Схема строения прокариотической клетки
У современных и ископаемых организмов известны два типа клеток: прокариотическая и эукариотическая. Они столь резко различаются по особенностям строения, что это послужило для выделения двух надцарств живого мира — прокариот, т.е. доядерных, и эукариот, т.е. настоящих ядерных организмов. Промежуточные формы между этими крупнейшими таксонами живого пока неизвестны.
Основные признаки и отличия прокариотических и эукариотических клеток (таблица):
|
Признаки |
Прокариоты |
Эукариоты |
|
ЯДЕРНАЯ МЕМБРАНА |
Отсутствует |
Имеется |
|
ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА |
Имеется |
Имеется |
|
МИТОХОНДРИИ |
Отсутствуют |
Имеются |
|
ЭПС |
Отсутствует |
Имеется |
|
РИБОСОМЫ |
Имеются |
Имеются |
|
ВАКУОЛИ |
Отсутствуют |
Имеются (особенно характерны для растений) |
|
ЛИЗОСОМЫ |
Отсутствуют |
Имеются |
|
КЛЕТОЧНАЯ СТЕНКА |
Имеется, состоит из сложного гетерополимерного вещества |
Отсутствует в животных клетках, в растительных состоит из целлюлозы |
|
КАПСУЛА |
Если имеется, то состоит из соединений белка и сахара |
Отсутствует |
|
КОМПЛЕКС ГОЛЬДЖИ |
Отсутствует |
Имеется |
|
ДЕЛЕНИЕ |
Простое |
Митоз, амитоз, мейоз |
Основное отличие прокариотических клеток от эукариотических заключается в том, что их ДНК не организована в хромосомы и не окружена ядерной оболочкой. Эукариотические клетки устроены значительно сложнее. Их ДНК, связанная с белком, организована в хромосомы, которые располагаются в особом образовании, по сути самом крупном органоиде клетки — ядре. Кроме того, внеядерное активное содержимое такой клетки разделено на отдельные отсеки с помощью эндоплазматической сети, образованной элементарной мембраной.
кариотические клетки обычно крупнее прокариотических. Их размеры варьируют от 10 до 100 мкм, тогда как размеры клеток прокариот (различных бактерий, цианобактерий — сине- зеленых водорослей и некоторых других организмов), как правило, не превышают 10 мкм, часто составляя 2-3 мкм. В эукариотической клетке носители генов — хромосомы — находятся в морфологически оформленном ядре, отграниченном от остальной клетки мембраной. В исключительно тонких, прозрачных препаратах живые хромосомы можно видеть с помощью светового микроскопа. Чаще же их изучают на фиксированных и окрашенных препаратах.
Хромосомы состоят из ДНК, которая находится в комплексе с белками- гистонами, богатыми аминокислотами аргинином и лизином. Гистоны составляют значительную часть массы хромосом.
Эукариотическая клетка имеет разнообразные постоянные внутриклеточные структуры — органоиды (органеллы), отсутствующие в прокариотической клетке.
Прокариотические клетки могут делиться на равные части перетяжкой или почковаться, т.е. образовывать дочернюю клетку меньшего размера, чем материнская, но никогда не делятся путем митоза. Клетки эукариотических организмов, напротив, делятся путем митоза (исключая некоторые очень архаичные группы). Хромосомы при этом "расщепляются" продольно (точнее, каждая нить ДНК воспроизводит около себя свое подобие), и их "половинки" — хроматиды (полноценные копии нити ДНК) расходятся группами к противоположным полюсам клетки. Каждая из образующихся затем клеток получает одинаковый набор хромосом.
Рибосомы прокариотической клетки резко отличаются от рибосом эукариот по величине. Ряд процессов, свойственных цитоплазме многих эукариотических клеток, — фагоцитоз, пиноцитоз и циклоз (вращательное движение цитоплазмы) — у прокариот не обнаружен. Прокариотической клетке в процессе обмена веществ не требуется аскорбиновая кислота, но эукариотические не могут без нее обходиться.
Существенно различаются подвижные формы прокариотических и эукариотических клеток. Прокариоты имеют двигательные приспособления в виде жгутиков или ресничек, состоящих из белка флагеллина. Двигательные приспособления подвижных эукариотических клеток получили название ундулиподиев, закрепляющихся в клетке с помощью особых телец кинетосом. Электронная микроскопия выявила структурное сходство всех ундулиподиев эукариотических организмов и резкие их отличия от жгутиков прокариот
1. Строение эукариотической клетки.
Клетки, образующие ткани животных и растений, значительно различаются по форме, размерам и внутреннему строению. Однако все они обнаруживают сходство в главных чертах процессов жизнедеятельности, обмена веществ, в раздражимости, росте, развитии, способности к изменчивости.
Клетки всех типов содержат два основных компонента, тесно связанных между собой, — цитоплазму и ядро. Ядро отделено от цитоплазмы пористой мембраной и содержит ядерный сок, хроматин и ядрышко.
лужидкая цитоплазма заполняет всю клетку и пронизана многочисленными канальцами. Снаружи она покрыта цитоплазматической мембраной. В ней имеются специализированные структуры-органоиды, присутствующие в клетке постоянно, и временные образования — включения. Мембранные органоиды: наружная цитоплазматическая мембрана (HЦM), эндоплазматическая сеть (ЭПС), аппарат Гольджи, лизосомы, митохондрии и пластиды. В основе строения всех мембранных органоидов лежит биологическая мембрана. Все мембраны имеют принципиально единый план строения и состоят из двойного слоя фосфолипидов, в который с различных сторон ива разную глубину погружены белковые молекулы. Мембраны органоидов отличаются друг от друга лишь наборами входящих в них белков.
Цитоплазматическая мембрана. У всех клеток растений, многоклеточных животных, у простейших и бактерий клеточная мембрана трехслойна: наружный и внутренний слои состоят из молекул белков, средний — из молекул липидов. Она ограничивает цитоплазму от внешней среды, окружает все органоиды клетки и представляет собой универсальную биологическую структуру. В некоторых клетках наружная оболочка образована несколькими мембранами, плотно прилегающими друг к другу. В таких случаях клеточная оболочка становится плотной и упругой и позволяет сохранить форму клетки, как, например, у эвглены и инфузории туфельки. У большинства растительных клеток, помимо мембраны, снаружи имеется еще толстая целлюлозная оболочка — клеточная стенка.
а хорошо различима в обычном световом микроскопе и выполняет опорную функцию за счет жесткого наружного слоя, придающего клеткам четкую форму.
