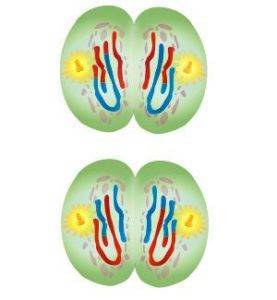
Мейотическое деление клетки
Что такое мейоз?
Редукционное деление клетки, другими словами – мейоз – это вид деления ядра, при котором число хромосом уменьшается в два раза.
Данный процесс происходит в два этапа:
- Редукционный;
На этом этапе в процессе мейоза число хромосом в клетке уменьшается вдвое.
- Эквационный;
В ходе второго деления гаплоидность клеток сохраняется.
Особенностью данного процесса является то, что протекает он только лишь в диплоидных, а также в чётных полиплоидных клетках. А всё потому, что в результате первого деления в профазе 1 в нечётных полиплоидах нет возможности обеспечить попарное слияние хромосом.
Фазы мейоза
В биологии деление происходит на протяжении четырёх фаз: профазы, метафазы, анафазы и телофазы. Мейоз не является исключением, особенностью данного процесса является то, что происходит он в два этапа, между которыми имеется короткая интерфаза.
Первое деление:
Профаза 1 является достаточно сложным этапом всего процесса в целом, состоит она из пяти стадий, которые внесены в следующую таблицу:
Заканчивается профаза образованием веретена деления, разрушением ядерных мембран и самого ядрышка.
Метофаза первого деления знаменательна тем, что хромосомы выстраиваются вдоль экваториальной части веретена деления.
Во время анафазы 1 сокращаются микротрубочки, биваленты разделяются и хромосомы расходятся к разным полюсам.
На этапе телофазы деспирализуются хромосомы и образуется новая ядерная оболочка.

Рис. 1. Схема мейоза первого этапа деления
Второе деление имеет такие признаки:
- Для профазы 2 характерна конденсация хромосом и разделение клеточного центра, продукты деления которого расходятся к противоположным полюсам ядра. Ядерная оболочка разрушается, образуется новое веретено деления, которое располагается перпендикулярно по отношению к первому веретену.
- В ходе метафазы хромосомы вновь располагаются на экваторе веретена.
- Во время анафазы хромосомы делятся и хроматиды располагаются по разным полюсам.
- Телофаза обозначена деспирализацией хромосом и появлением новой ядерной оболочки.
Рис. 2. Схема мейоза второго этапа деления
В результате из одной диплоидной клетки путём такого деления получаем четыре гаплоидных клетки. Исходя из этого, делаем выводы, что мейоз – это форма митоза, в результате которого из диплоидных клеток половых желёз образуются гаметы.
Значение мейоза
В ходе мейоза на этапе профазы 1 происходит процесс кроссинговера – перекомбинация генетического материала. Помимо этого во время анафазы, как первого, так и второго деления, хромосомы и хроматиды расходятся к разным полюсам в случайном порядке. Это объясняет комбинативную изменчивость исходных клеток.
В природе мейоз имеет огромное значение, а именно:
- Это один из основных этапов гаметогенеза;
Рис. 3. Схема гаметогенеза
- Осуществляет передачу генетического кода при размножении;
- Получаемые дочерние клетки не похожи на материнскую клетку, а также различаются между собой.
Мейоз очень важен для образования половых клеток, так как в результате оплодотворения гамет ядра сливаются. В противном случае в зиготе число хромосом было бы вдвое больше. Благодаря такому делению половые клетки гаплоидны, а при оплодотворении восстанавливается диплоидность хромосом.
Источник: obrazovaka.ru
Половое размножение — это возможность многократного увеличения потомства и передачи ему необходимых генетических признаков. Данные факторы с самого начала эволюции способствовали естественному отбору и устанавливали скорость изменений организмов.
Передача генетических признаков у растений, животных и людей происходит через половые клетки — яйцеклетки и сперматозоиды. Если бы половые клетки имели полный набор генетических характеристик, то после их слияния зарождался бы организм с удвоенным набором.
