Период покоя клетки называется
Биологическое значение деления клеток. Новые клетки возникают в результате деления уже существующих. Если делится одноклеточный организм, то из него образуются два новых. Многоклеточный организм также начинает свое развитие чаще всего с одной-единственной клетки. Путем многократных делений образуется огромное количество клеток, которые и составляют организм. Деление клеток обеспечивает размножение и развитие организмов, а значит, непрерывность жизни на Земле.
Клеточный цикл — жизнь клетки с момента ее образования в процессе деления материнской клетки до собственного деления (включая это деление) или гибели.
В течение этого цикла каждая клетка растет и развивается таким образом, чтобы успешно выполнять свои функции в организме. Далее клетка функционирует определенное время, по истечении которого либо делится, образуя дочерние клетки, либо погибает.
У различных видов организмов клеточный цикл занимает разное время: например, у бактерий он длится около 20 мин, у инфузории туфельки — от 10 до 20 ч.
етки многоклеточных организмов на ранних стадиях развития делятся часто, а затем клеточные циклы значительно удлиняются. Например, сразу после рождения человека клетки головного мозга делятся огромное число раз: 80 % нейронов головного мозга формируется именно в этот период. Однако большинство этих клеток быстро теряет способность к делению, а часть доживает до естественной смерти организма, вообще не делясь.
Клеточный цикл состоит из интерфазы и митоза (рис. 54).

Интерфаза — промежуток клеточного цикла между двумя делениями. В течение всей интерфазы хромосомы неспирализованы, они находятся в ядре клетки в виде хроматина. Как правило, интерфаза состоит из трех периодов: пре-синтетического, синтетического и постсинтетического.
Пресинтетический период (G,) — наиболее продолжительная часть интерфазы. Он может продолжаться у различных типов клеток от 2—Зч до нескольких суток. Во время этого периода клетка растет, в ней увеличивается количество органоидов, накапливается энергия и вещества для последующего удвоения ДНК- В течение Gj-периода каждая хромосома состоит из одной хроматиды, т. е. количество хромосом (п) и хроматид (с) совпадает. Набор хромосом и хро-
матид (молекул ДНК) диплоидной клетки в Grпериоде клеточного цикла можно выразить записью 2п2с.
В синтетическом периоде (S) происходит удвоение ДНК, а также синтез белков, необходимых для последующего формирования хромосом. В этот же период происходит удвоение центриолей.
Удвоение ДНК называют репликацией. В ходе репликации специальные ферменты разъединяют две цепи исходной материнской молекулы ДНК, разрывая водородные связи между комплементарными нуклеотидами. С разошедшимися цепями связываются молекулы ДНК-полимеразы — главного фермента репликации. Затем молекулы ДНК-полимеразы начинают двигаться вдоль материнских цепей, используя их в качестве матриц, и синтезировать новые дочерние цепи, подбирая для них нуклеотиды по принципу комплементарности (рис. 55). Например, если участок материнской цепи ДНК имеет последовательность нуклеотидов А Ц Г Т Г А, то участок дочерней цепи будет иметь вид ТГЦАЦТ. В связи с этим репликацию относят к реакциям матричного синтеза. В результате репликации образуются две идентичные двуцепочечные молекулы ДНК- В состав каждой из них входит одна цепочка исходной материнской молекулы и одна вновь синтезированная дочерняя цепочка.
К концу S-периода каждая хромосома состоит уже из двух идентичных сестринских хроматид, соединенных друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2п4с.
Постсинтетический период (G2) наступает после удвоения ДНК- В это время клетка накапливает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего С2-периода набор хромосом и хроматид в клетке остается неизменным — 2п4с.
Интерфаза завершается, и начинается деление, в результате которого образуются дочерние клетки. В ходе митоза (основного способа деления клеток эукариот) сестринские хроматиды каждой хромосомы отделяются друг от друга и попадают в разные дочерние клетки. Следовательно, молодые дочерние клетки, вступающие в новый клеточный цикл, имеют набор 2п2с.