На поверхности клеток мембрана образует удлиненные выросты — микроворсинки, складки, впячивания и выпячивания, что во много раз увеличивает всасывающую или выделительную поверхность. С помощью мембранных выростов клетки соединяются друг с другом в тканях и органах многоклеточных организмов, на складках мембран располагаются разнообразные ферменты, участвующие в обмене веществ. Отграничивая клетку от окружающей среды, мембрана регулирует направление диффузии веществ и одновременно осуществляет активный перенос их внутрь клетки (накопление) или наружу (выделение). За счет этих свойств мембраны концентрация ионов калия, кальция, магния, фосфора в цитоплазме выше, а концентрация натрия и хлора ниже, чем в окружающей среде. Через поры наружной мембраны из внешней среды внутрь клетки проникают ионы, вода и мелкие молекулы других веществ. Проникновение в клетку относительно крупных твердых частиц осуществляется путем фагоцитоза (от греч. "фаго” — пожираю, "питое” — клетка)[2]. При этом наружная мембрана в месте контакта с частицей прогибается внутрь клетки, увлекая частицу в глубь цитоплазмы, где она подвергается ферментативному расщеплению. Аналогичным путем в клетку попадают и капли жидких веществ; их поглощение называется пиноцитозом (от греч.
uot;пино” — пью, "цитос” — клетка). Наружная клеточная мембрана выполняет и другие важные биологические функции.
Цитоплазма на 85 % состоит из воды, на 10 % — из белков, остальной объем приходится на долю липидов, углеводов, нуклеиновых кислот и минеральных соединений; все эти вещества образуют коллоидный раствор, близкий по консистенции глицерину. Коллоидное вещество клетки в зависимости от ее физиологического состояния и характера воздействия внешней среды имеет свойства и жидкости, и упругого, более плотного тела. Цитоплазма пронизана каналами различной формы и величины, которые получили название эндоплазматической сети. Их стенки представляют собой мембраны, тесно контактирующие со всеми органоидами клетки и составляющие вместе с ними единую функционально-структурную систему для осуществления обмена веществ и энергии и перемещения веществ внутри клетки.
В стенках канальцев располагаются мельчайшие зернышки—гранулы, называемые рибосомами. Такая сеть канальцев называется гранулярной. Рибосомы могут располагаться на поверхности канальцев разрозненно или образуют комплексы из пяти-семи и более рибосом, называемые полисомами. Другие канальцы гранул не содержат, они составляют гладкую эндоплазматическую сеть. На стенках располагаются ферменты, участвующие в синтезе жиров и углеводов.
Внутренняя полость канальцев заполнена продуктами жизнедеятельности клетки. Внутриклеточные канальцы, образуя сложную ветвящуюся систему, регулируют перемещение и концентрацию веществ, разделяют различные молекулы органических веществ и этапы их, синтеза. На внутренней и внешней поверхности мембран, богатых ферментами, осуществляется синтез белков, жиров и углеводов, которые либо используются в обмене веществ, либо накапливаются в цитоплазме в качестве включений, либо выводятся наружу.
Рибосомы встречаются во всех типах клеток — от бактерий до клеток многоклеточных организмов. Это округлые тельца, состоящие из рибонуклеиновой кислоты (РНК) и белков почти в равном соотношении. В их состав непременно входит магний, присутствие которого поддерживает структуру рибосом. Рибосомы могут быть связаны с мембранами эндоплазматической сети, с наружной клеточной мембраной или свободно лежать в цитоплазме. В них осуществляется синтез белков. Рибосомы кроме цитоплазмы встречаются в ядре клетки. Они образуются в ядрышке и затем поступают в цитоплазму.
Комплекс Гольджи в растительных клетках имеет вид отдельных телец, окруженных мембранами. В животных клетках этот органоид представлен цистернами, канальцами и пузырьками. В мембранные трубки комплекса Гольджи из канальцев эндоплазматической сети поступают продукты секреции клетки, где они химически перестраиваются, уплотняются, а затем переходят в цитоплазму и либо используются самой клеткой, либо выводятся из нее. В цистернах комплекса Гольджи происходит синтез полисахаридов и их объединение с белками, в результате чего образуются гликопротеиды.
Митохондрии — небольшие тельца палочковидной формы, ограниченные двумя мембранами. От внутренней мембраны митохондрии отходят многочисленные складки — кристы, на их стенках располагаются разнообразные ферменты, с помощью которых осуществляется синтез высокоэнергетического вещества — аденозинтрифосфорной кислоты (АТФ). В зависимости от активности клетки и внешних воздействий митохондрии могут перемещаться, изменять свои размеры, форму. В митохондриях найдены рибосомы, фосфолипиды, РНК и ДНК. С присутствием ДНК в митохондриях связывают способность этих органоидов к размножению путем образования перетяжки или почкованием в период деления клетки, а также синтез части митохондриальных белков.
Лизосомы — мелкие овальные образования, ограниченные мембраной и рассеянные по всей цитоплазме. Встречаются во всех клетках животных и растений. Они возникают в расширениях эндоплазматической сети и в комплексе Гольджи, здесь заполняются гидролитическими ферментами, а затем обособляются и поступают в цитоплазму. В обычных" условиях лизосомы переваривают частицы, попадающие в клетку путем фагоцитоза, и органоиды отмирающих клеток. Продукты лизиса выводятся через мембрану лизосомы в цитоплазму, где они включаются в состав новых молекул. При разрыве лизоеомной мембраны ферменты поступают в цитоплазму и переваривают ее содержимое, вызывая гибель клетки.
Пластиды есть только в растительных клетках и встречаются, у большинства зеленых растений. В пластидах синтезируются и накапливаются органические вещества. Различают пластиды трех видов: хлоропласты, хромопласты и лейкопласты.
Хлоропласты — зеленые пластиды, содержащие зеленый пигмент хлорофилл. Они находятся в листьях, молодых стеблях, незрелых плодах. Хлоропласты окружены двойной мембраной. У высших растений внутренняя часть хлоропластов заполнена полужидким веществом, в котором параллельно друг другу уложены пластинки. Парные мембраны пластинок, сливаясь, образуют стопки, содержащие хлорофилл. В каждой стопке хлоропластов высших растений чередуются слои молекул белка и молекул липидов, а между ними располагаются молекулы хлорофилла. Такая слоистая структура обеспечивает максимум свободных поверхностей и облегчает захват и перенос энергии в процессе фотосинтеза.
