Линейные хромосомы
Рубрики
Источник: kaz-ekzams.ru
Линейные хромосомы бактерий
Афоризм Жака Моно: «То, что верно для E. coli, – верно и для других бактерий (слона)» получил широкое распространение. К счастью, на деле все обстоит не так скучно. До недавнего времени общепринятым было представление о кольцевой структуре бактериальных хромосом. Однако в 1989 г. была впервые описана у спирохеты Borrelia burgdorfery линейная бактериальная хромосома, которую идентифицировали с помощью электрофореза в импульсном электрическом поле. Размер этого генома составлял всего 960 т.п.о. Вскоре было обнаружено, что линейная и кольцевая хромосомы сосуществуют одновременно у Agrobacterium tumefaciens, а у грамположительных бактерий рода Streptomyces, обладающих одним из самых больших бактериальных геномов (8000 т.п.о.), имеется одна линейная хромосома. Представитель актиномицетов Rhodococcus fascians также, по-видимому, обладает линейной хромосомой. Линейные хромосомы у бактерий часто сосуществуют с линейными плазмидами и широко распространены в природе.
Линейные хромосомы и плазмиды наиболее хорошо изученных бактерий рода Streptomyces содержат концевые инвертированные повторы (terminal inverted repeats – TIRs), с которыми ковалентно связаны концевые белки (TP).
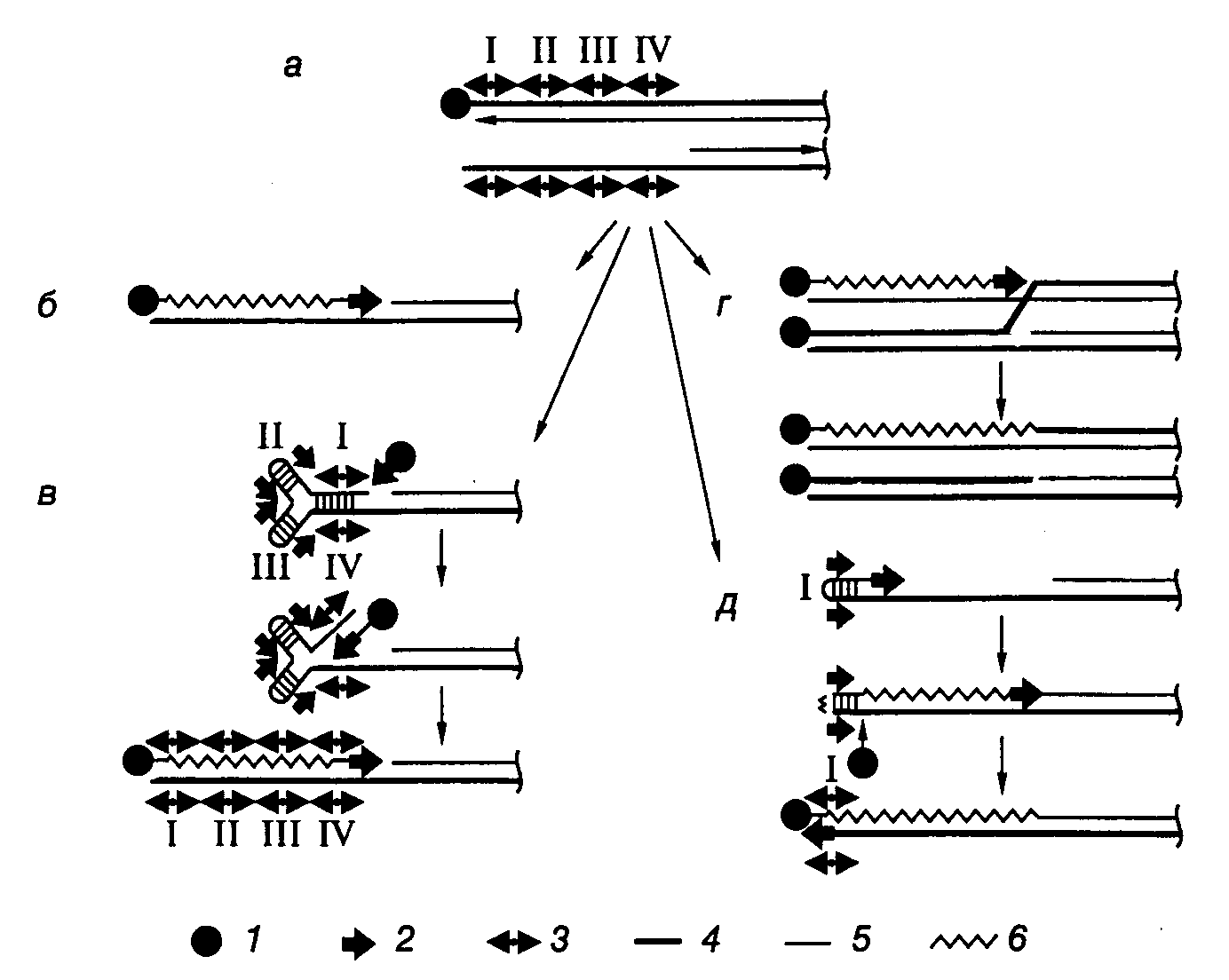
смотря на то что подобные структуры характерны для хромосом аденовирусов и бактериофага 29 Bacillus subtilis, механизм репликации хромосом стрептомицетов существенно отличается от такового вирусных геномов. Если у вирусов синтез ДНК инициируется на конце хромосомы с использованием в качестве затравки TP, ковалентно связанного с нуклеотидом, и продолжается через весь геном до его конца, то репликация хромосомы и линейных плазмид стрептомицетов начинается с внутренней области начала репликации oriC. Синтез ДНК распространяется в обе стороны от области начала репликации по стандартному полуконсервативному механизму и завершается на концах линейных молекул ДНК с образованием 3’-концевых брешей (рис. I.50,а). Наиболее простым решением проблемы заполнения этой бреши могла бы быть прямая инициация репликации теломерных участков хромосом с TP-белка, ковалентно связанного с инициирующим нуклеотидом, что имеет место у аденовирусов (см. рис. I.50,б). Действительно, стрептомицеты используют ТР для репликации теломерных участков, однако механизм распознавания теломер в данном случае существенно отличается. В настоящее время рассматриваются три модели заполнения брешей в теломерных участках линейных хромосом бактерий.