Таким образом, клеточный цикл охватывает промежуток времени от возникновения клетки до ее полного разделения на две дочерние и включает интерфазу (Gr, S-, С2-периоды) и митоз (см. рис. 54). Такая последовательность периодов клеточного цикла характерна для постоянно делящихся клеток, например для клеток росткового слоя эпидермиса кожи, красного костного мозга, слизистой оболочки желудочно-кишечного тракта животных, клеток образовательной ткани растений. Они способны делиться каждые 12—36 ч.
В противоположность этому большинство клеток многоклеточного организма встают на путь специализации и после прохождения части Gj-периода могут переходить в так называемый период покоя (Go-период). Клетки, пребывающие в Gn-периоде, выполняют свои специфические функции в организме, в них протекают процессы обмена веществ и энергии, но не происходит подготовка к репликации. Такие клетки, как правило, навсегда утрачивают способность к делению. Примерами могут служить нейроны, клетки хрусталика глаза и многие другие.
Однако некоторые клетки, находящиеся в Gn-периоде (например, лейкоциты, клетки печени), могут выходить из него и продолжать клеточный цикл, пройдя все периоды интерфазы и митоз. Так, клетки печени могут снова приобретать способность к делению спустя несколько месяцев пребывания в периоде покоя.
Клеточная гибель. Гибель (смерть) отдельных клеток или их групп постоянно встречается у многоклеточных организмов, так же как гибель одноклеточных организмов. Гибель клеток можно разделить на две категории: некроз (от греч. некрос — мертвый) и ап о птоз, который часто называют программируемой клеточной смертью или даже клеточным самоубийством.
Некроз — отмирание клеток и тканей в живом организме, вызванное действием повреждающих факторов. Причинами некроза может быть воздействие высоких и низких температур, ионизирующих излучений, различных химических веществ (в том числе токсинов, выделяемых болезнетворными микроорганизмами). Некротическая гибель клеток наблюдается также в результате их механического повреждения, нарушения кровоснабжения и иннервации тканей, при аллергических реакциях.
В повре>вденных клетках нарушается проницаемость мембран, останавливается синтез белков, прекращаются другие процессы обмена веществ, происходит разрушение ядра, органоидов и, наконец, всей клетки. Особенностью некроза является то, что такой гибели подвергаются целые группы клеток (например, при инфаркте миокарда из-за прекращения снабжения кислородом отмирает участок сердечной мышцы, содержащий множество клеток). Обычно отмирающие клетки подвергаются атаке лейкоцитов, и в зоне некроза развивается воспалительная реакция.
Апоптоз — запрограммированная гибель клеток, регулируемая организмом. В ходе развития и функционирования организма часть его клеток погибает без непосредственного повреждения. Этот процесс протекает на всех стадиях жизни организма, даже в эмбриональный период.
Во взрослом организме также постоянно происходит запланированная гибель клеток. Миллионами гибнут клетки крови, эпидермиса кожи, слизистой оболочки желудочно-кишечного тракта и др. После овуляции погибает часть фолликулярных клеток яичника, после лактации — клетки молочных желез. В организме взрослого человека ежедневно в результате апоптоза гибнет 50—70 миллиардов клеток. При апоптозе клетка распадается на отдельные фрагменты, окруженные плазмалеммой. Обычно фрагменты погибших клеток поглощаются лейкоцитами или соседними клетками без запуска воспалительной реакции. Восполнение утраченных клеток обеспечивается путем деления.
Таким образом, апоптоз как бы прерывает бесконечность клеточных делений. От своего «рождения» до апоптоза клетки проходят определенное количество нормальных клеточных циклов. После каждого из них клетка переходит либо к новому клеточному циклу, либо к апоптозу.
Источник: botana.biz
Деление клеток
Хромосомный набор
Хромосомный набор — совокупность хромосом, содержащихся в ядре. В зависимости от хромосомного набора клетки бывают соматическими и половыми.