Хромопласты — пластиды, в которых содержатся растительные пигменты (красный или бурый, желтый, оранжевый). Они сосредоточены в цитоплазме клеток цветков, стеблей, плодов, листьев растений и придают им соответствующую окраску. Хромопласты образуются из лейкопластов или хлоропластов в результате накопления пигментов каротиноидов.
Лейкопласты—бесцветные пластиды, располагающиеся в неокрашенных частях растений: в стеблях, корнях, луковицах и др. В лейкопластах одних клеток накапливаются зерна крахмала, в лейкопластах других клеток — масла, белки.
Все пластиды возникают из своих предшественников — пропластид. В них выявлена ДНК, которая контролирует размножение этих органоидов.
Клеточный центр, или центросома, играет важную роль при делении, клетки и состоит из двух центриолей. Он встречается у всех клеток животных и растений, кроме цветковых, низших грибов и некоторых, простейших. Центриоли в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В делящейся клетке первым делится клеточный центр, одновременно образуется ахроматиновое веретено, ориентирующее хромосомы при расхождении их к полюсам. В дочерние клетки отходит по одной центриоле.
У многих растительных и животных клеток имеются органоиды специального назначения: реснички, выполняющие функцию движения (инфузории, клетки дыхательных путей), жгутики (простейшие одноклеточные, мужские половые клетки у животных и растений и др.).
Включения — временные элемеаты, возникающие в клетке на определенной стадии ее жизнедеятельности в результате синтетической функции. Они либо используются, либо выводятся из клетки. Включениями являются также запасные питательные вещества: в растительных клетках—крахмал, капельки жира, белки, эфирные масла, многие органические кислоты, соли органических и неорганических кислот; в животных клетках — гликоген (в клетках печени и мышцах), капли жира (в подкожной клетчатке); Некоторые включения накапливаются в клетках как отбросы — в виде кристаллов, пигментов и др.
Вакуоли — это полости, ограниченные мембраной; хорошо выражены в клетках растений и имеются у простейших. Возникают в разных участках расширений эндоплазматической сети. И постепенно отделяются от нее. Вакуоли поддерживают тургорное давление, в них сосредоточен клеточный или вакуолярный сок, молекулы которого определяют его осмотическую концентрацию. Считается, что первоначальные продукты синтеза — растворимые углеводы, белки, пектины и др. — накапливаются в цистернах эндоплазматической сети. Эти скопления и представляют собой зачатки будущих вакуолей.
Цитоскелет. Одной из отличительных особенностей эукариотической клетки является развитие в ее цитоплазме скелетных образований в виде микротрубочек и пучков белковых волокон. Элементы цитоскелета тесно связаны с наружной цитоплазматической мембраной и ядерной оболочкой, образуют сложные переплетения в цитоплазме. Опорные элемеиты цитоплазмы определяют форму клетки, обеспечивают движение внутриклеточных структур и перемещение всей клетки.
Ядро клетки играет основную роль в ее жизнедеятельности, с его удалением клетка прекращает свои функции и гибнет. В большинстве животных клеток одно ядро, но встречаются и многоядерные клетки (печень и мышцы человека, грибы, инфузории, зеленые водоросли). Эритроциты млекопитающих развиваются из клеток-предшественников, содержащих ядро, но зрелые эритроциты утрачивают его и живут недолго.
Ядро окружено двойной мембраной, пронизанной порами, посредством которых оно тесно связано с каналами эндоплазматической сети и цитоплазмой. Внутри ядра находится хроматин — спирализованные участки хромосом. В период деления клетки они превращаются в палочковидные структуры, хорошо различимые в световой микроскоп. Хромосомы — это сложный комплекс белков с ДНК, называемый нуклеопротеидом.
Функции ядра состоят в регуляции всех жизненных отправлений клетки, которую оно осуществляет при помощи ДНК и РНК-материальных носителей наследственной информации. В ходе подготовки к делению клетки ДНК удваивается, в процессе митоза хромосомы расходятся и передаются дочерним клеткам, обеспечивая преемственность наследственной информации у каждого вида организмов.
Кариоплазма — жидкая фаза ядра, в которой в растворенном виде находятся продукты жизнедеятельности ядерных структур.
Ядрышко — обособленная, наиболее плотная часть ядра.
В состав ядрышка входят сложные белки и РНК, свободные или связанные фосфаты калия, магния, кальция, железа, цинка, а также рибосомы. Ядрышко исчезает перед началом деления клетки и вновь формируется в последней фазе деления.
Таким образом, клетка обладает тонкой и весьма сложной организацией. Обширная сеть цитоплазматических мембран и мембранный принцип строения органоидов позволяют разграничить множество одновременно протекающих в клетке химических реакций. Каждое из внутриклеточных образований имеет свою структуру и специфическую функцию, но только при их взаимодействии возможна гармоничная жизнедеятельность клетки.На основе такого взаимодействия вещества из окружающей среды поступают в клетку, а отработанные продукты выводятся из нее во внешнюю среду — так совершается обмен веществ. Совершенство структурной организации клетки могло возникнуть только в результате длительной биологической эволюции, в процессе которой выполняемые ею функции постепенно усложнялись.
Простейшие одноклеточные формы представляют собой и клетку, и организм со всеми его жизненными проявлениями. В многоклеточных организмах клетки образуют однородные группы — ткани. В свою очередь ткани формируют органы, системы, и их функции определяются общей жизнедеятельностью целостного организма.
Помимо организмов с типичной клеточной организацией {эукариотические клетки) существуют относительно простые, доядерные, или прокариотические, клетки — бактерии и синезеленые, у которых отсутствуют оформленное ядро, окруженное ядерной мембраной, и высокоспециализированные внутриклеточные органоиды. Особую форму организации живого представляют вирусы и бактериофаги (фаги). Их строение крайне упрощено: они состоят из ДНК (либо РНК) и белкового футляра. Свои функции обмена веществ и размножения вирусы и фаги осуществляют только внутри клеток другого организма: вирусы — внутри клеток растений и животных, фаги — в бактериальных клетках как паразиты на, генетическом уровне.
К прокариотам относят бактерии и сине-зелёные водоросли (цианеи). Наследственный аппарат прокариот представлен одной кольцевой молекулой ДНК, не образующей связей с белками и содержащей по одной копии каждого гена — гаплоидные организмы. В цитоплазме имеется большое количество мелких рибосом; отсутствуют или слабо выражены внутренние мембраны. Ферменты пластического обмена расположены диффузно. Аппарат Гольджи представлен отдельными пузырьками. Ферментные системы энергетического обмена упорядоченно расположены на внутренней поверхности наружной цитоплазматической мембраны. Снаружи клетка окружена толстой клеточной стенкой. Многие прокариоты способны к спорообразованию в неблагоприятных условиях существования; при этом выделяется небольшой участок цитоплазмы содержащий ДНК, и окружается толстой многослойной капсулой. Процессы метаболизма внутри споры практически прекращаются. Попадая в благоприятные условия, спора преобразуется в активную клеточную форму. Размножение прокариот происходит простым делением надвое.
