воздействие на микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока остается неизвестным.
Во время анафазы В расходятся сами полюса деления клетки, и, в отличии от анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей разных видов. Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки.
Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны. Так в клетках млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к противоположным полюсам и продолжается вплоть до удлинения митотического веретена в 1,5—2 раза по сравнению с метафазным. В некоторых других клетках анафаза В начинается только после того как хроматиды достигают полюсов деления. У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным. В растительных клетках анафаза В отсутствует.
Телофаза
Телофаза
Телофаза рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки.
Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга.
Окончание телофазы преимущественно совпадает с разделением тела материнской клетки — цитокинезом. При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы.
Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки и деление путём образования клеточной пластинки. Плоскость деления клетки детерминируется положением митотического веретена и проходит под прямым углом к длинной оси веретена.
При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно ещё в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократимое кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса.
Деление путём образования клеточной пластинки начинается с перемещения мелких ограниченных мембраной пузырьков по направлению к экваториальной плоскости клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру — раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных микротрубочек веретена деления, образующих цилиндрическую структуру, называемую фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых мембранных пузырьков продолжается рост клеточной пластинки вплоть до её окончательного слияния с мембраной материнской клетки. После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки.
Источник: studfile.net
В профазу ядра входят после <32-периода интерфазы; после репликации ДНК в 5-периоде они содержат удвоенное ее количество по сравнению с исходным в брпериоде.[ ...]
В профазе в начале происходит конденсация и спирализация (скручивание) хромосом, в результате чего они становятся видимыми при микроскопии окрашенных препаратов. Увеличивается диаметр каждого завитка. Ядерная мембрана растворяется под действием ферментов, ядрышко исчезает. Центросома делится на две центриоли, после чего последние расходятся к полюсам клетки. Отмечается также фосфорилирование отдельных клеточных белков. Затем между полюсами начинает формироваться ахромати-новая фигура, похожая на веретено. Оно состоит из белка и РНК. К концу этой фазы ахроматиновая фигура вытягивается вдоль клетки, становясь веретеном. Структурно веретено представляет собой двухполюсную структуру, построенную из микротрубочек и различных белков. Хроматиды (сестринские хроматиды) удерживаются вместе центромерой. Длительность профазы составляет примерно 30-60 минут.[ …]
Каллоза имеет тонкую фибриллярную структуру и пронизана каналами. Являясь запасным полисахаридом» она участвует в обмене веществ при формировании микроспор.[ …]
А — иитерфаэа; Б — профаза; В —анафаза; Г — метафаза; 1 — диплоидные, 2 тетраплоидные. По Атабековой.[ …]
Крайне редко у гаплоидов в профазе первого деления мейоза в материнских клетках микроспор можно обнаружить двойной набор хромосом, очевидно, возникающий вследствие слияния клеток в ранней профазе или даже в археспории. Двойной хромосомный набор, по-видимому, образуется в анафазе археспория с формированием реституционного ядра при первом или втором: делениях мейоза.[ …]
А — нормального; Б — колхнцинового: / — профаза, 2 — метафаза, 3 — анафаза, 4 — телофаза.[ …]
По Устиновой.[ …]
Изменения, наблюдаемые у хромосом в течение всей профазы I, обусловливаются их сокращением в длину в результате спирализации хромонем. Подсчет числа хромосом легче проводить в период диакинеза, когда биваленты свободно лежат в полости ядра.[ …]