Соматические и половые клетки
| Тип | Хромосомный набор | Характеристика |
| Соматические | 2n | Диплоидны — содержат двойной набор хромосом. В этих клетках хромосомы представлены парами. Хромосомы, принадлежащие к одной паре, называются гомологичными. |
| Половые | 1n | Гаплоидны — содержат одинарный набор хромосом. В этих клетках хромосомы представлены в единственном числе и не имеют пары в виде гомологичной хромосомы. |
Клеточный цикл
Клеточный цикл (жизненный цикл клетки) — существование клетки от момента её возникновения в результате деления материнской клетки до её собственного деления или смерти.
одолжительность клеточного цикла зависит от типа клетки, её функционального состояния и условий среды. Клеточный цикл включает митотический цикл и период покоя.
В период покоя (G0) клетка выполняет свойственные ей функции и избирает дальнейшую судьбу — погибает либо возвращается в митотический цикл. В непрерывно размножающихся клетках клеточный цикл совпадает с митотическим циклом, а период покоя отсутствует.
Митотический цикл состоит из четырёх периодов: пресинтетического (постмитотического) — G1, синтетического — S, постсинтетического (премитотического) — G2, митоза — М. Первые три периода — это подготовка клетки к делению (интерфаза), четвёртый период — само деление (митоз).
Интерфаза — подготовка клетки к делению — состоит из трёх периодов.
Периоды интерфазы
| Периоды | Число хромосом и хроматид | Процессы |
| Пресинтетический (G1) | 2n2c | Увеличивается объем цитоплазмы и количество органоидов, происходит рост клетки после предыдущего деления. |
| Синтетический (S) | 2n4c | Происходит удвоение генетического материала (репликация ДНК), синтез белковых молекул, с которыми связывается ДНК, и превращение каждой хромосомы в две хроматиды. |
| Постсинтетический (G2) | 2n4c | Усиливаются процессы биосинтеза, происходит деление митохондрий и хлоропластов, удваиваются центриоли. |
Деление эукариотических клеток
Основой размножения и индивидуального развития организмов является деление клетки.
Эукариотические клетки имеют три способа деления:
- амитоз (прямое деление),
- митоз (непрямое деление),
- мейоз (редукционное деление).
Амитоз — редкий способ деления клетки, характерный для стареющих или опухолевых клеток. При амитозе ядро делится путём перетяжки и равномерное распределение наследственного материала не обеспечивается. После амитоза клетка не способна вступать в митотическое деление.
Митоз
Митоз состоит из четырёх фаз.
Фазы митоза
| Фазы | Число хромосом и хроматид | Процессы |
| Профаза | 2n4c | Хромосомы спирализуются, центриоли (у животных клеток) расходятся к полюсам клетки, распадается ядерная оболочка, исчезают ядрышки, и начинает формироваться веретено деления. |
| Метафаза | 2n4c | Хромосомы, состоящие из двух хроматид, прикрепляются своими центромерами (первичными перетяжками) к нитям веретена деления. При этом все они располагаются в экваториальной плоскости. Эта структура называется метафазной пластинкой. |
| Анафаза | 2n2c | Центромеры делятся, и нити веретена деления растягивают отделившиеся друг от друга хроматиды к противоположным полюсам. Теперь разделённые хроматиды называются дочерними хромосомами. |
| Телофаза | 2n2c | Дочерние хромосомы достигают полюсов клетки, деспирализуются, нити веретена деления разрушаются, вокруг хромосом образуется ядерная оболочка, ядрышки восстанавливаются. Два образовавшихся ядра генетически идентичны. После этого следует цитокинез (деление цитоплазмы), в результате которого образуются две дочерние клетки. Органоиды распределяются между ними более или менее равномерно. |
Биологическое значение митоза:
- достигается генетическая стабильность;
- увеличивается число клеток в организме;
- происходит рост организма;
- возможны явления регенерации и бесполого размножения у некоторых организмов.
Мейоз
Как и митоз, каждое из мейотических делений состоит из четырёх фаз.