Средняя величина прокариотических клеток 5 мкм. У них нет никаких внутренних мембран, кроме впячиваний плазматической мембраны. Пласты отсутствуют. Вместо клеточного ядра имеется его эквивалент (нуклеоид), лишенный оболочки и состоящий из одной-единственной молекулы ДНК. Кроме того бактерии могут содержать ДНК в форме крошечных плазмид, сходных с внеядерными ДНК эукариот.
В прокариотических клетках, способных к фотосинтезу (сине-зеленые водоросли, зеленые и пурпурные бактерии) имеются различно структурированные крупные впячивания мембраны – тилакоиды, по своей функции соответствующие пластидам эукариот. Эти же тилакоиды или – в бесцветных клетках – более мелкие впячивания мембраны (а иногда даже сама плазматическая мембрана) в функциональном отношении заменяют митохондрии. Другие, сложно дифференцированные впячивания мембраны называют мезасомами; их функция не ясна.
Только некоторые органеллы прокариотической клетки гомологичны соответствующим органеллам эукариот. Для прокариот характерно наличие муреинового мешка – механически прочного элемента клеточной стенки
Сравнительная характеристика клеток растений, животных, бактерий, грибов
При сравнении бактерий с эукариотами можно выделить единственное сходство — наличие клеточной стенки, а вот сходства и различия эукариотических организмов заслуживают более пристального внимания. Следует начать сравнение с компонентов, которые свойственны и растениям, и животным, и грибам. Это ядро, митохондрии, Аппарат (комплекс) Гольджи, эндоплазматический ретикулум (или эндоплазматическая сеть) и лизосомы. Они характерны для всех организмов, имеют сходное строение и выполняют одинаковые функции. Теперь следует акцентировать внимание на различиях. Растительная клетка, в отличие от животной, имеет клеточную стенку, состоящую из целлюлозы. Кроме того, существую органеллы свойственные растительным клеткам — пластиды и вакуоли. Наличие этих компонентов обусловлено необходимостью растений поддерживать форму, при отсутствии скелета. Есть отличия и в особенностях роста. У растений он происходит в основном за счет увеличения размера вакуолей и растяжения клеток, в то время как у животных происходит увеличение объема цитоплазмы, а вакуоль вовсе отсутствует. Пластиды (хлоропласты, лейкопласты, хромопласты) характерны преимущественно для растений, поскольку их основная задача — это обеспечить автотрофный способ питания. У животных в противовес растениям существуют пищеварительные вакуоли, которые обеспечивают гетеротрофный способ питания. Грибы занимают особое положение и для их клеток характерны признаки свойственные и для растений, и для животных. Подобно животным грибам присущ гетеротрофный тип питания, содержащая хитин клеточная оболочка, а основным запасающим веществом является гликоген. В то же время для них, как для растений, характерен неограниченный рост, неспособность к передвижению и питание путем всасывания.
Сохранить в соцсетях:
Источник: krasgmu.net
Лекция № 3
Тема лекции: Основы цитологии. Эукариоты. Мембранные компоненты.
План лекции:
1. Строение эукариотической клетки
2. Цитоплазматическая мембрана
3. Мембранные компоненты клетки
1. Строение эукариотической клетки
Эукариотическая клетка имеет сложное строение. Бывают одноклеточные эукариоты (микроскопические грибы, простейшие) имногоклеточные. Последние могут состоять из нескольких клеток (круглые гельминты) и из миллиардов (млекопитающие). Организм человека состоит из 1013клеток, слона – 6,51014.
Размер эукариотической клетки – 0,1 мкм ÷ 20 мкм (в среднем). Бывают очень крупные клетки (яйцеклетки), нервные клетки достигают в размере нескольких метров.
Основные структурные элементы клетки: цитоплазматическая мембрана (плазмалемма); цитоплазма (цитозоль – вязкая жидкость плюс мембранные и немембранные компоненты), а также ядро.
Мембранные компоненты клетки:
— цитоплазматическая мембрана;
— ядро;
— вакуолярная система (эндоплазматическая сеть, аппарат Гольджи,
лизосомы, вакуоли растительных клеток);
— митохондрии;
— пластиды.
Немембранные компоненты клетки:
— хромосомы;
— цитоскелет;
— клеточный центр и центриоли;
— специальные органеллы движения (реснички и жгутики);
— рибосомы;
— включения.
Источник: StudFiles.net
Прокариотические клетки — это наиболее примитивные, очень просто устроенные, сохраняющие черты глубокой древности организмы. К прокариотическим (или доядерным) организмам относят бактерии и синезеленые водоросли (цианобактерии). На основании общности строения и резких отличий от других клеток их выделяют в самостоятельное царство дробянки.
Рассмотрим строение прокариотической клетки на примере бактерий.
Генетический аппарат представлен ДНК единственной кольцевой хромосомы, находится в цитоплазме и не отграничен от нее оболочкой. Такой аналог ядра называют нуклеоидом. ДНК не образует комплексов с белками и поэтому все гены, входящие в состав хромосомы, «работают», т.е. с них непрерывно считывается информация.
Прокариотическая клетка окружена мембраной, отделяющей цитоплазму от клеточной стенки, образованной из сложного, высокополимерного вещества. В цитоплазме органелл мало, но присутствуют многочисленные мелкие рибосомы (бактериальные клетки содержат от 5000 до 50 000 рибосом).
Цитоплазма пронизана мембранами, образующими эндоплазматическую сеть, в ней и находятся рибосомы, осуществляющие синтез белков.
Внутренняя часть клеточной стенки представлена плазматической мембраной, выпячивания которой в цитоплазму образуют мезосомы, участвующие в построении клеточных перегородок, репродукции, и являются местом прикрепления ДНК. Дыхание у бактерий осуществляется в мезосомах, у сине-зеленых водорослей в цитоплазматических мембранах.
У многих бактерий внутри клетки откладываются запасные вещества: полисахариды, жиры, полифосфаты. Резервные вещества, включаясь в обмен веществ, могут продлевать жизнь клетки в отсутствие внешних источников энергии.