Известно, что число хромосом строго определено. Если происходит изменение в заданном наборе, то это приводит к развитию тяжелых патологий у плода или же к его гибели. Механизм, который в два раза уменьшает количество хромосом — мейотическое деление клеток (мейоз).
Мейоз происходит методом двух делений в последовательном порядке. Редукционное деление (первое) приводит к удвоенному уменьшению количества хромосом. Второе деление направлено на уравнивание хромосом (эквационное деление), в целом повторяющее митоз. Мейоз состоит из ряда фаз, в ходе которых хромосомы изменяются:
- профаза;
- метафаза;
- анафаза;
- телофаза.
Фазы аналогичны фазам митоза.
Профаза. Происходит увеличение ядра, оболочка фрагментируется. Процесс спирализации приводит к формированию тонких нитей хромосом. Уже по окончанию профазы заметно, что хромосома состоит из двух хроматид. Хромосомы находятся в беспорядочном положении в цитоплазме клетки. Центросомы расходятся к противоположным полюсам.
Метафаза. На данной стадии хромосомы достигают наибольшей спирализации и размещаются на экваторе. В этой фазе можно легко пересчитать количество хромосом и изучить их морфологию.
Анафаза. В этой фазе каждая хромосома распадается на равные хроматиды, расходящиеся к противоположным полюсам. Из-за идентичности хроматид у полюсов клетки создается равнозначный генетический материал.
Телофаза. Фаза, в которой дочерние хромосомы деспирализуются, и ДНК становится возможным транскрибировать. Далее образуется ядерная оболочка вокруг хромосом, а в ядрах — ядрышки. Нити распадаются и на этом процесс кариокинеза заканчивается. Затем начинается цитокинез. Следовательно, в процессе митоза образуются две дочерние клетки с аналогичным хромосомным набором.
Биологическое значение мейоза
Мейотическое деление клеток обеспечивает передачу генетического материала потомкам. Мейоз — это один из самых важных этапов в половом размножении. В процессе происходит уменьшение хромосом от числа 46 до 23 (у человека).
Комбинативная изменчивость, которая обеспечивает генетическую уникальность человека, достигается мейотическим делением.
Мейотическое деление называют делением созревания, так как формирование гамет напрямую связано с редукцией хромосом.
Значение митоза в биологических процессах
Митоз — это непрямое деление клеток, способствующее зарождению двух дочерних клеток с равным набором хромосом из одной соматической. Митоз длится от 2 до 3 часов у растительных клеток, а у животных — от 30 до 60 минут.
Роль митоза заключается в создании клеток с одинаковым генетическим материалом. Достигается это методом репликации, когда ДНК удваиваются и воспроизводятся две равные хроматиды, впоследствии распределяющиеся между клетками. Митоз является необходимым для адекватного развития многоклеточного организма.
Амитоз
Амитоз — это прямое деление ядра методом перетяжки. Встречается у одноклеточных, например, у полиплоидных ядер инфузорий. Кроме того, таким способом делятся некоторые физически ослабленные, постепенно отмирающие клетки животных или растений. Также амитоз характерен для разных злокачественных процессов, воспалений и т.д.
Что касается растений, то амитоз имеется в эндосперме семян, черешках листьев.
Для животных и людей амитоз характерен для клеток роговицы глаза, печени и хрящей.
При таком типе деления часто происходит редукция одного ядра. Возможно возникновение двух- или многоядерной клетки. В случае, когда после деления ядра осуществляется деление цитоплазмы, то распределение ДНК и клеток происходит хаотично.
Гибель клеток
В многоклеточных организмах очень часто наблюдается смерть клеточных групп или клеток в отдельности. Существует две категории гибели клеток: некроз (необратимое отмирание) и апоптоз (фрагментация клетки на отдельные тельца).
Деление клеток и онкологические заболевания
Процесс деления находится под контролем двух генов — протоонкогенов и генов-супрессоров.
Протоонкогены выполняют функцию блокирования белков, которые стимулируют деление клеток. Гены-супрессоры препятствуют делению клеток.
Спровоцировать мутирование клеток способно химическое воздействие канцерогенов, вирусы и лучи. Происходят изменения в ДНК-структуре, способствующие превращению протоонкогенов в активные онкогены. Таким образом, нарушения в процессе деления клеток приводят к образованию опухолей.