Второе мейотическое деление осуществляется в течение нескольких фаз (профаза II, метафаза II, анафаза II, телофаза II) и сходно с митотическим делением. В профазе II хромосомы вторичных спер-матоцитов остаются у полюсов. В метафазе II центромера каждой из двойных хромосом делится, обеспечивая каждую новую хромосому собственной центромерой. В анафазе II начинается формирование веретена, к полюсу которого двигаются новые хромосомы. В телофазе II второе мейотическое деление заканчивается, в результате чего каждый сперматоцит II порядка дает два спермати-да, из которых дифференцируются затем сперматозоиды. Как и во вторичном сперматоците, число хромосом в сперматиде является гаплоидным (п). Однако хромосомы сперматид являются одиночными, тогда как хромосомы вторичных сперматоцитов II являются двойными, будучи построенными из двух хроматид. Вторичное мейотическое деление является делением митотического типа (экваториальным делением). Оно разделяет двойные сестринские хроматиды и отличается от редукционного деления, в котором гомологичные хромосомы разделены. Единственное существенное отличие от классического митоза заключается в том, что здесь имеется гаплоидный набор хромосом.[ …]
Митоз совершается на протяжении четырех последовательных фаз, а именно: профазы, метафазы, анафазы и телофазы (рис. 59.).[ …]
Деление материнских клеток, как микроспор, так и мегаспор, по-видимому, блокируется, в результате чего семена либо не образуются вовсе, либо образуются в очень небольшом количестве (отсюда и происходит название вируса). Колдуэлл [320] отметил, что растения, инфицированные вирусами, обычно образуют меньше семян но сравнению со здоровыми растениями, и высказал предположение, что вызванные вирусами нарушения мейоза, вероятно, довольно широко распространены и, возможно, служат причиной того, что вирус не передается через семена. Было показано, например, [1653], что вирус штриховатой мозаики ячменя, который передается через семена ячменя, но не передается через семена кукурузы, оказывает мутагенное действие на растения кукурузы. Это действие выражалось в увеличении частоты утраты генов-маркеров семенами поколения 1< в тех случаях, когда для опыления использовалась пылхща растений, зараженных вирусом. При помощи тестов па инокуляцию вирус не удалось обнаружить ни в нормальных проростках, пи в проростках с пониженным содержанием хлорофилла поколения Р2.[ ...]
На стадии пахинемы, которая является наиболее долгой во времени стадией в мейотической профазе, происходит конденсация бивалентов и разделение каждой хроматиды надвое, в результате чего каждый бивалент представляет собой сложную спиральную структуру, состоящую из четырех сестринских хроматид (тетрад). В конце этой стадии начинается разделение спаренных хромосом-бивалентов. Теперь гомологичные хромосомы могут наблюдаться рядом. Поэтому в некоторых препаратах можно видеть четыре хромосомы, которые образуются в результате дупликации каждого гомолога, формирующего сестринские хроматиды. На этой стадии происходят обмены между гомологами и формирование хиазм.[ …]
На основании проведенных исследований митоз подразделяют на три периода: реорганизация профазы, при которой в интерфазном ядре происходят распад клеточных структур (ядрышка, ядерной оболочки) и синтез структурных элементов хромосом и митотического аппарата; деление и движение, при которых осуществляются метафаза и анафаза; реконструкция, при которой ■стадия телофазы завершается делением клетки — цитокинезом, или цитотомией.[ …]
Первое мейотическое деление начинается в первичном сперматоците и характеризуется длинной профазой, которая состоит из переходящих одна в другую профазы I и профазы II.[ …]
Во время интерфазы большая часть молекул ДНК находится в деспирализованном состоянии, но во время профазы митоза они становятся компактными и спирализованными, что происходит в результате изменений в связанных с ними белках, которые будут рассмотрены позднее (с. 449). После завершения митоза хромосомный материал деспирализуется и ядро входит в стадию интерфазы.[ …]