Фазы мейоза
| Фазы | Число хромосом и хроматид | Процессы |
| Профаза I | 2n4c | Происходят процессы, аналогичные процессам профазы митоза. Кроме того, гомологичные хромосомы, представленные двумя хроматидами, сближаются и «слипаются» друг с другом. Этот процесс называется конъюгацией. При этом происходит обмен участков гомологичных хромосом — кроссинговер (перекрест хромосом), то есть обмен наследственной информацией. После конъюгации гомологичные хромосомы отделяются друг от друга. |
| Метафаза I | 2n4c | Происходят процессы, аналогичные процессам метафазы митоза. |
| Анафаза I | 1n2c | В отличие от анафазы митоза, центромеры не делятся и к полюсам клетки отходит не по одной хроматиде от каждой хромосомы, а по одной хромосоме, состоящей из двух хроматид и скреплённой общей центромерой. |
| Телофаза I | 1n2c | Образуются две клетки с гаплоидным набором. |
| Интерфаза | 1n2c | Короткая. Репликации (удвоения) ДНК не происходит и, следовательно, диплоидность не восстанавливается. |
| Профаза II | 1n2c | Аналогичны процессам во время митоза. |
| Метафаза II | 1n2c | Аналогичны процессам во время митоза. |
| Анафаза II | 1n1c | Аналогичны процессам во время митоза. |
| Телофаза II | 1n1c | Аналогичны процессам во время митоза. |
Биологическое значение мейоза:
- основа полового размножения;
- основа комбинативной изменчивости.
Деление прокариотических клеток
У прокариот митоза и мейоза нет. Бактерии размножаются бесполым путём — делением клетки при помощи перетяжек или перегородок, реже почкованием. Этим процессам предшествует удвоение кольцевой молекулы ДНК.
Кроме того, для бактерий характерен половой процесс — конъюгация. При конъюгации по специальному каналу, образующемуся между двумя клетками, фрагмент ДНК одной клетки передаётся другой клетке, то есть изменяется наследственная информация, содержащаяся в ДНК обоих клеток. Поскольку количество бактерий при этом не увеличивается, для корректности используют понятие «половой процесс», но не «половое размножение».
Источник: examer.ru
Клеточный цикл.
Закономерные изменения структурно-функциональных характеристик клетки во времени составляют содержание ее жизненного цикла (клеточного цикла). Клеточный цикл- это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти.
Обязательным компонентом клеточного цикла является митотический цикл-комплекс взаимосвязанных и детерминированных хронологически событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления. Митотический цикл включает в себя митоз ,а также период покоя (G0), постмитотический (G1), синтетический (S) и предмитотический(G2) периоды интерфазы.
Интерфаза (периоды и процессы, проходящие здесь).
Интерфаза – это период между двумя клеточными делениями. В интерфазе ядро компактное, не имеет выраженной структуры, хорошо видны ядрышки. Совокупность интерфазных хромосом представляет собой хроматин . В состав хроматина входят: ДНК, белки и РНК в соотношении 1 : 1,3 : 0,2, а также неорганические ионы. Структура хроматина изменчива и зависит от состояния клетки.
Период покоя клетки ( G 0)- в период покоя судьба клетки не известна: она либо может начать подготовку к делению, либо погибнуть.
Постмитотический период ( G 1 ) . Фаза G1 – это основное рабочее состояние клетки. В этом состоянии идет транскрипция и трансляция, восстановление объема и внутреннего содержания клетки,идет размножение пластид и митохондрий.
Синтетический период ( S 1) – это период, когда ДНК в ядре удваивается. Репликация ДНК начинается во многих, однако строго определенных, местах, причем где-то раньше, где-то позже; тем не менее, к концу S-фазы каждая молекула ДНК удваивается полностью. В S-фазе в клетке активно синтезируются гистоны и прочие белки хроматина.
