Строение хромосомы эукариотической клетки рисунок
Эндоплазматическая сеть
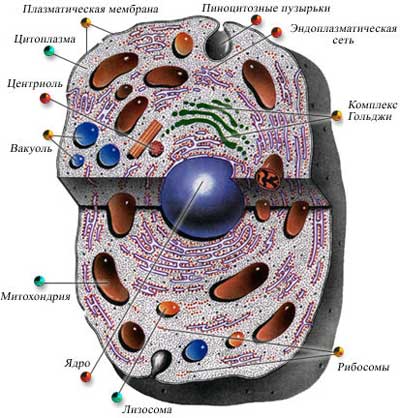
Эндоплазматическая сеть (ЭПС), или эндоплазматический ретикулум (ЭПР), — одномембранный органоид. Представляет собой систему мембран, формирующих «цистерны» и каналы, соединенных друг с другом и ограничивающих единое внутреннее пространство — полости ЭПС. Мембраны с одной стороны связаны с цитоплазматической мембраной, с другой — с наружной ядерной мембраной. Различают два вида ЭПС: 1) шероховатая (гранулярная), содержащая на своей поверхности рибосомы, и 2) гладкая (агранулярная), мембраны которой рибосом не несут.
Функции: 1) транспорт веществ из одной части клетки в другую, 2) разделение цитоплазмы клетки на компартменты ( «отсеки»), 3) синтез углеводов и липидов (гладкая ЭПС), 4) синтез белка (шероховатая ЭПС), 5) место образования аппарата Гольджи.
Аппарат Гольджи
Аппарат Гольджи, или комплекс Гольджи, — одномембранный органоид. Представляет собой стопки уплощенных «цистерн» с расширенными краями. С ними связана система мелких одномембранных пузырьков (пузырьки Гольджи). Каждая стопка обычно состоит из 4-х–6-ти «цистерн», является структурно-функциональной единицей аппарата Гольджи и называется диктиосомой. Число диктиосом в клетке колеблется от одной до нескольких сотен. В растительных клетках диктиосомы обособлены.

Аппарат Гольджи обычно расположен около клеточного ядра (в животных клетках часто вблизи клеточного центра).
Функции аппарата Гольджи: 1) накопление белков, липидов, углеводов, 2) модификация поступивших органических веществ, 3) «упаковка» в мембранные пузырьки белков, липидов, углеводов, 4) секреция белков, липидов, углеводов, 5) синтез углеводов и липидов, 6) место образования лизосом. Секреторная функция является важнейшей, поэтому аппарат Гольджи хорошо развит в секреторных клетках.
Лизосомы
Лизосомы — одномембранные органоиды. Представляют собой мелкие пузырьки (диаметр от 0,2 до 0,8 мкм), содержащие набор гидролитических ферментов. Ферменты синтезируются на шероховатой ЭПС, перемещаются в аппарат Гольджи, где происходит их модификация и упаковка в мембранные пузырьки, которые после отделения от аппарата Гольджи становятся собственно лизосомами. Лизосома может содержать от 20 до 60 различных видов гидролитических ферментов. Расщепление веществ с помощью ферментов называют лизисом.
Различают: 1) первичные лизосомы, 2) вторичные лизосомы. Первичными называются лизосомы, отшнуровавшиеся от аппарата Гольджи. Первичные лизосомы являются фактором, обеспечивающим экзоцитоз ферментов из клетки.
Вторичными называются лизосомы, образовавшиеся в результате слияния первичных лизосом с эндоцитозными вакуолями. В этом случае в них происходит переваривание веществ, поступивших в клетку путем фагоцитоза или пиноцитоза, поэтому их можно назвать пищеварительными вакуолями.
Автофагия — процесс уничтожения ненужных клетке структур. Сначала подлежащая уничтожению структура окружается одинарной мембраной, затем образовавшаяся мембранная капсула сливается с первичной лизосомой, в результате также образуется вторичная лизосома (автофагическая вакуоль), в которой эта структура переваривается. Продукты переваривания усваиваются цитоплазмой клетки, но часть материала так и остается непереваренной. Вторичная лизосома, содержащая этот непереваренный материал, называется остаточным тельцем. Путем экзоцитоза непереваренные частицы удаляются из клетки.
Автолиз — саморазрушение клетки, наступающее вследствие высвобождения содержимого лизосом. В норме автолиз имеет место при метаморфозах (исчезновение хвоста у головастика лягушек), инволюции матки после родов, в очагах омертвления тканей.