Выделяют 4 основные формы клеток:
1) Кокки – бактерии, имеющие сферическую форму. Сферические бактерии после деления могут образовывать:
а) диплококки – две клетки в одной капсуле. Представители: пневмококк — возбудитель пневмонии;
б) стрептококки – образованны кокками в виде цепочки. Представители: возбудители ангины и скарлатины;
в) стафилококки – напоминают виноградную гроздь. Представители: разные штаммы стафилококков вызывают фурункулёз, воспаление лёгких, пищевые отравления и некоторые другие заболевания.
2) Бациллы – прямые, палочковидные бактерии:
а) неспорообразующие палочки называют бактериями. Представители: обычные кишечные симбионты, возбудители брюшного тифа, клубеньковые бактерии;
б) спорообразующие палочки называют бациллами. Представители: очень много в почве, например, азотофиксирующие бактерии, возбудители сибирской язвы, возбудитель туберкулёза – палочка Коха.
3) Спириллы, спирохеты – спиралевидной формы.
а ) спириллы – спиральные палочки с одним жгутиком. Представители: обычные обитатели ротовой полости.
б) спирохеты – форма клеток очень сложна, но есть различия по способу передвижения. Представители: обычные обитатели ротовой полости, возбудитель сифилиса.
4) Вибрионы – короткие палочки, всегда изогнутые в виде запятой. Представители: возбудитель холеры.
Прокариоты появились на Земле около 3,5 млрд лет назад и были, вероятно, первой клеточной формой жизни, дав начало современным прокариотам и эукариотам.
Источник: biology623.blogspot.com
Для всех микроорганизмов, входящих в царство, характерен прокариотический тип организации клетки, что определяется особенностями их ультраструктуры, а также строения и функций ряда макромолекул. Из всех известных клеток прокариотическая самая простая и, вероятно, первая клетка, возникшая около 3,6 млрд. лет назад.
В настоящее время предполагают, что в какой-то момент времени эволюция клеток пошла по двум самостоятельным направлениям. Появились две группы организмов —прокариоты, у которых ядерный материал не был ограничен оболочкой, и эукариоты, имеющие оформленное ядро с ядерной оболочкой.
Основные отличия прокариотов от эукариотов состоят в следующем:
в клетках прокариотов отсутствуют компартменты, или органеллы, ограниченные от цитоплазмы специализированными внутриклеточными липопротеидными мембранами: эндоплазматическая сеть (ретикулюм), митохондрии, аппарат Гольджи, лизосомы, хлоропласты;
ядерная структура прокариотов, называемая нуклеоидом, не имеет ядерной оболочки с поровым комплексом и представлена макромолекулой ДНК с белками (без гистонов);
геном прокариотической клетки организован в одну кольцевую хромосому, которая представляет собой единый репликон и не делится митозом;
дополнительные репликоны могут быть представлены кольцевыми молекулами плазмидных ДНК;
прокариотическая клетка содержит только один тип рибосом с константой седиментации 708, причем часть рибосом ассоциирована с цитоплазматической мембраной, что никогда не наблюдается у эукариотов;
клеточная стенка прокариотов содержит характерный только для бактерий биогетерополимер — пептидогликан.
Некоторые прокариоты обладают структурами, отсутствующими у эукариотов:
подвижные бактерии имеют особые бактериальные жгутики из белков-флагеллинов;
спорообразующие формы бактерий в неблагоприятных условиях превращаются в уникальные по степени устойчивости типы покоящихся клеток — бактериальные споры;
клетки прокариотов очень малы; диаметр большинства клеток бактерий не превышает 1 мкм, однако длина может быть значительной, например, у некоторых спирохет — до 500 мкм. Малые размеры прокариотов связаны, как полагают, с отсутствием в их ультраструктуре специализированных мембранных систем, что затрудняет координацию внутриклеточных процессов пропорционально увеличению размеров клетки.
Клеточное строение четко отделяет прокариотов от вирусов. Подчеркивая примитивность организации бактериальных клеток, необходимо однако отметить, что они эволюционировали в своем направлении в течение гораздо большего времени, чем эукарио-тические, и, хотя эволюционные возможности прокариотической клетки, по всей видимости, ограничены, в процессе эволюции происходили изменения их клеточной организации, что привело постепенно к ее усложнению.
По целому ряду признаков бактерии имеют принципиальные различия с эукариотами, и знание особенностей их устройства и функционирования позволяет понять возможность избирательного антимикробного действия химиотерапевтических препаратов. Применение электронной микроскопии и тонких цитохимических исследований позволило изучить их ультраструктуру (рис. 1). Обязательными компонентами бактериальной клетки являются цитоплазматическая мембрана, окружающая цитоплазму, в которой содержатся рибосомы и нуклеоид. Клетки всех бактерий, за исключением L-форм и микоплазм, имеют клеточную стенку. Другие структуры являются дополнительными и определяют морфологические и функциональные особенности различных видов: капсулы, жгутики, пили, споры, включения.

Рис. 1. Схема строения прокариотической клетки:
/ — капсула; 2 — клеточная стенка; 3 — цитоплазматическая мембрана; 4 — мезосомы; 5 — цитоплазма; 6 — нуклеоид; 7 — плазмида;
8 — рибосомы и полисомы; 9 — жгутики; 10 — пили; 11 — гранулы гликогена; 12 — липидные капельки; 13 — гранулы волютина; 14 — включения серы
Поверхностные структуры. Капсула — это наружный, самый верхний слизистый слой клетки различной толщины фибриллярной или глобулярной структуры. Она имеет полисахаридную, му-кополисахаридную или полипептидную природу и содержит до 98 % воды. В зависимости от толщины различают микрокапсулу (толщиной менее 0,2 мкм) и макрокапсулу. Капсула не является обязательным структурным элементом клетки. Биологический смысл капсулообразования определяется целым рядом функций, среди которых: защита от фагоцитов и вирусов, токсинов и радиации; иммунологическая мимикрия у патогенных бактерий; сохранение влаги в условиях пониженной влажности; прикрепление клетки к плотной поверхности.
Пили (фимбрии, ворсинки, реснички) — это прямые цилиндрические образования белковой природы длиной 0,3—10 мкм, диаметром до 10 нм, равномерно покрывающие поверхность клетки (до нескольких сотен на клетку), не выполняющие локомоторную функцию.
Различают пили общего типа, способствующие прикреплению бактериальной клетки к субстрату, клеткам человека (явление адгезии микроорганизмов) и пили половые, участвующие в передаче генетического материала от клетки-донора к клетке-реципиенту в процессе конъюгации, а также обуславливающие адсорбцию специфических бактериофагов на клетках.