Среди белков хроматина имеется очень малая по количеству, но очень разнообразная и важная часть – специфические генные регуляторы (это те белковые репрессоры и активаторы, которые включают и выключают гены). Генов – десятки тысяч. Регуляторов меньше, так как каждый включает или выключает многие гены – иначе мы имели бы свой регулятор на каждый ген и впали бы в порочный круг. Важно подчеркнуть, что каждая клетка многоклеточного организма несет в себе все гены, присущие этому организму, но в каждой конкретной клетке работает только малая часть генов, тогда как остальные нужны в других типах клеток или в другие периоды жизни. Гены включаются и выключаются по мере необходимости, но при делении клеток определенного типа важно, чтобы включенные и выключенные состояния генов, характерные для данного типа, в целом были унаследованы. При репликации ДНК удваивается, и надо, чтобы регуляторные белки не только были дополнительно синтезированы в таком же количестве, которое было исходно, но и сели на свои места. Это достигается за счет кооперативного эффекта , который проявляют регуляторные белки, – наличие молекулы регуляторного белка, связанной с ДНК, провоцирует в своей непосредственной близости связывание такого же белка с таким же регуляторным сайтом вновь синтезированной ДНК. Об этом феномене принято говорить как об эпигенетическом наследовании состояния гена.
И в то же время репликация – это именно тот критический момент, когда многие гены выключаются или включаются в ходе индивидуального развития. В течение периода G1 среди других белков могут быть синтезированы новые регуляторы, и во время S перода они могут успешно конкурировать со старыми за вновь синтезированные регуляторные области ДНК. Или, наоборот, бывают недосинтезированы старые регуляторы, в результате созданные заново регуляторные области ДНК оказываются не занятыми или занятыми регуляторами, сродство которых к ним меньше. Кроме того, каждый белок-регулятор в моменты репликации ДНК вынужден конкурировать за те участки вновь синтезируемой ДНК, к которым он специфичен, с таким неспецифическим репрессором генной активности, как линкерный гистон Н1 (это тот гистон, который связывается с ДНК после того, как остальные гистоны образовали бусы из нуклеосом, и укладывает их в фибриллу диаметром 30 нм). Так, за счет некоторых изменений в присутствии регуляторов на регуляторных последовательностях ДНК тех или иных генов, в ходе индивидуального развития многоклеточного организма клетки и приобретают новые свойства.
Наконец, в клетке есть еще одна структура, удваивающаяся именно в S-периоде. Это центросома. В периоде G1 центросома выглядит так:
аморфное образование, внутри него находятся две расположенные перпендикулярно друг другу центриоли (но у растений центриолей нет). Центросома является местом, откуда формируется такой элемент цитоскелета, как микротрубочки. В интерфазе миркотрубочки растут от центросомы по направлению ко всей клеточной периферии. Некоторые из них становятся нестабильными и быстро разбираются на отдельные молекулы тубулина. В конце периода G1 центриоли расходятся на несколько микрон. А в S-периоде рядом с каждой центриолью строится вторая центриоль, и центросома удваивается.
Предмитотический период ( G 2) – подготовка к делению. На данной стадии нарабатываются определенные белки. В это время завершается формирование двух центросом, а система интерфазных микротрубочек начинает разрушаться, высвобождая тубулин, из которого микротрубочки состоят. Хромосомы в это время уже начинают дополнительно конденсироваться.Клетка готова к делению.
C обственно митоз.
Митоз-способ деления ядра,которое приводит к образованию двух дочерних клеток,в каждой из которых имеется точно такой же набор хромосом что и в родительских клетках. Собственно митоз также делится на несколько стадий. Митоз наступает при появлении в клетке специального митоз-стимулирующего фактора, который не может возникнуть, пока в клетке не закончилась репликация ДНК и другие подготовительные процессы. Под действием этого фактора запускается каскад фосфорилирования множества белков. В фосфорилированном состоянии они начинают активно функционировать. Один из наиболее интенсивно фосфорилируемых белков (до 6 фосфатных групп на молекулу) – это гистон Н1. При этом он теряет в сродстве с ДНК (так как его положительный заряд частично компенсируется отрицательно заряженными фосфатными группами), и с ней связываются другие белки, специфичные именно для митоза, что приводит к гораздо более плотной упаковке хромосом, чем в интерфазе. Еще один белок, который фосфорилируется в том же каскаде, запускающем митоз — когезин. В нефосфорилированном состоянии он соединяет вместе две сестринские хроматиды, образовавшиеся в результате репликации ДНК в S-фазе, образуя своего рода кольца вокруг пары хроматид. Фосфорилирование когезина в начале мейоза приводит к раскрытию колец и рассоедниению сестринских хроматид, за исключением центромеры,. Имеется механизм, который в этом районе снова фосфорилирует когезин, так что именно здесь сестринские хроматиды остаются соединенными друг с другом.