Функции лизосом: 1) внутриклеточное переваривание органических веществ, 2) уничтожение ненужных клеточных и неклеточных структур, 3) участие в процессах реорганизации клеток.
Вакуоли
Вакуоли — одномембранные органоиды, представляют собой «емкости», заполненные водными растворами органических и неорганических веществ. В образовании вакуолей принимают участие ЭПС и аппарат Гольджи. Молодые растительные клетки содержат много мелких вакуолей, которые затем по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну большую центральную вакуоль. Центральная вакуоль может занимать до 95% объема зрелой клетки, ядро и органоиды оттесняются при этом к клеточной оболочке. Мембрана, ограничивающая растительную вакуоль, называется тонопластом. Жидкость, заполняющая растительную вакуоль, называется клеточным соком. В состав клеточного сока входят водорастворимые органические и неорганические соли, моносахариды, дисахариды, аминокислоты, конечные или токсические продукты обмена веществ (гликозиды, алкалоиды), некоторые пигменты (антоцианы).
В животных клетках имеются мелкие пищеварительные и автофагические вакуоли, относящиеся к группе вторичных лизосом и содержащие гидролитические ферменты. У одноклеточных животных есть еще сократительные вакуоли, выполняющие функцию осморегуляции и выделения.
Функции вакуоли: 1) накопление и хранение воды, 2) регуляция водно-солевого обмена, 3) поддержание тургорного давления, 4) накопление водорастворимых метаболитов, запасных питательных веществ, 5) окрашивание цветов и плодов и привлечение тем самым опылителей и распространителей семян, 6) см. функции лизосом.
Эндоплазматическая сеть, аппарат Гольджи, лизосомы и вакуоли образуют единую вакуолярную сеть клетки, отдельные элементы которой могут переходить друг в друга.
Митохондрии
Форма, размеры и количество митохондрий чрезвычайно варьируют. По форме митохондрии могут быть палочковидными, округлыми, спиральными, чашевидными, разветвленными. Длина митохондрий колеблется в пределах от 1,5 до 10 мкм, диаметр — от 0,25 до 1,00 мкм. Количество митохондрий в клетке может достигать нескольких тысяч и зависит от метаболической активности клетки.
Митохондрия ограничена двумя мембранами. Наружная мембрана митохондрий (1) гладкая, внутренняя (2) образует многочисленные складки — кристы (4). Кристы увеличивают площадь поверхности внутренней мембраны, на которой размещаются мультиферментные системы (5), участвующие в процессах синтеза молекул АТФ. Внутреннее пространство митохондрий заполнено матриксом (3). В матриксе содержатся кольцевая ДНК (6), специфические иРНК, рибосомы прокариотического типа (70S-типа), ферменты цикла Кребса.
Митохондриальная ДНК не связана с белками («голая»), прикреплена к внутренней мембране митохондрии и несет информацию о строении примерно 30 белков.
я построения митохондрии требуется гораздо больше белков, поэтому информация о большинстве митохондриальных белков содержится в ядерной ДНК, и эти белки синтезируются в цитоплазме клетки. Митохондрии способны автономно размножаться путем деления надвое. Между наружной и внутренней мембранами находится протонный резервуар, где происходит накопление Н+.
Функции митохондрий: 1) синтез АТФ, 2) кислородное расщепление органических веществ.
Согласно одной из гипотез (теория симбиогенеза) митохондрии произошли от древних свободноживущих аэробных прокариотических организмов, которые, случайно проникнув в клетку-хозяина, затем образовали с ней взаимовыгодный симбиотический комплекс. В пользу этой гипотезы свидетельствуют следующие данные. Во-первых, митохондриальная ДНК имеет такие же особенности строения как и ДНК современных бактерий (замкнута в кольцо, не связана с белками). Во-вторых, митохондриальные рибосомы и рибосомы бактерий относятся к одному типу — 70S-типу. В-третьих, механизм деления митохондрий сходен с таковым бактерий. В-четвертых, синтез митохондриальных и бактериальных белков подавляется одинаковыми антибиотиками.
Пластиды
Пластиды характерны только для растительных клеток. Различают три основных типа пластид: лейкопласты — бесцветные пластиды в клетках неокрашенных частей растений, хромопласты — окрашенные пластиды обычно желтого, красного и оранжевого цветов, хлоропласты — зеленые пластиды.