Клетки с измененным ДНК имеют свойство саморазрушаться путем запрограммированного отмирания клеток (апоптоза), останавливая рост опухолей. Однако не во всех случаях развитие опухоли прекращается.
Источник: www.probirka.org
Что такое мейоз?
Редукционное деление клетки, другими словами – мейоз – это вид деления ядра, при котором число хромосом уменьшается в два раза.
Данный процесс происходит в два этапа:
- Редукционный;
На этом этапе в процессе мейоза число хромосом в клетке уменьшается вдвое.
- Эквационный;
В ходе второго деления гаплоидность клеток сохраняется.
Особенностью данного процесса является то, что протекает он только лишь в диплоидных, а также в чётных полиплоидных клетках. А всё потому, что в результате первого деления в профазе 1 в нечётных полиплоидах нет возможности обеспечить попарное слияние хромосом.
Фазы мейоза
В биологии деление происходит на протяжении четырёх фаз: профазы, метафазы, анафазы и телофазы. Мейоз не является исключением, особенностью данного процесса является то, что происходит он в два этапа, между которыми имеется короткая интерфаза.
Первое деление:
Профаза 1 является достаточно сложным этапом всего процесса в целом, состоит она из пяти стадий, которые внесены в следующую таблицу:
Заканчивается профаза образованием веретена деления, разрушением ядерных мембран и самого ядрышка.
Метофаза первого деления знаменательна тем, что хромосомы выстраиваются вдоль экваториальной части веретена деления.
Во время анафазы 1 сокращаются микротрубочки, биваленты разделяются и хромосомы расходятся к разным полюсам.
На этапе телофазы деспирализуются хромосомы и образуется новая ядерная оболочка.
Рис. 1. Схема мейоза первого этапа деления
Второе деление имеет такие признаки:
- Для профазы 2 характерна конденсация хромосом и разделение клеточного центра, продукты деления которого расходятся к противоположным полюсам ядра. Ядерная оболочка разрушается, образуется новое веретено деления, которое располагается перпендикулярно по отношению к первому веретену.
- В ходе метафазы хромосомы вновь располагаются на экваторе веретена.
- Во время анафазы хромосомы делятся и хроматиды располагаются по разным полюсам.
- Телофаза обозначена деспирализацией хромосом и появлением новой ядерной оболочки.
Рис. 2. Схема мейоза второго этапа деления
В результате из одной диплоидной клетки путём такого деления получаем четыре гаплоидных клетки. Исходя из этого, делаем выводы, что мейоз – это форма митоза, в результате которого из диплоидных клеток половых желёз образуются гаметы.
Значение мейоза
В ходе мейоза на этапе профазы 1 происходит процесс кроссинговера – перекомбинация генетического материала. Помимо этого во время анафазы, как первого, так и второго деления, хромосомы и хроматиды расходятся к разным полюсам в случайном порядке. Это объясняет комбинативную изменчивость исходных клеток.
В природе мейоз имеет огромное значение, а именно:
- Это один из основных этапов гаметогенеза;
Рис. 3. Схема гаметогенеза
- Осуществляет передачу генетического кода при размножении;
- Получаемые дочерние клетки не похожи на материнскую клетку, а также различаются между собой.
Мейоз очень важен для образования половых клеток, так как в результате оплодотворения гамет ядра сливаются. В противном случае в зиготе число хромосом было бы вдвое больше. Благодаря такому делению половые клетки гаплоидны, а при оплодотворении восстанавливается диплоидность хромосом.
Источник: obrazovaka.ru
При половом размножении сходство потомков с родителями обеспечивается через половые клетки. Несмотря на свои ничтожные по сравнению с телом организма размеры, они несут в себе всю наследственную информацию, предопределяющую ход развития будущего организма.
Основой полового размножения является оплодотворение (сингамия), то есть слияние двух половых клеток.