Рис. I.50. Модель достройки теломерных участков хромосом и плазмид Streptomyces
а– структура теломеры после репликации: верхняя цепь ДНК полностью реплицирована, в нижней имеется одноцепочечная брешь, обозначены четыре палиндромные последовательности нуклеотидов;б– маловероятный механизм с участием концевого белка и ДНК-полимеразы;в–д– альтернативные модели репликации, основанные на других механизмах.1– концевой белок,2– ДНК-полимераза,3 – палиндром,4– родительская цепь ДНК,5– дочерняя цепь,6– репаративный синтез
В соответствии с первой моделью одноцепочечный участок теломеры, содержащий TIR-последовательность, образует концевую шпильку путем комплементарных взаимодействий нуклеотидов внутренних участков бреши и 3’-концевых нуклеотидов (см. рис. I.50,в). В этом случае синтез ДНК, репарирующий одноцепочечную брешь, инициируется на двухцепочечном участке, образованном палиндромными последовательностями I-IV, с участием ТР и ДНК-полимеразы и продолжается вдоль 3’-концевого одноцепочечного участка хромосомы. Согласно второй модели ТР инициирует репликацию на полностью двухцепочечной дочерней ДНК, вытесняя 5’-концевую цепь родительской ДНК, с которой связан ТР (см. рис. I.50,г). Вытесняемая цепь далее спаривается с выступающим 3’-концом хромосомы, после чего такая разветвленная структура разрешается с помощью гомологичной рекомбинации. Эта модель предполагает участие в заполнении брешей белка RecA (для переноса цепи ДНК) и продуктов генов ruv (для разрешения структуры Холидея), что подтверждается генетическими данными. В третьей модели одноцепочечный палиндром I образует шпильку, 3’-конец которой служит затравкой для синтеза ДНК, в результате которого заполняется брешь (см. рис. I.50,д). ТР образует одноцепочечный разрыв напротив первоначального 3’-конца, который является затравкой для последующего синтеза ДНК. В результате шпилька разворачивается и восстанавливается структура теломеры. Эта модель аналогична модели «катящейся шпильки», предложенной для объяснения механизма репликации генома парвовирусов. В данной модели роль ТР отличается от его функций в качестве белка-затравки в рассмотренных выше примерах.