Хлоропласты. В клетках высших растений хлоропласты имеют форму двояковыпуклой линзы. Длина хлоропластов колеблется в пределах от 5 до 10 мкм, диаметр — от 2 до 4 мкм. Хлоропласты ограничены двумя мембранами. Наружная мембрана (1) гладкая, внутренняя (2) имеет сложную складчатую структуру. Наименьшая складка называется тилакоидом (4). Группа тилакоидов, уложенных наподобие стопки монет, называется граной (5). В хлоропласте содержится в среднем 40–60 гран, расположенных в шахматном порядке. Граны связываются друг с другом уплощенными каналами — ламеллами (6). В мембраны тилакоидов встроены фотосинтетические пигменты и ферменты, обеспечивающие синтез АТФ. Главным фотосинтетическим пигментом является хлорофилл, который и обусловливает зеленый цвет хлоропластов.
Внутреннее пространство хлоропластов заполнено стромой (3). В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты цикла Кальвина, зерна крахмала (7). Внутри каждого тилакоида находится протонный резервуар, происходит накопление Н+. Хлоропласты, также как митохондрии, способны к автономному размножению путем деления надвое. Они содержатся в клетках зеленых частей высших растений, особенно много хлоропластов в листьях и зеленых плодах. Хлоропласты низших растений называют хроматофорами.
Функция хлоропластов: фотосинтез. Полагают, что хлоропласты произошли от древних эндосимбиотических цианобактерий (теория симбиогенеза). Основанием для такого предположения является сходство хлоропластов и современных бактерий по ряду признаков (кольцевая, «голая» ДНК, рибосомы 70S-типа, способ размножения).
Лейкопласты. Форма варьирует (шаровидные, округлые, чашевидные и др.). Лейкопласты ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя образует малочисленные тилакоиды. В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты синтеза и гидролиза запасных питательных веществ. Пигменты отсутствуют. Особенно много лейкопластов имеют клетки подземных органов растения (корни, клубни, корневища и др.). Функция лейкопластов: синтез, накопление и хранение запасных питательных веществ. Амилопласты — лейкопласты, которые синтезируют и накапливают крахмал, элайопласты — масла, протеинопласты — белки. В одном и том же лейкопласте могут накапливаться разные вещества.
Хромопласты. Ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя или также гладкая, или образует единичные тилакоиды. В строме имеются кольцевая ДНК и пигменты — каротиноиды, придающие хромопластам желтую, красную или оранжевую окраску. Форма накопления пигментов различная: в виде кристаллов, растворены в липидных каплях (8) и др. Содержатся в клетках зрелых плодов, лепестков, осенних листьев, редко — корнеплодов. Хромопласты считаются конечной стадией развития пластид.
Функция хромопластов: окрашивание цветов и плодов и тем самым привлечение опылителей и распространителей семян.
Все виды пластид могут образовываться из пропластид. Пропластиды — мелкие органоиды, содержащиеся в меристематических тканях. Поскольку пластиды имеют общее происхождение, между ними возможны взаимопревращения. Лейкопласты могут превращаться в хлоропласты (позеленение клубней картофеля на свету), хлоропласты — в хромопласты (пожелтение листьев и покраснение плодов). Превращение хромопластов в лейкопласты или хлоропласты считается невозможным.
Рибосомы
Рибосомы — немембранные органоиды, диаметр примерно 20 нм. Рибосомы состоят из двух субъединиц — большой и малой, на которые могут диссоциировать. Химический состав рибосом — белки и рРНК. Молекулы рРНК составляют 50–63% массы рибосомы и образуют ее структурный каркас. Различают два типа рибосом: 1) эукариотические (с константами седиментации целой рибосомы — 80S, малой субъединицы — 40S, большой — 60S) и 2) прокариотические (соответственно 70S, 30S, 50S).
В составе рибосом эукариотического типа 4 молекулы рРНК и около 100 молекул белка, прокариотического типа — 3 молекулы рРНК и около 55 молекул белка.
время биосинтеза белка рибосомы могут «работать» поодиночке или объединяться в комплексы — полирибосомы (полисомы). В таких комплексах они связаны друг с другом одной молекулой иРНК. Прокариотические клетки имеют рибосомы только 70S-типа. Эукариотические клетки имеют рибосомы как 80S-типа (шероховатые мембраны ЭПС, цитоплазма), так и 70S-типа (митохондрии, хлоропласты).
Субъединицы рибосомы эукариот образуются в ядрышке. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка.
Функция рибосом: сборка полипептидной цепочки (синтез белка).