Физиологическая специализация половых клеток обусловила особенности их морфологии и физиологии, из-за чего женские и мужские половые клетки значительно отличаются друг от друга. Яйцеклетка женского организма не
только отвечает за передачу наследственной информации потомству, но и за питание эмбриона на ранних стадиях его развития. Мужская половая клетка, сперматозоид, этой функцией не обладает, а обеспечивает передачу наследственных свойств отцовского организма потомкам и стимулирует яйцеклетку к развитию.
В основе развития половых клеток лежит мейоз, или редукционное деление клеток. В него вступают незрелые половые клетки, достигшие определенной дифференциации. То есть он совершается в период образования гамет, или гаметогенеза. Если бы каждая половая клетка имела диплоидный набор хромосом, заключающийся в соматической клетке, то число хромосом удваивалось бы в каждой последующей генерации. А так как кариотип у каждого вида животных постоянен, то число хромосом в гаметах должно быть
гаплоидным. За уменьшение числа хромосом в гаметах (их редукцию) и отвечает мейоз.
Сущность мейоза заключается в том, что происходит два быстро следующих друг за другом деления ядра, а хромосомы при этом редуплицируются только один раз. Первое деление называется редукционным. При этом происходит уменьшение числа хромосом в ядрах вдвое за счет сближения гомологичных хромосом, называемого конъюгацией. При этом они, как правило, взаимно перекручиваются и состоят из двух соединенных центромерой хроматид. Во время конъюгации гомологичные хромосомы в местах перекручивания могут разрываться и обмениваться между собой гомологичными участками. В этот процесс, называемый кроссинговером, вовлекаются только две (любыехроматиды из четырех — по одной из каждой гомологичной хромосомы (см. рис. 3). Кроссинговер приводит к перекомбинации генетического материала в хромосомах, а это означает, что в гаметах будут не только хромосомы, идентичные родительским (не обменявшиеся участками), но и хромосомы с новой комбинацией материала двух гомологичных хромосом. Из каждой пары гомологичных хромосом материнской клетки в дочерние ядра попадает по одной, то есть гомологичные хромосомы, в отличие от негомологичных, которые комбинируются по-разному, зависят друг от друга. Второе деление — обычное и называется эквационным, или уравнительным. При этом центромера каждой хромосомы делится и дочерние хромосомы (бывшие хроматиды) в равном, но уменьшенном вдвое количестве расходятся кпротивоположным полюсам клетки. В результате двух делений мейоза из одной диплоидной родительской клетки образуется четыре гаплоидных дочерних клетки, содержащие по 39 хромосом (по одной из пары гомологичных хромосом). После образования зиготы в ней окажется 78 хромосом из 39 гомологичных пар.
Если бы каждая половая клетка имела диплоидный набор хромосом, заключающийся в соматической клетке, то число хромосом удваивалось бы в каждой последующей генерации. А так как кариотип у каждого вида животных постоянен, то число хромосом в гаметах должно быть гаплоидным. За уменьшение хромосом в гаметах (их редукцию) и отвечает мейоз.
Сущность мейоза заключается в том, что происходит два быстро следующих друг за другом деления ядра, а хромосомы при этом редуплицируются только один раз. Первое деление называется редукционным. При этом происходит уменьшение числа хромосом в ядрах вдвое за счет сближения гомологичных хромосом, называемого конъюгацией. При этом они, как правило, взаимно перекручиваются и состоят из двух соединенных центромерой хроматид. Во время конъюгации гомологичные хромосомы в местах перекручивания могут разрываться и обмениваться между собой гомологичными участками. В этот процесс, называемый кроссинговером, вовлекаются только две (любые) хроматиды из четырех — по одной из каждой гомологичной хромосомы (см. рис. 3). Кроссинговер приводит к перекомбинации генетического материала в хромосомах, а это означает, что в гаметах будут не только хромосомы, идентичные родительским (не обменявшиеся участками), но и хромосомы с новой комбинацией материала двух гомологичных хромосом. Из каждой пары гомологичных хромосом материнской клетки в дочерние ядра попадает по одной, то есть гомологичные хромосомы, в отличие от негомологичных, которые комбинируются по-разному, зависят друг от друга. Второе деление — обычное и называется эквационным, или уравнительным. При этом центромера каждой хромосомы делится и дочерние хромосомы (бывшие хроматиды) в равном, но уменьшенном вдвое количестве расходятся к противоположным полюсам клетки. В результате двух делений мейоза из одной диплоидной родительской клетки образуется четыре гаплоидных дочерних клетки, содержащие по 39 хромосом (по одной из пары гомологичных хромосом). После образования зиготы в ней окажется 78 хромосом из 39 гомологичных пар.