Неизвестно, как много форм линейных бактериальных хромосом существует в природе. Не изучены и таксономические проблемы, связанные с топологией хромосом в царстве эубактерий. Если каждый тип хромосом характерен для отдельного таксономического домена, то можно предполагать, что топология хромосом играет важную роль в эволюции бактерий. Альтернативно топологические взаимопревращения хромосом могут быть относительно частыми событиями, а линейные и кольцевые хромосомы присутствуют только у близких видов бактерий. Нестабильность хромосом стрептомицетов (образование протяженных делеций и амплификация последовательностей нуклеотидов) недавно стали связывать с перестройками в их концевых участках, часть из которых сопровождалась образованием кольцевых хромосом. Таким образом, эволюционная роль топологии бактериальных хромосом может быть определена только в результате будущих исследований.
Источник: StudFiles.net
История изучения хромосом
В ряде экспериментов, начатых в середине 1880-х, Теодор Бовери определенно продемонстрировал, что хромосомы являются векторами наследственности. Его двумя принципами были последовательность хромосом и индивидуальность хромосом. Второй принцип был очень оригинальным. Вильгельм Ру предположил, что каждая хромосома несет разную генетическую нагрузку. Бовери смог протестировать и подтвердить эту гипотезу. При помощи повторного открытия, сделанного в ранней работе Грегора Менделя, в начале 1900-х, Бовери смог отметить связь между правилами наследования и поведением хромосом. Бовери повлиял на два поколения американских цитологов: среди них Эдмунд Бичер Уилсон, Уолтер Саттон и Теофилус Пейнтер (в действительности Уилсон и Пейнтер работали с ним).
В своей знаменитой книге «Клетка в развитии и наследственности» Уилсон связал вместе независимую работу Бовери и Саттона (около 1902 г.), назвав хромосомную теорию наследственности «Теорией Саттона-Бовери» (имена иногда переставляются местами). Эрнст Мэйр отмечает, что теория была горячо оспорена некоторыми знаменитыми генетиками, например, Уильямом Бэйтсоном, Вильгельмом Йохансеном, Ричардом Гольдшмидтом и Т.Х. Морганом, все они обладали довольно догматичным складом ума. В итоге полное доказательство было получено от хромосомных карт в собственной лаборатории Моргана.
Прокариоты и хромосомы
Прокариоты – бактерии и археи – обычно имеют одну круглую хромосому, но существует много вариаций.
В большинстве случаев размер хромосом бактерий может варьироваться от 160000 пар оснований в эндосимбиотической бактерии Candidatus Carsonella ruddii до 12200000 пар оснований в обитающей в почве бактерии Sorangium cellulosum. Спирохеты рода Borrelia являются замечательным исключением из этой классификации вместе с такими бактериями, как Borrelia burgdorferi (причина болезни Лайма), содержащими одну линейную хромосому.
Структура в последовательностях
Хромосомы прокариотов имеют меньшую структуру на основе последовательности, чем эукариоты. Бактерии обычно обладают одной точкой (происхождение дублирования), откуда начинается дублирование, в то время как некоторые археи содержат множество точек происхождения дублирования. Гены в прокариотах часто организованы в опероны и обычно не содержат интроны, в отличие от эукариотов.
Упаковка ДНК
Прокариоты не имеют ядер. Вместо этого их ДНК организована в структуру под названием нуклеоид. Нуклеоид – это отдельная структура, которая занимает определенный участок клетки бактерии. Однако эта структура динамична, поддерживается и трансформируется действиями похожих на гистон белков, которые связываются с бактериальной хромосомой. В археях ДНК в хромосомах даже более организованы, при этом ДНК упакованы в структуры, аналогичные нуклеосомам эукариотов.
Бактериальные хромосомы склонны привязываться к плазменной мембране бактерии. В молекулярном биологическом приложении это позволяет ее изоляцию от ДНК плазмида посредством центрифугирования лизированной бактерии и осаждения мембран (и присоединенной ДНК).
Хромосомы прокариотов и плазмиды являются, как ДНК эукариотов, в целом сверхспиральными. ДНК должна выделиться сначала в ослабленном состоянии для доступа к транскрипции, регулированию и дублированию.
В эукариотах
Эукариоты (клетки с ядрами, обнаруживаемые в растениях, дрожжах и животных) обладают большими линейными хромосомами, содержащимися в клеточном ядре. Каждая хромосома имеет одну центромеру, одно или два плеча выступают из центромеры, хотя в большинстве обстоятельств эти плечи, как таковые, не видны. К тому же большинство эукариотов обладают одним круглым митохондриальным геномом, а некоторые эукариоты могут иметь дополнительные маленькие круглые или линейные цитоплазматические хромосомы.