Форма хромосом разнообразна и специфична для данного вида организмов. В период профазы хромосомы состоят из двух продольных половинок — хроматид. В свою очередь, каждая хроматида состоит из нитей — хромонем, расположенных параллельно продольной оси хромосомы. Количество нитей ДНК, а также взаиморасположение ДНК и белков в хромосомах до настоящего времени не ясно. Хромосомы дифференцированы но длине; в отдельных участках хромосом (локусах) расположены определенные геыы, несущие информацию, соответствующую образованию одного белка.[ …]
Гаплоидные растения высокостерильны, поскольку их хромосомная структура препятствует нормальному течению мейоза. В профазе мейоза в ядрах гаплоидов можно наблюдать образование петель в результате соединения сегментов одной и той же хромосомы, а также неправильное или частичное соединение различных хромосом (рис. 68).[ …]
В приготовленных таким образом препаратах не менее, чем на 10 полях зрения подсчитывалось количество клеток в интерфазе, профазе, метафазе, анафазе и телофазе.[ …]
На рисунке 50 представлен процесс изменения ядра при митозе в клетках корешков алоэ. Весь цикл деления клетки подразделяется на профазу, метафазу, анафазу, телофазу и цитокинез. Границы между этими фазами провести крайне трудно, поскольку митоз представляет собой единый процесс, в ходе которого смена фаз осуществляется постепенно.[ …]
Течение мейоза у различных гаплоидов во многом сходно, что позволяет дать общее описание этого процесса. В гаплоидных клетках в профазе I наблюдается несоответствие хромомер спаренных нитей вследствие ассоциации негомологичных хромосом (см. рис. 68). Спаривание негомологщных хромосом непродолжительно, не дает хиазм и вскоре заканчивается их разъединением. В этот период число хромосом в материнских клетках микроспор может быть и менее гаплоидного набора (п) из-за конъюгации двух или даже т рех хромосом.[ …]
Опыты по гипофизэктомии позволили установить критиче» скую стадию в сперматогенезе у акулы S. caniculus (Dodd fej: al., I960).[ …]
В процессе митоза происходит удвоение числа хромосом, которые поровну распределяются между двумя дочерними клетками. Следовательно, количество ДНК должно быть удвоено на какой-то стадии до того, как во время профазы становится видимым двойной набор хромосом. Точную стадию синтеза ДНК можно определить двумя способами: 1) путем введения на различных стадиях роста в растительные ткани, например в ткань кончиков корней, радиоактивного тимидииа (одного из оснований ДИК) и определения времени, в течение которого он включится в состав ДНК; 2) путем определения с помощью спектрофотометрии времени, в течение которого количество ДНК в ядре удвоится. Эти методики позволяют выявить стадию синтеза ДНК, обозначаемую как Б-фаза. Периоды до и после Б-фазы, во время которых не происходит синтеза ДНК, обозначаются как Ог и Ог-периоды (рис. 1.1). Продолжительность йр и С2-периодов в клетках различных типов может значительно варьировать.[ …]
Второе деление мейоза. Процесс этот по сути — митоз в клетке с гаплоидным числом хромосом; он проходит быстро, поскольку подготовлен уже в период первого деления мейоза. Парные сестринские хроматиды, связанные в центромерных участках, пройдя профазу и метафазу, переходят к анафазе, где разъединяются и расходятся к полюсам клетки. Следовательно, при мейозе в результате двукратного деления из одной диплоидной клетки возникают четыре гаплоидных, отличающихся друг от друга (благодаря кроссинговеру) по генетической структуре.[ …]
В стадии телофазы I хромосомы достигают полюсов, чем заканчивается первое мейотическое деление. После телофазы I наступает короткая интерфаза (интеркинез), в которой хромосомы деспи-рализуются и становятся диффузными, или телофаза I переходит прямо в профазу II второго мейотического деления. Ни в одном, ни в другом случае репликации ДНК не отмечается. После первого мейотического деления клетки называют сперматоцитами II порядка. Количество хромосом в каждой такой клетке снижается от 2п до п, но содержание ДНК еще не изменяется.[ …]
Мейоз в сперматогенезе протекает в несколько стадий (фаз). Между делениями имеются две интерфазы. Процесс мейоза очень динамичен, поэтому микроскопические различия между разными стадиями отражают скорее не характер самих стадий, а скорее свойства хромосом на разных стадиях (рис. 82). Интерфаза I характеризуется тем, что в ней происходит репликация хромосом (удвоение ДНК), которая к началу ранней профазы I почти полностью завершается.[ …]