Цитоскелет
Цитоскелет образован микротрубочками и микрофиламентами. Микротрубочки — цилиндрические неразветвленные структуры. Длина микротрубочек колеблется от 100 мкм до 1 мм, диаметр составляет примерно 24 нм, толщина стенки — 5 нм. Основной химический компонент — белок тубулин. Микротрубочки разрушаются под воздействием колхицина. Микрофиламенты — нити диаметром 5–7 нм, состоят из белка актина. Микротрубочки и микрофиламенты образуют в цитоплазме сложные переплетения. Функции цитоскелета: 1) определение формы клетки, 2) опора для органоидов, 3) образование веретена деления, 4) участие в движениях клетки, 5) организация тока цитоплазмы.
Клеточный центр
Клеточный центр включает в себя две центриоли и центросферу. Центриоль представляет собой цилиндр, стенка которого образована девятью группами из трех слившихся микротрубочек (9 триплетов), соединенных между собой через определенные интервалы поперечными сшивками. Центриоли объединены в пары, где они расположены под прямым углом друг к другу. Перед делением клетки центриоли расходятся к противоположным полюсам, и возле каждой из них возникает дочерняя центриоль. Они формируют веретено деления, способствующее равномерному распределению генетического материала между дочерними клетками. В клетках высших растений (голосеменные, покрытосеменные) клеточный центр центриолей не имеет. Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы, они возникают в результате дупликации уже имеющихся центриолей. Функции: 1) обеспечение расхождения хромосом к полюсам клетки во время митоза или мейоза, 2) центр организации цитоскелета.
Органоиды движения
Присутствуют не во всех клетках. К органоидам движения относятся реснички (инфузории, эпителий дыхательных путей), жгутики (жгутиконосцы, сперматозоиды), ложноножки (корненожки, лейкоциты), миофибриллы (мышечные клетки) и др.
Жгутики и реснички — органоиды нитевидной формы, представляют собой аксонему, ограниченную мембраной. Аксонема — цилиндрическая структура; стенка цилиндра образована девятью парами микротрубочек, в его центре находятся две одиночные микротрубочки. В основании аксонемы находятся базальные тельца, представленные двумя взаимно перпендикулярными центриолями (каждое базальное тельце состоит из девяти триплетов микротрубочек, в его центре микротрубочек нет). Длина жгутика достигает 150 мкм, реснички в несколько раз короче.
Миофибриллы состоят из актиновых и миозиновых миофиламентов, обеспечивающих сокращение мышечных клеток.
Источник: licey.net
Строение хромосом на микроскопическом уровне
Хромосомы, как отдельные структуры, становятся доступными для исследования только после значительной конденсации хроматина, которая наступает во время митоза (в соматических клетках), либо во время мейоза (при образовании половых клеток). Начавшаяся в профазе конденсация хроматина заканчивается в метафазе, поэтому, как правило, хромосомы изучаются на стадии метафазной пластинки.
В интерфазе хромосомы находятся в деконденсированном состоянии, и определить их как отдельные структуры не представляется возможным.
В метафазе каждая хромосома имеет как бы иксобразную форму и состоит из двух идентичных половин — хроматид (сестринских хромосом), тесно прилежащих друг к другу только в области первичной перетяжки (центромеры), а на остальном протяжении между хроматидами видна большая щель. Центромера — это тот участок, где хромосома находится в деконденсированном состоянии, и к ней прикрепляются нити веретена деления. Центромера делит хромосомы на плечи. По положению центромеры выделяют три вида хромосом.
1. Метацентрические, у которых плечи имеют примерно одинаковую длину (т.е. центромера расположена посередине хромосомы).
2. Субметацентрические, у которых центромера смещена от середины, располагается субмедиально и делит хромосому на два плеча неравной длины. Верхнее всегда меньшее.
З. Акроцентрические, у которых центромера расположена почти на конце хромосомы, отделяя от длинного плеча очень короткое верхнее плечо.
Верхние короткие плечи принято обозначать буквой «р», а нижние длинные буквой «q«. Характерной чертой для некоторых хромосом является наличие вторичных перетяжек, они возникают в участках неполной конденсации хромосом и располагаются в околоцентромерных участках 1-й, 9-й и 16-й хромосом. Вторичные перетяжки имеются также в 13-15 и 21 -22-й хромосомах, однако здесь они занимают удаленное от центромеры положение, отделяя небольшой концевой участок короткого плеча хромосом в виде спутника. Эти хромосомы называют спутничными. В этих хромосомах в области вторичной перетяжки сосредоточены гены, кодирующие р-РНК, и в прилежащих участках кариоплазмы образуются ядрышки. Поэтому такого рода вторичные перетяжки называют ядрышковыми организаторами. В хромосомных наборах одних людей указанные хромосомы имеют вторичную перетяжку, а в этих же хромосомах у других людей её может не быть.