Теоретически в каждую гамету попадает одинаковое количество хромосом матери и отца. Но не следует забывать о независимом поведении негомологичных хромосом и кроссинговере гомологичных хромосом во время мейоза, что сопровождается рекомбинацией генов и приводит к появлению мириад гамет, отличающихся от материнской клетки и между собой по набору хромосом, не говоря уже о том, что любая из гамет может участвовать в образовании зиготы.
Способы, которые позволяли бы нам контролировать при разведении собак случайное распределение хромосом и находящихся в них генов, неизвестны, что делает генетические прогнозы крайне сложными и увлекательными. Но из этого вовсе не следует, что мы должны быть фаталистами и считать плоды своей племенной работы чистой случайностью. Конечно, определенный элемент случайности при этом имеется.
Индивидуальность каждой хромосомы в кариотипе (аутосом и половых «X» и «Y») обусловлена не только их формой и размером, но и набором генов. Участок ДНК, в котором расположен ген, обусловливающий какой-то признак, называют локусом. Например, локус пигментации шерсти, локус группы крови и др. Гены одного локуса обозначают прописными или строчными латинскими буквами. Так, у собаки ген черной окраски обозначается буквой «В», ген короткошерстности — «d», ген крипторхизма «с».
Гены различных признаков расположены в хромосоме линейно. Поэтому иногда наблюдается совместное наследование признаков, гены которых расположены в данной хромосоме. Такое наследование называют «сцепленным».
У собак установлено сцепленное наследование признаков, гены которых присутствуют в Х-хромосоме, а именно: крипторхизм (ген с) и болезнь крови — гемофилия (ген h).
Для некоторых пород собак уже частично составлены карты хромосом, которые указывают, в каком участке той или иной хромосомы расположен локус, несущий ген данного признака.
По своему основному действию гены могут быть доминантными (обозначаются заглавными буквами А, В, С, D) и рецессивными (обозначаются прописными буквами а, в, с, d). Каждый ген из пары гомологических хромосом данного локуса получает символ двух букв и это соответствует генотипу данного локуса данного признака.
Например, генотип собаки по локусу черной пигментации шерсти будет записан в виде двух букв ВВ, если аллели отца и матери по этому локусу доминантны. Генотип для двух признаков будет записан четырьмя буквами. Например, если собака имеет крипторхизм (ген с) и черную окраску шерсти (ген В), то генотип по этим признакам записывается сс ВВ.
Сочетание аллелей в локусе и образованные ими генотипы могут быть; гомозиготный доминантный (ВВ), гомозиготный рецессивный (вв), гетерозиготный (неоднородный) (Вв). Следовательно, генотип — это совокупность наследственных задатков генов. Он может быть гомозиготным или гетерозиготным, а фенотип — это комплекс реализованных наследственных задатков в определённых качествах и количествах признака. Некоторые условия среды могут создавать оптимальную возможность реализации генотипа, а другие тормозят реализацию наследственности.
Гены отдельных локусов могут иметь не два аллельных состояния, а несколько. Это вызывается многократным мутированием исходного доминантного гена или множественным аллелизмом. В результате создается серия рецессивных аллелей, что увеличивает наследственное разнообразие признаков. Серии множественных аллелей часто наблюдаются с генами, обусловливающими синтез пигмента шерсти у собак. Каждый новый аллель такой серии вызывает синтез нового пигмента, обусловливающего возникновение новой окраски шерсти, где последующий рецессивен к предыдущему.
По данным Робертсона серия множественных аллелей такого типа была давно использована в селекции многих пород собак и привела к большому разнообразию мастей у собак разных пород.
Известна следующая серия окрасок: сплошная черная (ген As), доминантная желтая (ген AY), зонарная пигментация (ген агути А), чепрачная (ген aSA), кофейная сплошная (гена).