Жгутики — органы движения бактерий в виде спирально изогнутых цилиндрических образований белковой природы (белки-флагеллины) на поверхности клетки длиной 3—12 мкм и толщиной 10—30 нм, прикрепленные базальным телом (системой дисков) к цитоплазматической мембране (см. вкл. I). Число и располо-
жение жгутиков может быть различным и является видовым признаком (рис. 2). Различают монотрихи (бактерии с одним жгутиком на конце), амфитрихи (бактерии с жгутиками, расположенными по полюсам), лофотрихи (клетки с пучком жгутиков на одном конце) и перитрихи (с 2—30 жгутиками по всему телу клетки).
Пили и жгутики не являются обязательными органоидами бактериальной клетки.
Клеточная стенка — один из основных структурных элементов бактерии, выполняющий механическую защиту клетки. Кроме микоплазм и L-форм, клетки всех бактерий покрыты клеточной стенкой, толщина которой у разных видов колеблется в пределах 0,01—14 мкм. Она представляет собой плотную эластичную структу —
Рис. 2. Основные формы бактерий (по А. А. Воробьеву и соавт., 1994):
/ — стафилококки; 2 — стрептококки; 3 — сарцины; 4 — гонококки;
5— пневмококки; 6— капсулы пневмококков; 7— коринебактерии дифтерии; 8 — клостридии; 9 — бациллы; 10 — вибрионы; 11 — спириллы; 12 — трепонсмы; — боррелии; 14 — лептоспиры; 15— актиномицеты; 16 — расположение жгутиков: а — монотрихи; б — лофотрихи; в — амфитрихи; г — перитрихи
ру, которая окружает протопласт клетки и придает ей постоянную форму и жесткость. Клеточная стенка препятствует осмотическому набуханию и разрыву клеток, когда они попадают в гипотоническую среду. Вода, другие малые молекулы и разные ионы легко проникают через крошечные поры в клеточной стенке, но через них не проходят крупные молекулы белков и нуклеиновых кислот.
Основным химическим компонентом клеточной стенки является специфический гетерополимер — пептидогликан (муреин, мукопептид, глюкозаминопептид, гликопептид), состоящий из цепочек, в которых чередуются остатки N-ацетилглюкозамина и М-ацетилмурамовой кислоты, соединенные между собой β-1,4-гликозидными связями. Это резко отличает оболочечные структуры бактерий от эукариотических и создает «ахиллесову пяту» бактерий, используемую для антимикробной химиотерапии.
Организация цитоплазмы. Цитоплазматическая мембрана (ЦМ) Носится к числу обязательных клеточных структур, имеет толщину 7—13 нм и располагается непосредственно под клеточной стенкой, ограничивая протопласт клетки. По своему строению мембраны бактериальных, животных и растительных клеток очень сходны. В настоящее время большинством ученых принята жидкостно-мозаичная модель строения ЦМ. Согласно этой модели ЦМ состоит из двойного слоя (15—30 % молекул фосфолипидов и триглицеридов с направленными внутрь гидрофобными концами и гидрофильными «головками» наружу. В него мозаично погружены молекулы белка (50—70%). В мембране имеются также углеводы (2—5 %) и РНК. ЦМ представляет собой пластическое «текучее» образование, которое играет важнейшую роль обмене веществ, является полупроницаемой структурой, поддерживает осмотическое давление, контролирует как поступление веществ в клетку, так и выведение конечных метаболитов по системе субстрат специфичных пермеаз (ферментов-переносчиков, локализованных на мембране). С ЦМ связаны процессы дыхания, доставляющие клетке энергию, то есть те функции, за которые в эукариотической клетке ответственны мембраны митохондрий и хлоропластов.
Выделяют так называемые мезосомы — впячивания ЦМ — смешанные мембранные системы, образованные трубочками, пузырьками и ламеллами. Предполагается выполнение ими функций центра дыхательной активности бактерий, участие в делении клетки и расхождении дочерних хромосом после репликации.
Цитоплазма заполняет собой объем бактерии, ограниченный ЦМ. Это сложная коллоидная система, которая состоит из бельков, нуклеиновых кислот, углеводов, липидов, минеральных веществ и 70—80 % воды. Цитоплазма является местонахождением внутриклеточных органелл (нуклеоида, рибосом, различных включений) и участвует во внутриклеточном метаболизме. Характер-
ными чертами организации цитоплазмы прокариотов по сравнению с животными и растительными клетками являются отсутствие эндоплазматического ретикулюма и высокая электронная плотность.
Нуклеоид — ядерный материал бактериальной клетки. Он представлен двойной нитью макромолекулы ДНК с молекулярной массой 2—310 в соединении с белками, среди которых отсутствуют характерные для эукариотов ядерные (гистоны и гистоноподобные) белки. В отличие от настоящего ядра эукариотических клеток нуклеоид не имеет ядерной перфорированной мембраны, не делится митозом и представляет собой в период деления одну кольцевую хромосому, кодирующую всю генетическую информацию.
Плазмиды — необязательные внутриклеточные структуры в виде внехромосомных кольцевых участков ДНК, способных к саморепликации. Обуславливают наследование дополнительных признаков: лекарственной устойчивости, токсигенности, бактериоциногенности и др.
Рибосомы — органеллы, в которых осуществляется синтез белка. Каждая рибосома имеет размеры 20х30х30 нм и константу седиментации 70S (так как при ультрацентрифугировании рибосомы оседают со скоростью около 70 единиц Сведенберга (S), в отличие от более крупных цитоплазматических рибосом эукариотов с константой седиментации 808). В свободном состоянии бактериальная рибосома находится в виде двух субъединиц — 30S и 50S, обе субъединицы содержат примерно по 40 % рибосомальной РНК и 60 % белка. Во время синтеза белка рибосомы с помощью информационной РНК образуют полисомы, обычно связанные с ЦМ. Бактерии могут содержать от 5000 до 50 000 рибосом, что зависит от возраста клетки и условий культивирования.
Знание различий между рибосомами бактерий и эукариотических клеток имеет важное значение для понимания механизмов антимикробного действия тех антибиотиков, которые подавляют синтез белка на бактериальных рибосомах и не затрагивают функции 80S рибосом.
Споры (эндоспоры) бактерий — покоящиеся формы некоторых видов грамположительных бактерий в неблагоприятных условиях внешней среды.
Спорообразование происходит в несколько стадий, при полном созревании споры вегетативная часть клетки лизируется и отмирает (см. вкл. I, II).