Локализация их в различных хромосомах неодинакова. Эухроматиновые участки хромосом во время интерфазы находятся в деспирализованном состоянии и вновь спирализуются в профазе следующего деления. Поскольку эти участки содержат основной комплекс генов, их считают активными зонами хромосом.[ …]
Это может быть следствием ряда причин: может быть нарушение процесса деления связано с действием на процессы роста, предшествующие делению; возможно нарушается образование веретенного аппарата, что свойственно карбаматам, так как веретенный аппарат образуется в профазе (2); возможно удлиняется период профазы, так как резкое возрастание МИ связано в основном с увеличением числа профазных фигур, и, наконец, возможно под действием гербицидов приостанавливается деление части клеток на каком-то этапе, и это стимулирует вступление в деление других покоящихся центров (защитная реакция). Все эти предположения могут относиться к механизму действия данных гербицидов. Нас в данном исследовании интересует непосредственное действие карбина и три-аллата на наследственные структуры — хромосомы. Снижение МИ через 44 часа на вариантах с гербицидами до уровня контроля и отсутствие разрывов хромосом (отсутствие анафазных аберраций) можно рассматривать как положительный факт, позволяющий нам предположить, что карбин и триаллат не являются серьезными мутагенными агентами в посевах ячменя.[ …]
Описание митоза, сделанное после исследования клетки с помощью светового микроскопа, может быть дополнено наблюдениями ультраструктуры ядра под электронным микроскопом. Согласно электронной микроскопии первые изменения структуры ядра осуществляются уже в ранней профазе. При этом в процессы перестройки ядра последовательно вовлекаются все его компоненты. Первоначальные изменения проявляются в конденсировании хроматина во всем объеме ядра. По мере нарастания этого процесса возникают митотические хромосомы с максимальной плотностью упаковки в них фибрилл дезоксииуклеопротеида.[ …]
Собственно митоз. Митотический аппарат, под которым понимают всю совокупность структур, составляющих ахроматическую фигуру митоза (астросфера, окружающая центриоль, и митотическое веретено, или веретено деления), не является постоянной органеллой клетки. Он формируется в поздней профазе или в ранней метафазе. При подготовке к делению клетка обеспечивает синтез основной массы веществ, идущих на построение митотического аппарата, занимающих значительную часть деля- щейся клетки, а также богатых энергией и регулирующих деятельность веретена.[ …]
Совокупность всех преобразований, связанных с делением ядра, составляет м и тоти-ческий цикл, или митоз. У водорослей, как и у других растений, реорганизация ядра следует в строгой последовательности в несколько этапов (фаз). Всего различают 5 фаз: интерфаза, или состояние покоя (самая продолжительная по времени), профаза, метафаза, анафаза и телофаза. Изтелофазы ядро снова переходит в интерфазу.[ …]
Длительность митоза первичного ядра в пыльцевом зерне зависит от условий среды (температура, влажность воздуха), характера питания микроспор и особенностей развития тапе-; тума пыльника. Так, в пыльцевых зернах традесканции длительность отдельных фаз митоза в период гаметогенеза следующая (при температуре 30°С): профазы — 30—34 ч (наибольшая часть этого времени приходится на раннюю профазу), метафазы и анафазы — по 0,5, телофазы — 0,2 ч. Весь митоз длится от 31,2 до 35,2 ч (Бкшоп, 1950).[ …]
Микротрубочки играют также какую-то роль в определении того, где будет формироваться клеточная пластинка и где она сольется со стенкой родительской клетки. В покоящихся клетках микротрубочки лежат в периферической цитоплазме в непосредственной близости от плазмалеммы. В клетках, в которых должно начаться деление, но ядро еще не вступило в профазу, такие «пристеночные микротрубочкн» исчезают, а в периферической цитоплазме вблизи продольных стенок появляется состоящий из большого числа трубочек тяж, который располагается под прямым углом к оси клетки (рис. 1.7). Этот «препрофазный тяж» огибает цитоплазму по периферии в средней части клетки. Когда клеточная пластинка полностью разовьется, она сливается с боковыми стенками родительской клетки так, что делит этот препрофазный тяж почти равномерно между двумя дочерними клетками. Следовательно, можно считать, что препрофазный тяж фактически предопределяет положение и ориентацию клеточной пластинки.[ …]