В ядерных хромосомах эукариотов неуплотненная ДНК существует в полуупорядоченной структуре, где она завернута вокруг гистонов (структурные белки), формируя композитный материал под названием хроматин.
Хроматин
Хроматин – это комплекс ДНК и белка, содержащийся в ядре эукариота, который упаковывает хромосомы. Структура хроматина варьируется значительно между различными этапами клеточного цикла, в соответствии с требованиями ДНК.
Межфазный хроматин
Во время межфазы (период клеточного цикла, когда клетка не делится) можно различить два вида хроматина:
- Эухроматин, который состоит из активной ДНК, то есть выраженной в качестве белка.
- Гетерохроматин, который состоит по большей части из неактивной ДНК. Как кажется, он служит структурным целям во время хромосомных стадий. Гетерохроматин можно далее разделить на два типа:
- Конститутивный гетерохроматин, никогда не выражаемый. Он расположен вокруг центромеры и обычно содержит повторные последовательности.
- Факультативный гетерохроматин, иногда выражаемый.
Метафазный хроматин и деление
На ранних стадиях митоза или мейоза (деление клетки) пряди хроматина становятся все более уплотненными. Они перестают функционировать, как доступный генетический материал (останавливается транскрипция), и становятся компактной транспортабельной формой. Эта компактная форма делает индивидуальные хромосомы видимыми, и они образуют классическую структуру с четырьмя плечами, с парой сестринских хроматид, присоединенных друг к другу в центромере. Более короткие плечи называются «p плечи» (от французского слова «petit» — маленький), а более длинные плечи называются «q плечи» (буква «q» следует за буквой «p» в латинском алфавите; q-g «grande» — большой). Это единственный натуральный контекст, в котором отдельные хромосомы видны при помощи оптического микроскопа.
Во время митоза микротрубочки вырастают из центросом, расположенных на противоположных концах клетки, и также присоединяются к центромере в специализированных структурах под названием кинетохоры, одна из которых присутствует на каждой сестринской хроматиде. Специальная последовательность оснований ДНК в области кинетохоров обеспечивает вместе со специальными белками долговременное присоединение к этой области. Микротрубочки затем оттягивают хроматиды к центросомам, чтобы каждая дочерняя клетка наследовала один набор хроматид. Когда клетки разделились, хроматиды раскручиваются, и ДНК может снова транскрибироваться. Несмотря на свой внешний вид, хромосомы структурно сильно уплотненные, что позволяет этим гигантским ДНК структурам помещаться в клеточные ядра.
Человеческие хромосомы
Хромосомы у людей могут быть разделены на два типа: аутосомы и половые хромосомы. Определенные генетические черты связаны с полом человека и передаются через половые хромосомы. Аутосомы содержат оставшуюся часть генетической наследуемой информации. Все действуют тем же образом во время деления клеток. В человеческих клетках содержатся 23 пары хромосом (22 пары аутосом и одну пару половых хромосом), что дает в целом 46 на клетку. В добавление к ним в человеческих клетках имеется много сотен копий митохондриального генома. Задание последовательности человеческого генома обеспечило много информации о каждой хромосоме. Ниже приводится таблица, в которой собрана статистика для хромосом на основе информации о геноме человека Института Сенгера в базе данных VEGA (Комментарии к геному позвоночных). Число генов — это приблизительная оценка, так как она частично основана на предсказании генов. Общая длина хромосом – это тоже приблизительная оценка, основанная на оцененном размере областей непоследовательных гетерохроматинов.