Первая стадия митоза – профаза . Главное, что происходит в профазе, – дополнительная упаковка (конденсация ) хромосом. В такой степени, что они становятся похожими сначала на спутанные нити, видимые в световой микроскоп.
В профазе происходят важные события и в цитоплазме. Имевшиеся в клетке микротрубочки деполимеризуются. При этом клетка как правило теряет свою специфическую форму и округляется. Вокруг центросом образуется так называемая звезда – система из радиально расходящихся микротрубочек, которые постепенно удлиняются. Микротрубочки в процессе митоза начинают обновляться в 20 раз быстрее, чем в интерфазе, и небольшому числу длинных микротрубочек приходит на смену множество коротких. Интенсивная сборка и разборка микротрубочек необходима для правильного течения митоза.
Когда микротрубочки двух звезд достигают друг друга, центросомы начинают расходиться к разным концам клетки и становятся ее полюсами, а сами микротрубочки формируют веретено деления . Дело в том, что многие микротрубочки, исходящие от разных полюсов навстречу друг другу, соединяются друг с другом определенными белками, которые стабилизируют их и предотвращают от деполимеризации.
Затем наступает прометафаза , которая знаменуется важнейшим событием – ядерная мембрана дефрагментируется на пузырьки и ядро исчезает как структура. При этом происходит деполимеризация ламины ядерного скелета, состоящего из филаментов определенных белков, подстилающих ядерную мембрану.Этот процесс также связан с фосфорилированием этих белков. Содержимое ядра объединяется с цитоплазмой. Тем самым восстанавливается состояние, похожее на прокариотическое, при котором ДНК находится в том же компартменте, что и рибосомы. Во время деления ядро исчезает. Это, по-видимому, указывает на то, что ядро – это временная рабочая структура, призванная разобщить тарнскрипцию и трансляцию, хотя бы ценой существенных энергетических затрат на ядерный транспорт и на то, чтобы от него, ядра, избавляться при всяком делении клетки и восстанавливать после него.
В прометафазе хромосомы конденсируются окончательно и принимают вид парных образований, напоминающих двойные палочки или червяков, причем каждая пара соединяется в месте своего рода перетяжки – это называется метафазные хромосомы .
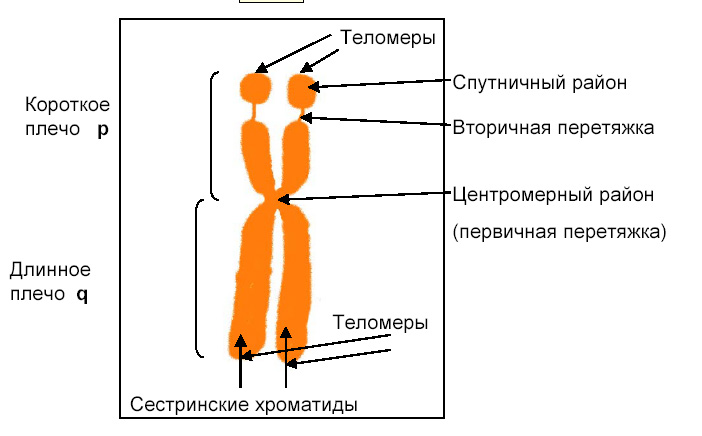
(Теломера – это конец хромосомы, имеющий специфическую последовательность нуклеотидов. Вторичная перетяжка соответствует ядрышку – это место, где находятся гены рРНК – оно не конденсируется в той же степени, что и остальная хромосома. Спутник – это участок «нормальной» хромосомы за вторичной перетяжкой. Вторичная перетяжка и соответственно спутник есть далеко не на всех хромосомах, поэтому они помогают их идентифицировать.)
Источник: MirZnanii.com