В процессе спорообразования (споруляции) можно выделить несколько основных этапов. Переходящая к спорообразованию клетка перестает расти; как правило, она содержит два и более нуклеоидов. На первом этапе часть клеточной ДНК локализуется в одном из полюсов клетки. Затем часть цитоплазмы с заключен-
Иной в ней хромосомой обособляется цитоплазматической мембраной, как бы врастающей в глубь клетки, при этом образуется проспорь, окруженная двойной мембранной оболочкой.
Затем между двумя мембранами идет формирование многослойной стенки и коры (кортекса) споры пептидогликановой природы. Снаружи мембран образуется также полипептидная оболочка и экзоспорий, окружающий спору в виде свободного чехла. Полностью сформировавшаяся бактериальная спора — это уплотненный участок клетки с нуклеоидом и рибосомами, ограниченный плотной многослойной оболочкой, пропитанной кальциевыми Солями дипиколиновой кислоты.
Спорообразование характерно для палочковидных бактерий — бацилл и клостридий (см. рис. 2). Различают центральное, терминальное и субтерминальное расположение спор в вегетативной части клетки, что является дифференциально-диагностическим признаком возбудителя.
В одной бактерии образуется одна спора, находящаяся в стадии покоя, при этом все процессы обмена веществ практически сведены к нулю, но сохраняется потенциальная жизнеспособность клетки. Поскольку увеличение числа микроорганизмов в этом процессе не происходит, спорообразование у бактерий не является способом размножения, а лишь приспособлением для выживания. Уникальные по степени своей устойчивости к физическим и химическим факторам бактериальные споры могут сохраняться до внешней среде без потери жизнеспособности длительное время (десятки лет), затрудняя борьбу со спороносными патогенными бактериями.
Внутриплазматические включения. Термином «включения» обозначают такие внутриклеточные структуры бактерий, которые, очевидно, не являются абсолютно необходимыми для их жизнедеятельности. Однако их природа и функции могут быть различны. В одних случаях включения являются продуктами обмена бактериальной клетки, в других — запасом питательных веществ.
Из резервных полисахаридов особенно распространены глюканы — гликоген, крахмал, гранулеза. Они выявляются в клетках бацилл, клостридий, энтеробактерий и др.
Запасные липиды представлены полиэфиром β-оксимасляной кислоты и восками. Воски, эфиры высокомолекулярных жирных кислот и спиртов характерны для микобактерий.
У коринебактерий резерв фосфора создается в виде зерен полифосфатов (волютина), имеющих диагностическое значение.
Источник: studopedia.ru
Строение бактериальной клетки
Размеры — от 1 до 15 мкм. Основные формы: 1) кокки (шаровидные), 2) бациллы (палочковидные), 3) вибрионы (изогнутые в виде запятой), 4) спириллы и спирохеты (спирально закрученные).
Бактериальная клетка ограничена оболочкой. Внутренний слой оболочки представлен цитоплазматической мембраной (1), над которой находится клеточная стенка (2); над клеточной стенкой у многих бактерий — слизистая капсула (3). Строение и функции цитоплазматической мембраны эукариотической и прокариотической клеток не отличаются. Мембрана может образовывать складки, называемые мезосомами (7). Они могут иметь разную форму (мешковидные, трубчатые, пластинчатые и др.).
На поверхности мезосом располагаются ферменты. Клеточная стенка толстая, плотная, жесткая, состоит из муреина (главный компонент) и других органических веществ. Муреин представляет собой правильную сеть из параллельных полисахаридных цепей, сшитых друг с другом короткими белковыми цепочками. В зависимости от особенностей строения клеточной стенки бактерии подразделяются на грамположительные (окрашиваются по Граму) и грамотрицательные (не окрашиваются). У грамотрицательных бактерий стенка тоньше, устроена сложнее и над муреиновым слоем снаружи имеется слой липидов. Внутреннее пространство заполнено цитоплазмой (4).
Генетический материал представлен кольцевыми молекулами ДНК. Эти ДНК можно условно разделить на «хромосомные» и плазмидные. «Хромосомная» ДНК (5) — одна, прикреплена к мембране, содержит несколько тысяч генов, в отличие от хромосомных ДНК эукариот она не линейная, не связана с белками. Зона, в которой расположена эта ДНК, называется нуклеоидом. Плазмиды — внехромосомные генетические элементы. Представляют собой небольшие кольцевые ДНК, не связаны с белками, не прикреплены к мембране, содержат небольшое число генов. Количество плазмид может быть различным. Наиболее изучены плазмиды, несущие информацию об устойчивости к лекарственным препаратам (R-фактор), принимающие участие в половом процессе (F-фактор). Плазмида, способная объединяться с хромосомой, называется эписомой.
В бактериальной клетке отсутствуют все мембранные органоиды, характерные для эукариотической клетки (митохондрии, пластиды, ЭПС, аппарат Гольджи, лизосомы).
В цитоплазме бактерий находятся рибосомы 70S-типа (6) и включения (9). Как правило, рибосомы собраны в полисомы. Каждая рибосома состоит из малой (30S) и большой субъединиц (50S). Функция рибосом: сборка полипептидной цепочки. Включения могут быть представлены глыбками крахмала, гликогена, волютина, липидными каплями.
У многих бактерий имеются жгутики (10) и пили (фимбрии) (11). Жгутики не ограничены мембраной, имеют волнистую форму и состоят из сферических субъединиц белка флагеллина. Эти субъединицы расположены по спирали и образуют полый цилиндр диаметром 10–20 нм. Жгутик прокариот по своей структуре напоминает одну из микротрубочек эукариотического жгутика. Количество и расположение жгутиков может быть различным. Пили — прямые нитевидные структуры на поверхности бактерий. Они тоньше и короче жгутиков. Представляют собой короткие полые цилиндры из белка пилина. Пили служат для прикрепления бактерий к субстрату и друг к другу. Во время конъюгации образуются особые F-пили, по которым осуществляется передача генетического материала от одной бактериальной клетки к другой.
Спорообразование у бактерий — способ переживания неблагоприятных условий. Споры формируются обычно по одной внутри «материнской клетки» и называются эндоспорами. Споры обладают высокой устойчивостью к радиации, экстремальным температурам, высушиванию и другим факторам, вызывающим гибель вегетативных клеток.
Размножение. Бактерии размножаются бесполым способом — делением «материнской клетки» надвое. Перед делением происходит репликация ДНК.