Взаимоотношение между аллелями этой серии таково, что от доминантного исходного гена А остальные аллели серии сопровождаются ослаблением интенсивности признака окраски и составляют такой ряд: As>AY> А >aSA> а1.
Под влиянием мутагенных факторов (радиации, ультразвука, химических веществ) происходит изменение структуры гена, его азотистых оснований молекулы ДНК — это толковые (или генные) мутации. Воздействие любого мутагенного фактора может вызывать перестройку участков хромосом или обмен участками разных хромосом друг с другом и даже может увеличивать их число в кариотипе (полиплоидия). Такие изменения называют хромосомными мутациями в отличие от генных мутаций, при которых изменяется только структуры ДНК. В результате мутационных изменений в строении гена (ДНК) или хромосом происходит изменение и появление новых свойств, признаков.
Онтогенетические мутации, происходящие в соматических клетках, могут вызывать онкологические перестройки в таких клетках и в тканях. Если мутационный процесс происходит в гаметах родителей, то это приводит к появлению у их потомков новых признаков и свойств, часто имеющих и патологические свойства с проявлением аномалий, нарушением обмена веществ: анемия, гемофилия, анкилоз и др.
Мутационная изменчивость служит важным источником создания новых признаков, которые могут закрепляться в ряде поколений при селекции, управляемой кинологом.
У собак, например, некоторые мутационные признаки закреплены путем ортоселекции и стали породным признаком (мопсовидность, коротконогость, короткошерстностьи др.)
1.3 Закономерности наследования признаков
Закономерности наследования признаков родителей их потомством выявлены путем проведения скрещивания родительских пар. Начало этих работ заложено еще Т. Менделем (1862 г.).
Закономерности наследования генов у собак различаются внутриаллельным (в пределах локуса) и их межаллельным взаимодействиям. Если скрещивать самца и самку, различающихся по какому-то одному признаку, то можно установить, какой из признаков имеет доминантный тип, то есть проявляется у потомка первого поколения, а какой остается в скрытом состоянии, так как обусловлен рецессивным аллелем. При этом в первом поколении все потомки имеют фенотип с доминантным признаком, а их генотип гетерозиготный по обоим аллелям. Bэтом проявляется первый закон Менделя: единообразие потомства первого поколения (Ft) по доминантному признаку. При скрещивании животных этого поколения между собой во втором поколении (F2) появляются животные двух фенотипов: 75 % животных с доминантным состоянием признака и 25 % — с рецессивным, т. е. в соотношении 3:1.
Это второй закон Менделя — закон «расщепления» признаков у потомства F2 на доминантные и рецессивные фенотипы. Например, при вязке короткошерстной собаки (LL) с длинношерстной собакой(11) их потомство (FAбудет иметь гетерозиготный генотип (L1), а фенотип — короткошерстный.
Если будем вязать гетерозиготных собак между собой, то у их потомства во втором поколении (F2), 75 процентов собак будет короткошерстными, а 25 процентов длинношерстными. По генотипу расщепление выразится: 25 % LL, 50 %L1 , 25 % 11 , что дает соотношение 1:2:1.
Если при вязке учитывать не один признак (моногибридное скрещивание), то наследование будет сопровождаться увеличением разнообразия фенотипов и генотипов, во втором поколении в результате большого комбинирования у потомства исходных родительских признаков. Примером этого может служить разнопородное скрещивание коричневого (вв) короткошерстного (LL) добермана (генотип LLbb) и длинношерстного (11) с черной окраской (ВВ) ньюфаундленда (генотип 11ВВ).
Случается и такое взаимодействие аллелей одного и того же локуса, когда в признаке проявляется одновременно действие обоих аллельных генов локуса. Этот тип наследования назван кодоминированием генов. Он часто проявляется в генотипах, обусловливающих синтез разных белков. Так, например, в организме собаки в результате кодоминантного действия аллельных генов А и В локуса гемоглобина в эритроцитах образуется три типа гемоглобина с генотипами АА, ВВ и АВ. Это нормальные гемоглобины, но несколько отличаются биохимически, что полезно для жизни животного.