Хромосомы дифференцируются на специфические участки — структурные элементы хромонем (наиболее уплотненные участки хромонем) —хромомеры, которые в световом микроскопе имеют вид темноокрашенных гранул, располагающихся по длине хромосомы в определенном порядке.[ …]
Каждое ядро обычно содержит одно, но иногда и большее количество ядрышек. Обычно они бывают диаметром 1—2 мкм, хотя встречаются и более крупные. Ядрышки состоят из очень плотного вещества и лишены мембранного барьера. В них, в отличие от хроматина, скапливается не ДНК, а РНК. Во время ядерного деления ядрышки претерпевают циклические преобразования, связанные с разрыхлением, потерей красящей способности и диспергированием (рассеиванием) во время первой стадии ядерного деления (профазы) и конденсацией, восстановлением красящей способности с полной реконструкцией своего облика к концу этого процесса (в тело-фазе).[ …]
Анализ всех известных генотипических различий в генетической рекомбинации у организмов-эукариотов свидетельствует о том, что они обусловлены инбридингом, межвидовыми скрещиваниями, видовыми, популяционными и индивидуальными особенностями. Видовые, индивидуальные и другие особенности касаются уровня рекомбинации. Известны также специфические гены, мутации которых поражают способность к рекомбинации. Они гены у эукариот влияют на спаривание хромосом в мейозе, вследствие чего последние теряют способность к формированию пар в первой профазе мейоза, или на формирование хиазм. У бактерий известно несколько генов гес, продукты которых контролируют генетическую рекомбинацию бактерий. У фагов также обнаружены генетические системы, контролирующие их рекомбинацию. В совокупности все эти данные свидетельствуют о подверженности рекомбинаций генетическому контролю.[ …]
В телофазу хромосомы постепенно деспирализуются, формируются новые дочерние ядра. Собственно процесс деспи-рализации начинается еще в ранней телофазе, когда на полюсах образуются две компактные группы хромосом. Далее хромосомы постепенно утрачивают четкость контуров. Одновременно с деспирализадией происходит образование оболочки у вновь возникших дочерних ядер в результате скопления цистерн эндоплазматической сети вокруг хромосом. Процесс реконструкции дочерних ядер как бы повторяет ход профазы в обратном порядке.[ …]
Последняя треть XIX в. ознаменовалась крупнейшими открытиями в области цитологической науки: в растительной клетке были обнаружены особые внутриклеточные структуры — хромосомы, а также описаны способы деления ядра. Тогда же зоологи Шнейдер (1873) и Бючли (1874) наблюдали картину деления ядра в животных клетках (черви), но не сумели обобщить и оценить виденного ими явления. Эти интересные открытия были сделаны русским ученым И. Д. Чистяковым в 1874 г. Со времени выхода в свет его классического труда о структуре и делении ядра растительной клетки началось развитие цитологии в России. В 1875 г. Э. Страсбургером также было детально описано деление ядра оплодотворенной яйцеклетке ели. Им же были предложены термины: «профаза», «метафаза», «анафаза», «гаплоидное» и «диплоидное» число хромосом.[ …]
Источник: ru-ecology.info
Определение митоза
Митоз — это тип деления клеток, при котором одна материнская делится, чтобы произвести две новые генетически идентичные дочерние. В контексте клеточного цикла митоз является частью процесса деления, происходящего последовательно, при котором ДНК ядра разделяется на два равных набора хромосом:
- Во время митоза одна клетка делится один раз, чтобы сформировать две одинаковые.
- Главной целью митоза является рост и замена изношенных клеток.
- Если не исправить ошибки, возникшие во время митоза, это вызовет изменения в ДНК, что приведёт к генетическим нарушениям.
В процессе продолжительного развития и роста митоз наполняет организм клетками, а на протяжении всей жизни он заменяет старые изношенные новыми. Для одноклеточных эукариот, каковыми являются дрожжи, митотические деления фактически являются основой размножения, добавляя в популяцию новых особей.