|
Хромосомы |
Гены |
Общее число комплементарных пар оснований нуклеиновых кислот |
Упорядоченные комплементарные пары оснований нуклеиновых кислот |
|
1 |
422 |
247199719 |
224999719 |
|
2 |
1491 |
242751149 |
237712649 |
|
3 |
155 |
199446827 |
194704827 |
|
4 |
446 |
191263063 |
187297063 |
|
5 |
609 |
180837866 |
177702766 |
|
6 |
2281 |
170896993 |
167273993 |
|
7 |
2135 |
158821424 |
154952424 |
|
8 |
1106 |
146274826 |
142612826 |
|
9 |
192 |
140442298 |
120312298 |
|
10 |
1793 |
135374737 |
131624737 |
|
11 |
379 |
134452384 |
131130853 |
|
12 |
143 |
132289534 |
130303534 |
|
13 |
924 |
114127980 |
95559980 |
|
14 |
1347 |
106360585 |
88290585 |
|
15 |
921 |
100338915 |
81341915 |
|
16 |
909 |
88822254 |
78884754 |
|
17 |
1672 |
78654742 |
77800220 |
|
18 |
519 |
76117153 |
74656155 |
|
19 |
1555 |
63806651 |
55785651 |
|
20 |
1008 |
62435965 |
59505254 |
|
21 |
578 |
46944323 |
34171998 |
|
22 |
1092 |
49528953 |
34893953 |
|
X (половая хромосома) |
1846 |
154913754 |
151058754 |
|
Y (половая хромосома) |
454 |
57741652 |
25121652 |
|
Итого |
32185 |
3079843747 |
2857698560 |
Число хромосом в различных организмах
Эукариоты
В этих таблицах дается общее число хромосом (включая половые) в клеточных ядрах. Например, диплоидные человеческие клетки содержат 22 разных вида аутосомов, каждый присутствует в двух копиях, и две половых хромосомы. Это дает 46 хромосом в целом. Другие организмы имеют более двух копий своих хромосом, например, гексаплоидная хлебная пшеница содержит шесть копий семи разных хромосом, всего 42 хромосомы.
|
Число хромосом в некоторых растениях |
|
||||
|
Виды растений |
№ |
|
|||
|
Arabidopsis thaliana (диплоид) |
10 |
|
|||
|
Рожь (диплоид) |
14 |
|
|||
|
Маис (диплоид или палеотетраплоид) |
20 |
|
|||
|
Пшеница айнкорн (диплоид) |
14 |
|
|||
|
Твердая пшеница (тетраплоид) |
28 |
|
|||
|
Хлебная пшеница (гексаплоид) |
42 |
|
|||
|
Культивированный табак (тетраплоид) |
48 |
|
|||
|
Ужовник погремушковый (диплоид) |
около 1200 |
|
|||
|
|
|||||
|
Число хромосом (2n) в некоторых животных |
|
||||
|
Виды |
№ |
|
|||
|
Дрозофила фруктовая |
8 |
|
|||
|
Гуппи (poecilia reticulata) |
46 |
|
|||
|
Земляной червь (Octodrilus complanatus) |
36 |
|
|||
|
Домашняя кошка |
38 |
|
|||
|
Лабораторная мышь |
40 |
|
|||
|
Кролик (Oryctolagus cuniculus) |
44 |
|
|||
|
Заяц |
48 |
|
|||
|
Гориллы, шимпанзе |
48 |
|
|||
|
Слоны |
56 |
|
|||
|
Осел |
62 |
|
|||
|
Собака |
78 |
|
|||
|
Золотая рыбка |
100-104 |
|
|||
|
Морская свинка |
64 |
|
|||
|
Садовая улитка |
54 |
|
|||
|
Тибетская лиса |
36 |
|
|||
|
Домашняя свинья |
38 |
|
|||
|
Лабораторная крыса |
42 |
|
|||
|
Сирийский хомяк |
44 |
|
|||
|
Человек |
46 |
|
|||
|
Домашняя овца |
54 |
|
|||
|
Корова |
60 |
|
|||
|
Лошадь |
64 |
|
|||
|
Зимородок |
132 |
|
|||
|
Шелкопряд |
56 |
|
|||
|
|
|||||
|
Число хромосом в других организмах |
|||||
|
Виды |
Большие хромосомы |
Промежуточные хромосомы |
Микрохромосомы |
||
|
Trypanosoma brucei |
11 |
6 |
~100 |
||
|
Домашний голубь (Columba livia domestics) |
18 |
— |
59-63 |
||
|
Курица |
8 |
2 половых хромосомы |
60 |
||
Нормальные члены отдельных видов эукариотов имеют то же число ядерных хромосом (см. таблицу). Другие хромосомы эукариотов, то есть митохондриальные и похожие на плазмиды маленькие хромосомы, значительнее варьируются в количестве, и на каждую клетку может быть тысяча копий.
Виды с бесполовым воспроизведением имеют один набор хромосом, тех же самых, что в клетках организма. Однако бесполые виды могут быть гаплоидными и диплоидными.