Редко у бактерий наблюдается половой процесс, при котором происходит рекомбинация генетического материала. Следует подчеркнуть, что у бактерий никогда не образуются гаметы, не происходит слияние содержимого клеток, а имеет место передача ДНК от клетки-донора к клетке-реципиенту. Различают три способа передачи ДНК: конъюгация, трансформация, трансдукция.
Конъюгация — однонаправленный перенос F-плазмиды от клетки-донора в клетку-реципиента, контактирующих друг с другом. При этом бактерии соединяются друг с другом особыми F-пилями (F-фимбриями), по каналам которых фрагменты ДНК и переносятся. Конъюгацию можно разбить на следующие этапы: 1) раскручивание F-плазмиды, 2) проникновение одной из цепей F-плазмиды в клетку-реципиента через F-пилю, 3) синтез комплементарной цепи на матрице одноцепочечной ДНК (происходит как в клетке-доноре (F+), так и в клетке-реципиенте (F—)).
Трансформация — однонаправленный перенос фрагментов ДНК от клетки-донора к клетке-реципиенту, не контактирующих друг с другом. При этом клетка-донор или «выделяет» из себя небольшой фрагмент ДНК, или ДНК попадает в окружающую среду после гибели этой клетки. В любом случае ДНК активно поглощается клеткой-реципиентом и встраивается в собственную «хромосому».
Трансдукция — перенос фрагмента ДНК от клетки-донора к клетке-реципиенту с помощью бактериофагов.
Вирусы
Вирусы открыты в 1892 г. Д.И. Ивановским при изучении мозаичной болезни табака (пятнистость листьев). Вирусы — неклеточные формы жизни. Проявляют признаки, характерные для живых организмов, только во время паразитирования в клетках других организмов. Вирусы являются внутриклеточными паразитами, но, в отличие от других паразитов, они паразитируют на генетическом уровне (ультрапаразиты). Существует несколько точек зрения на происхождение вирусов: 1) вирусы возникли в результате дегенерации клеточных организмов; 2) вирусы можно рассматривать как группу «потерявшихся», вышедших из-под контроля клетки генов («осколок генома»); 3) вирусы произошли от клеточных органоидов и др.
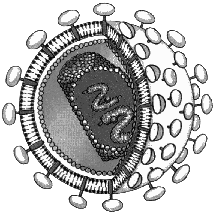
Вирусы состоят из нуклеиновой кислоты (ДНК или РНК) и белков, образующих оболочку вокруг этой нуклеиновой кислоты, т.е. представляют собой нуклеопротеидный комплекс. В состав некоторых вирусов входят липиды и углеводы. Вирусы содержат всегда один тип нуклеиновой кислоты — либо ДНК, либо РНК. Причем каждая из нуклеиновых кислот может быть как одноцепочечной, так и двухцепочечной, как линейной, так и кольцевой.
Размеры вирусов — 10–300 нм. Форма вирусов: шаровидная, палочковидная, нитевидная, цилиндрическая и др.
Капсид — оболочка вируса, образована белковыми субъединицами, уложенными определенным образом. Капсид защищает нуклеиновую кислоту вируса от различных воздействий, обеспечивает осаждение вируса на поверхности клетки-хозяина. Суперкапсид характерен для сложноорганизованных вирусов (ВИЧ, вирусы гриппа, герпеса). Возникает во время выхода вируса из клетки-хозяина и представляет собой модифицированный участок ядерной или наружной цитоплазматической мембраны клетки-хозяина.
Если вирус находится внутри клетки-хозяина, то он существует в форме нуклеиновой кислоты. Если вирус находится вне клетки-хозяина, то он представляет собой нуклеопротеидный комплекс, и эта свободная форма существования называется вирионом. Вирусы обладают высокой специфичностью, т.е. они могут использовать для своей жизнедеятельности строго определенный круг хозяев.
Вирусы, паразитирующие в бактериальных клетках, называются бактериофагами. Бактериофаг состоит из головки, хвостика и хвостовых отростков, с помощью которых он осаждается на оболочке бактерий. В головке содержится ДНК или РНК. Фаг частично растворяет клеточную стенку и мембрану бактерии и за счет сократительной реакции хвостика «впрыскивает» свою нуклеиновую кислоту в ее клетку.
Только паразитируя в клетке-хозяине, вирус может репродуцироваться, воспроизводить себе подобных.
В цикле репродукции вируса можно выделить следующие стадии.
- Осаждение на поверхности клетки-хозяина.
- Проникновение вируса в клетку-хозяина (могут попасть в клетку-хозяина путем: а) «инъекции», б) растворения оболочки клетки вирусными ферментами, в) эндоцитоза; попав внутрь клетки вирус переводит ее белок-синтезирующий аппарат под собственный контроль).
- Встраивание вирусной ДНК в ДНК клетки-хозяина (у РНК-содержащих вирусов перед этим происходит обратная транскрипция — синтез ДНК на матрице РНК).
- Транскрипция вирусной РНК.
- Синтез вирусных белков.
- Синтез вирусных нуклеиновых кислот.
- Самосборка и выход из клетки дочерних вирусов. Затем клетка либо погибает, либо продолжает существовать и производить новые поколения вирусных частиц.
Вирусы способны паразитировать в клетках большинства существующих живых организмов, вызывая различные заболевания (грипп, коревая краснуха, полиомиелит, СПИД, оспа, бешенство и др.). Возбудитель СПИДа — вирус иммунодефицита человека (ВИЧ) — относится к ретровирусам. Имеет сферическую форму, диаметром 100–150 нм. Наружная оболочка вируса состоит из мембраны, образованной из клеточной мембраны клетки-хозяина. В мембрану встроены рецепторные «грибовидные» образования. Под наружной оболочкой располагается сердцевина вируса, имеющая форму усеченного конуса и образованная особыми белками. Внутри сердцевины располагаются две молекулы вирусной РНК. Каждая молекула РНК содержит 9 генов ВИЧ и фермент (обратная транскриптаза), осуществляющий синтез вирусной ДНК на матрице вирусной РНК.
Вирус иммунодефицита человека поражает главным образом CD4-лимфоциты (хелперы), на поверхности которых есть рецепторы, способные связываться с поверхностным белком ВИЧ. Кроме того, ВИЧ проникает в клетки ЦНС, нейроглии, кишечника. Иммунная система организма человека утрачивает свои защитные свойства и оказывается не в состоянии противостоять возбудителям различных инфекций. Средняя продолжительность жизни инфицированного человека составляет 7–10 лет.
Источником заражения служит только человек — носитель вируса иммунодефицита. СПИД передается половым путем, через кровь и ткани, содержащие вирус иммунодефицита, от матери к плоду.
Источник: licey.net