Митоз у животных впервые смог открыть В.Флемминг в 1882 году, а в 1888 году Э. Страсбургер у растений.
Стадии деления клеток
Митоз состоит из четырёх основных стадий. Они имеют чёткую очерёдность:
- профаза;
- метафаза;
- анафаза;
- телофаза.
Здесь часто возникают расхождения, поскольку некоторые учебники перечисляют 5, разбивая профазу на раннюю фазу (интерфазу) и позднюю фазу (называемую прометафазой). Они происходят в строгом последовательном порядке, а также цитокинез, который начинается в анафазе или телофазе. Для того чтобы стало понятно, какими характеристиками обладают фазы, следует рассмотреть кратко сущность каждой из них.
Интерфаза
Почти 80% продолжительности жизни клетки тратится в межфазной стадии митоза. На этом этапе деление нет, но происходит период роста и подготовка к делению. ДНК дублируется во время этой фазы, создавая две копии каждой цепи, называемой хромосомой — молекулой ДНК, которая несёт всю или часть наследственной информации организма.
Можно перечислить стадии, характерные для интерфазы. Они разделены на:
- фазу G1,
- фазу S,
- фазу G2.
Фаза G1 — это период до синтеза ДНК, в течение которого клетка увеличивается в размерах. Во время фаз G1 клетки растут и контролируют свою среду, чтобы определить, следует ли им инициировать ещё один раунд деления.
Профаза
После подготовительной стадии интерфазы профазу считают первой истинной стадией митотического процесса. Во время ранней профазы клетка начинает разрушать одни структуры и создавать другие, готовясь к делению хромосом. Дублированные хромосомы из межфазной стадии конденсируются. Это означает, что они уплотняются и плотно наматываются. Ядерная оболочка разрушается, и на краях делящейся клетки формируется аппарат, известный как митотическое веретено.
Шпиндель состоит из сильных белков, называемых микротрубочками, которые являются частью «скелета» клетки и управляют делением посредством удлинения. Шпиндель постепенно удлиняется во время профазы. Его смысл заключается в организации хромосом и их перемещении во время деления.
К концу профазы ядерная оболочка разрушается, и микротрубочки достигают от каждого полюса до экватора. Кинетохоры, специализированные области в центромерах хромосом, — области ДНК, где сестринские хроматиды наиболее тесно связаны — прикрепляются к типу микротрубочек, называемых волокнами кинетохор. Эти волокна взаимодействуют с полярными волокнами веретена, соединяя кинетохоры с полярными волокнами, что стимулирует миграцию хромосом к ядру. Эту часть процесса иногда называют прометафазой, потому что она происходит непосредственно перед метафазой.
Метафаза
В самом начале метафазной стадии пары конденсированных хромосом выстраиваются вдоль экватора вытянутой клетки. Поскольку они уплотнены, они могут двигаться легче, не запутываясь.
Во время метафазы ядерная мембрана полностью исчезает. В клетках животных две пары центриолей располагаются на противоположных полюсах, а полярные волокна продолжают простираться от полюсов к центру. Хромосомы движутся случайным образом до тех пор, пока они не прикрепятся с обеих сторон их центромер к полярным волокнам.
Хромосомы располагаются на метафазной пластинке под прямым углом к полюсам веретена и удерживаются там равными силами полярных волокон, оказывающих давление на центромеры хромосом. Метафазная пластинка не является физической структурой — это просто термин для плоскости, в которой расположены хромосомы.
Прежде чем перейти к следующей стадии анафазы, происходит проверка, все ли хромосомы находятся на метафазной пластинке, а кинетохоры правильно прикреплены к микротрубочкам. Это называется контрольной точкой шпинделя. Она обеспечивает равномерное распределение пар хромосом, также называемых сестринскими хроматидами, между двумя дочерними на стадии анафазы. Если хромосома неправильно выровнена или прикреплена, деление прекратится до тех пор, пока проблема не будет решена.
В редких случаях она не прекращает деление, и при митозе допускаются ошибки. Это может привести к изменениям ДНК, которые потенциально могут привести к генетическим нарушениям.