Взаимодействие аллелей генов разных локусов приводит к появлению нового признака у потомства, которого не было у родителей. Этот тип наследования называется «новообразованием при скрещивании». Например, при вязке коричневой суки с голубым кобелем, потомство у доберманов будет иметь черную окраску шерсти.
При комплементарном типе взаимодействия генов, расположенных в разных участках хромосом, взаимодействуют два доминантных гена разных локусов, причем каждый из них не дает фенотипического проявления, а совместное комплементарное их расположение приводит к формированию нового признака, которого не было ни у суки, ни у кобеля.
Например, комплементарное воздействие генов у собак выявлено в виде паралича задних конечностей у помесного потомства, полученного от скрещивания здоровых родителей немецкого дога с сенбернаром. Заболевание проявляется в разной степени: от слабой парализованности до полной неподвижности.
В наследовании некоторых генов наблюдается действие «генов-модификаторов», которые обусловливают разную степень проявления признаков. Например, степень пятнистости окраса шерсти собак различается от сплошной черной, через серию большей или меньшей пятнистости и почти до полностью белой окраски шерсти по всему телу (доги, овчарки, колли, фокстерьеры и другие породы).
Существенное значение в наследовании имеет так называемое плейотропное (множественное) действие гена, когда один и тот же ген влияет на образование разных признаков. У собак описан ген (N), имеющий плейотропное действие. Он вызывает бесшерстность. Известны также гены дефектов и недоразвития зубной системы, у борзых — генов белой окраски шерсти и глухоты. У собак породы дункер описаны полулетальные гены «крапчатости», вызывающие крапчатость окраски шерсти, уменьшение размера глазного яблока, дефект радужной оболочки (Коломбо), глаукому с выпячиванием глазного яблока и далее слепоту. Плейотропное действие гена может вызвать голубую окраску радужной оболочки, глухоту, общую слабость, пониженную функцию размножения.
Особый тип наследования наблюдается при взаимодействии между несколькими доминантными генами разных локусов в виде так называемого эпистаза. В этом случае гены, образующие эпистатическую серию, характерны тем, что каждый последующий ген как бы подавляется доминантным геном, занимающим предыдущее место в эпистатической серии, и, в свою очередь, он доминантен по отношению к последующему.
Это хорошо прослежено в наследовании окраса у короткошерстных собак.
Существует так называемое полимерное (полигенное) воздействие генов разных локусов на один и тот же признак. Каждый из этих генов усиливает развитие признака, поэтому степень проявления признака зависит от количества доминантных генов разных локусов. Полигенное действие генов обусловливает наследственность количественных признаков (размер и живая масса тела, плодовитость, скорость бега у собак и т. п.).
Из приведенного перечня действия и взаимодействия генов (внутриаллельное: доминирование, рецессивность, кодоминирование, сверхдоминирование, модифицирующее действие; межаллельное: новообразования, комплементарность, полигения, плейотропия) ясно, что проявление действия генов многообразно и сложно. Передача генов от родителей потомкам и наследственное формирование признаков является сложным процессом, механизм которого заложен в молекулярной структуре ДНК. Воздействие некоторых внешних факторов может вызвать наследственную изменчивость, т. е. генное или хромосомное.
Источник: poznayka.org
Интерфаза
- Фаза G1: этап развития клетки перед синтезом ДНК. На этой стадии клетка подготавливаясь к делению увеличивается в массе.
- S-фаза: период, в течение которого синтезируется ДНК. Для большинства клеток эта фаза занимает короткий промежуток времени.
- Фаза G2: период после синтеза ДНК, но до начала профазы. Клетка продолжает синтезировать дополнительные белки и увеличиваться в размерах.
В последней фазе интерфазы клетка все еще имеет нуклеолы. Ядро окружено ядерной мембраной, а клеточное хромосомы дублируются, но находятся в форме хроматина. В клетках животных две пары центриолей, образованных из репликации одной пары, расположены за пределами ядра. В конце интерфазы клетка переходит в первый этап мейоза.
Читайте также: Основные сходства и различия между митозом от мейозом.