Виды с половым воспроизведением имеют соматические клетки (клетки организма), которые являются диплоидными [2n], имеющими два набора хромосом, один от матери и другой от отца. Гаметы, репродуктивные клетки, являются гаплоидными [n]: у них один набор хромосом. Гаметы получены мейозом диплоидной клетки зародышевой линии. Во время мейоза соответствующие хромосомы отца и матери могут обмениваться маленькими частями друг друга (скрещивание), и тем самым образуют новые хромосомы, которые не унаследованы только от того или другого родителя. Когда соединяются мужская и женская гаметы (оплодотворение), формируется новый диплоидный организм.
Некоторые виды животных и растений полиплоидные [Xn]: в них есть более двух наборов гомологических хромосом. Важные для сельского хозяйства растения, такие как табак или пшеница, часто полиплоидные, по сравнению с наследственными видами. Пшеница имеет гаплоидное число семи хромосом, обнаруженное в некоторых культурных растениях, а также в диких предках. Более распространенные макаронная и хлебная пшеница – полиплоидные, имеющие 28 (тетраплоид) и 42 (гексаплоид) хромосомы, по сравнению с 14 (диплоид) хромосомами в дикой пшенице.
Прокариоты
Виды прокариотов в целом имеют одну копию каждой главной хромосомы, но большинство клеток может легко выжить с многочисленными копиями. Например, Buchnera, симбионт тли, имеет много копий своей хромосомы, количество которых колеблется от 10 до 400 копий на клетку. Однако в некоторых больших бактериях, таких как Epulopiscium fishelsoni, могут присутствовать до 100 000 копий хромосомы. Количество копий плазмидов и похожих на плазмиды маленьких хромосом, как в эукариотах, значительно колеблется. Число плазмидов в клетке почти полностью определяется скоростью деления плазмидов – быстрое деление порождает высокое число копий.
Кариотип
В целом кариотип – это характерное хромосомное дополнение эукариотических видов. Подготовка и изучение кариотипов – это часть цитогенетики.
Хотя дублирование и транскрипция ДНК высоко стандартизированы в эукариотах, то же самое нельзя сказать для их кариотипов, которые обычно весьма изменчивы. Виды числа хромосом и их детальная организация могут варьироваться. В некоторых случаях между видами может быть значительное колебание. Часто имеется:
- колебание между двумя полами;
- колебание между зародышевой линией и сомой (между гаметами и оставшейся частью организма);
- колебание между членами популяции из-за сбалансированного генетического полиморфизма;
- географическое колебание между расами;
- мозаика или иные аномалии
Также колебание в кариотипе может возникнуть в ходе развития из оплодотворенной яйцеклетки.
Техника определения кариотипа обычно называется кариотипированием. Клетки могут быть блокированы частично через деление (в метафазе) в искусственных условиях (в реакционной пробирке) колхицином. Эти клетки затем окрашиваются, фотографируются и упорядочиваются в кариограмму, с набором упорядоченных хромосом, аутосом в порядке длины и половых хромосом (здесь X/Y) в конце.
Как и во многих видах с половым воспроизведением, у человека имеются специальные гоносомы (половые хромосомы, в противоположность аутосомам). Это XX у женщин и XY у мужчин.
Историческое примечание
На исследование человеческого кариотипа ушло много лет, прежде чем был получен ответ на самый основной вопрос: Сколько хромосом содержится в нормальной диплоидной человеческой клетке? В 1912 г. Ганс вон Винивартер сообщил о 47 хромосомах в сперматогониях и 48 – в оогониях, включая механизм определения пола XX/XO. Пейнтер в 1922 г. не был уверен по поводу диплоидного числа человека – 46 или 48, вначале склоняясь к 46. Он пересмотрел позднее свое мнение с 46 на 48, и правильно настаивал на том, что человек обладает системой XX/XY.
Для окончательного решения проблемы нужны были новые техники:
- Использование клеток в культуре;
- Подготовка клеток в гипотоническом растворе, где они набухают и распространяют хромосомы;
- Задержка митоза в метафазе раствором колхицина;
- Раздавливание препарата на предметодержателе, стимулируя хромосомы в единой плоскости;
- Разрезание микрофотографии и упорядочение результатов в неопровержимой кариограмме.
Только в 1954 г. было подтверждено диплоидное число человека – 46. Учитывая техники Винивартера и Пейнтера, их результаты были довольно примечательными. Шимпанзе (ближайший живущий родственник современных людей) имеет 48 хромосом.
Источник: www.nazdor.ru