Анафаза
Во время анафазы сестринские хроматиды тянутся к противоположным полюсам (концам) вытянутой клетки. Белковый «клей», удерживающий их вместе, разрушается, позволяя им развалиться. Это означает, что дубликаты ДНК клетки оказываются по обе стороны от клетки и готовы полностью разделиться. Каждая сестринская хроматида теперь является собственной «полной» хромосомой. Теперь они называются дочерними хромосомами. На этом этапе микротрубочки становятся короче, что позволяет начать процесс разделения клеток.
Дочерние хромосомы проходят через механизм веретена, чтобы достичь противоположных полюсов. Когда хромосомы приближаются к полюсу, они сначала мигрируют центромерами, а волокна кинетохор сокращаются.
Телофаза
На заключительной стадии телофазы деление практически завершено. Оболочка ядра, которая ранее была разрушена, чтобы позволить микротрубочкам получить доступ и рекрутировать хромосомы к экватору делящейся клетки, превращается в две новые ядерные оболочки вокруг разделённых сестринских хроматид. Телофазу ещё называют обратной профазой, потому что во время профазы происходит спирализация и укорочение хромосом, а во время телофазы — деспирализация
Полярные волокна продолжают удлиняться, и ядра начинают формироваться на противоположных полюсах, создавая ядерные оболочки из оставшихся частей ядерной оболочки родительской клетки, плюс части эндомембранной системы. Митотический веретен распадается на свои строительные блоки, и образуются два новых ядра — по одному на каждый набор хромосом. Во время этого процесса вновь появляются ядерные мембраны и ядрышки, и хроматиновые волокна хромосом раскрываются, возвращаясь к своей прежней нитевидной форме.
После телофазы митоз практически завершён — удвоение произошло. Тем не менее, деление не завершено, пока не произойдёт цитокинез.
Цитокинез
Цитокинез — это деление цитоплазмы, начинающееся до окончания анафазы и заканчивающееся вскоре после телофазной стадии митоза.
Во время цитокинеза кольцо белков, называемых актином и миозином (те же белки, что и в мышцах), сжимает вытянутую клетку в две совершенно новые. Группа нитей из белка под названием актин ответственна за защемление, создавая складку, так называемую борозду расщепления.
Процесс отличается в растениях, потому что они имеют клеточную стенку и слишком жёсткие, чтобы делиться таким образом. В растительных клетках структура, называемая клеточной пластинкой, формируется в их середине, разделяя её на две дочерние с новой стенкой между ними.
В этот момент цитоплазма поровну поделена между двумя новыми клетками, каждая из которых генетически идентична, содержит собственное ядро и полную копию ДНК организма. Теперь они начинают свой собственный путь и могут сами повторять процесс митоза в зависимости от того, кем они становятся.
Таблица по фазам
| Фаза | Процесс |
| Профаза | Хромосомы становятся видимыми в результате спирализации. Каждая хромосома состоит из двух хроматид. Ядерная оболочка и ядрышко разрушаются. |
| Метафаза | Хромосомы располагаются по экватору, образуется двухполюсное митотическое веретено. |
| Анафаза | Центромеры делятся и хроматиды расходятся с помощью нитей митотического веретена к полюсам. |
| Телофаза | Исчезает митотическое веретено. Вокруг разошедшихся хромосом образуются новые ядерные оболочки. Образование двух клеток. |
Последовательная схема
Биологическое значение
Митоз является одним из способов репликации в биологии. У одноклеточных организмов митоз является единственной жизнеспособной формой размножения. У сложных организмов роль митоза заключается в восстановлении повреждённых тканей и помощи организму осуществлять рост. Основными целями митоза являются:
- Бесполое размножение. В одноклеточном организме (амёбе) митоз — это способ размножения.
- Рост. По мере старения растений и животных, большинство из них увеличивается в размерах. Митоз создаёт клетки, необходимые для увеличения массы тела, а также большее их число, чтобы справиться с ростом, например, новые клетки крови. Особенностью является то, что не все клетки человеческого организма подвергаются митозу или другим формам размножения, например, нервные и мышечные.