Мейоз I:
Профаза I
В профазе I мейоза происходят следующие изменения:
- Хромосомы конденсируются и присоединяются к ядерной оболочке.
- Возникает синапсис (попарное сближение гомологичных хромосом) и образуется тетрада. Каждая тетрада состоит из четырех хроматид.
- Может произойти генетическая рекомбинация.
- Хромосомы сгущаются и отсоединяются от ядерной оболочки.
- Подобно митозу, центриоли мигрируют друг от друга, а ядерная оболочка и ядрышки разрушаются.
- Хромосомы начинают миграцию к метафазной (экваториальной) пластине.
В конце профазы I клетка входит в метафазу I.
Метафаза I
В метафазе I мейоза происходят следующие изменения:
- Тетрады выравниваются на метафазной пластине.
- Центромеры гомологичных хромосом ориентированы на противоположные полюса клетки.
В конце метафазы I клетка входит в анафазу I.
Анафаза I
В анафазе I мейоза происходят происходят следующие изменения:
- Хромосомы перемещаются в противоположные концы клетки. Подобно митозу, кинетохоры взаимодействуют с микротрубочками, чтобы переместить хромосомы к полюсам клетки.
- В отличие от митоза, сестринские хроматиды остаются вместе после того, как гомологичные хромосомы перемещаются в противоположные полюса.
В конце анафазы I клетка входит в телофазу I.
Телофаза I
В телофазе I мейоза происходят следующие изменения:
- Волокна веретена продолжают перемещать гомологичные хромосомы на полюса.
- Как только движение завершено, каждый полюс клетки имеет гаплоидное количество хромосом.
- В большинстве случаев цитокинез (деление цитоплазмы) происходит одновременно с телофазой I.
- В конце телофазы I и цитокинеза образуются две дочерние клетки, каждая из которых имеет половину числа хромосом исходной родительской клетки.
- В зависимости от типа клетки могут возникать различные процессы при подготовке к мейозу II. Однако генетический материал не реплицируется снова.
В конце телофазы I клетка входит в профазу II.
Мейоз II:
Профаза II
В профазе II мейоза происходят следующие изменения:
- Ядерная мембрана и ядра разрушаются, пока появляется веретено деления.
- Хромосомы больше не реплицируются в этой фазе.
- Хромосомы начинают мигрировать к метафазной пластинке II (на экваторе клеток).
В конце профазы II клетки входят в метафазу II.
Метафаза II
В метафазе II мейоза происходят следующие изменения:
- Хромосомы выстраиваются на метафазной пластинке II в центре клеток.
- Кинетохорные нити сестринских хроматид расходятся к противоположным полюсам.
В конце метафазы II клетки входят в анафазу II.
Анафаза II
В анафазе II мейоза происходят следующие изменения:
- Сестринские хроматиды разделяются и начинают перемещаться к противоположным концам (полюсам) клетки. Волокна веретена деления, не связанные с хроматидами, вытягиваются и удлиняют клетки.
- Как только парные сестринские хроматиды отделены друг от друга, каждая из них считается полной хромосомой, называемые дочерними хромосомами.
- При подготовке к следующему этапу мейоза два полюса клеток также отдаляются друг от друга во время анафазы II. В конце анафазы II каждый полюс содержит полную компиляцию хромосом.
После анафазы II клетки входят в телофазу II.
Телофаза II
В телофазе II мейоза происходят следующие изменения:
- Образуются отдельные ядра на противоположных полюсах.
- Происходит цитокинез (деление цитоплазмы и образование новых клеток).
- В конце мейоза II производятся четыре дочерние клетки. Каждая клетка имеет половину числа хромосом от исходной родительской клетки.
Результат мейоза
Конечным результатом мейоза является производство четырех дочерних клеток. Эти клетки имеют в двое меньше хромосом относительно родительской. При мейозе продуцируются только половые клетки. Другие типы клеток делятся посредством митоза. Когда половые гаплоидные клетки объединяются во время оплодотворения, они становятся диплоидной клеткой. Диплоидные клетки имеют полный набор гомологичных хромосом.
Понравилась статья? Поделись с друзьями:
Источник: NatWorld.info