- Регенерация. Некоторые животные могут регенерировать части тела. Когда организм получает травму, возникает процесс деления, чтобы заменить повреждённые клетки. Этот ремонт особенно важен для кожи и кровеносных сосудов, которые защищают и насыщают кислородом мышцы и органы в организме. Митоз также помогает заменить кровь, потерянную через рану. В некоторых организмах, таких как ящерицы, митоз может заменить целые потерянные конечности, такие как хвосты или ноги.
- Ошибки. Поскольку митоз так важен для восстановления и роста, при появлении ошибок возникают серьёзные проблемы. Одной из основных разновидностей осложнений является рак. Мутации в ДНК могут происходить во время процесса митоза, и если они не пойманы, могут возникнуть раковые клетки. Ошибки также могут возникать во время развития плода, что приводит к хромосомным расстройствам, таким как синдром Дауна и синдром Тёрнера.
Эукариоты и прокариоты
Митоз встречается только в эукариотах. Прокариоты, в которых отсутствует ядро, делятся с помощью другого процесса, называемого бинарным делением. Митоз варьируется между организмами. Например, в организме животных происходит открытый митоз, где ядерная оболочка разрушается до того, как хромосомы отделяются, тогда как грибы подвергаются закрытому митозу, где хромосомы делятся внутри неповреждённого ядра.
Большое количество клеток животных претерпевает изменение формы, известное как округление митотических клеток, чтобы принять почти сферическую морфологию в начале митоза. Большинство клеток человека получаются путём деления митотических клеток. Важные исключения включают гаметы — сперматозоиды и яйцеклетки, которые получаются в процессе мейоза.
Источник: nauka.club
Понятие «митотический цикл»
Все процессы, которые происходят в клетке, начиная от одного деления до другого, и заканчивая получением двух дочерних клеток, называется митотическим циклом. Жизненным циклом клетки также является состояние покоя и период выполнения своих прямых функций.
К основным стадиям митоза относятся:
- Самоудвоение или редупликация генетического кода, который передаётся от материнской клетки к двум дочерним. Процесс влияет на структуру и образование хромосом.
- Клеточный цикл – состоит из четырёх периодов: пресинтетического, синтетического, постсинтетического и, собственно, митоза.
Первые три периода (пресинтетический, синтетический и постсинтетический) относятся к интерфазе митоза.
Процесс непосредственного деления клетки, митоз, происходит в четыре фазы, соответствуя такой последовательности:
- Профаза;
- Метафаза;
- Анафаза;
- Телофаза.
Рис. 1. Фазы митоза
Познакомиться с кратким описанием каждой фазы можно в таблице «Фазы митоза», которая представлена далее.
Нетипичные формы митоза
В природе иногда встречаются и нетипичные формы митоза:
- Амитоз – способ прямого деления ядра, при котором сохраняется строение ядра, ядрышко не распадается, хромосомы при этом не просматриваются. В результате получаем двухъядерную клетку.
Рис. 2. Амитоз
- Политения – кратно увеличиваются клетки ДНК, но без увеличения содержания хромосом.
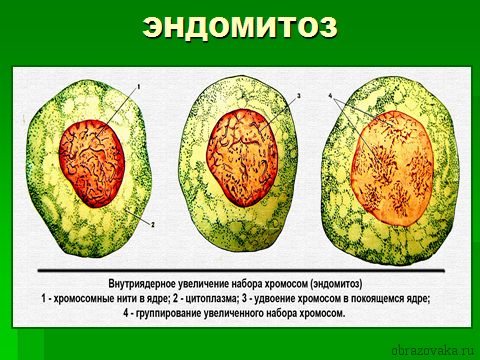
- Эндомитоз – в ходе процесса после репликации ДНК нет разделения хромосом на дочерние хроматиды. При этом число хромосом увеличивается в десятки раз, возникают полиплоидные клетки, которые могут привести к мутации.
Рис. 3. Эндомитоз
Источник: obrazovaka.ru