Источник: StudFiles.net
Рубрики
Источник: kaz-ekzams.ru
| Эукариотическая клетка. Строение эукариотической клетки |
|
Строение эукариотической клетки. Плазмалемма (клеточная оболочка) животных клеток образована мембраной, покрытой снаружи слоем гликокаликса толщиной 10-20 нм. Плазмалемма выполняет отграничивающую, барьерную, транспортную и рецепторную функции. Благодаря свойству избирательной проницаемости плазмалемма регулирует химический состав внутренней среды клетки. В плазмалемме размещены молекулы рецепторов, которые избирательно распознают определенные биологически активные вещества (гормоны). В пластах и слоях соседние клетки удерживаются благодаря наличию разного вида контактов, которые представлены участками плазмалеммы, имеющими особое строение. Изнутри к мембране примыкает кортикальный (корковый) слой цитоплазмы толщиной 0,1—0,5 мкм. Цитоплазма. В цитоплазме находится целый ряд оформленных структур, имеющих закономерные особенности строения и поведения в разные периоды жизнедеятельности клетки. Каждая из этих структур несёт определенную функцию. Отсюда возникло сопоставление их с органами целого организма, в связи с чем они получили название органеллы, или органоиды. В цитоплазме откладываются различные вещества — включения (гликоген, капли жира, пигменты). Цитоплазма пронизана мембранами эндоплазматической сети. Эндоплазматическая сеть (ЭДС). Эндоплазматическая сеть — это разветвлённая сеть каналов и полостей в цитоплазме клетки, образованная мембранами. На мембранах каналов находятся многочисленные ферменты, обеспечивающие жизнедеятельность клетки. Различают 2 вида мембран ЭДС — гладкие и шероховатые. На мембранах гладкой эндоплазматической сети находятся ферментные системы, участвующие в жировом и углеводном обмене. Основная функция шероховатой эндоплазматической сети — синтез белков, который осуществляется в рибосомах, прикрепленных к мембранам. Эндоплазматическая сеть — это общая внутриклеточная циркуляционная система, по каналам которой транспортируются вещества внутри клетки и из клетки в клетку. Рибосомы осуществляют функцию синтеза белков. Рибосомы представляют собой сферические частицы диаметром 15-35нм, состоящие из 2 субъединиц неравных размеров и содержащие примерно равное количество белков и РНК. Рибосомы в цитоплазме располагаются или прикрепляются к наружной поверхности мембран эндоплазматической сети. В зависимости от типа синтезируемого белка рибосомы могут объединяться в комплексы — полирибосомы. Рибосомы присутствуют во всех типах клеток. Комплекс Гольджи. Основным структурным элементом комплекса Гольджи является гладкая мембрана, которая образует пакеты уплощенных цистерн, или крупные вакуоли, или мелкие пузырьки. Цистерны комплекса Гольджи соединены с каналами эндоплазматической сети. Синтезированные на мембранах эндоплазматической сети белки, полисахариды, жиры транспортируются к комплексу, конденсируются внутри его структур и «упаковываются» в виде секрета, готового к выделению, либо используются в самой клетке в процессе её жизнедеятельности. Митохондрии. Всеобщее распространение митохондрий в животном и растительном мире указывают на важную роль, которую митохондрии играют в клетке. Митохондрии имеют форму сферических, овальных и цилиндрических телец, могут быть нитевидной формы. Размеры митохондрий 0,2-1мкм в диаметре, до 5-7мкм в длину. Длина нитевидных форм достигает 15-20мкм. Количество митохондрий в клетках различных тканей неодинаково, их больше там, где интенсивны синтетические процессы (печень) или велики затраты энергии. Стенка митохондрий состоит из 2-х мембран — наружной и внутренней. Наружная мембрана гладкая, а от внутренней внутрь органоида отходят перегородки — гребни, или кристы. На мембранах крист находятся многочисленные ферменты, участвующие в энергетическом обмене. Основная функция митохондрий — синтез АТФ. Лизосомы — небольшие овальные тельца диаметром около 0,4мкм, окруженные одной трехслойной мембраной. В лизосомах находится около 30 ферментов, способных расщеплять белки, нуклеиновые кислоты, полисахариды, липиды и др. вещества. Расщепление веществ с помощью ферментов называется лизисом, поэтому и органоид назван лизосомой. Полагают, что лизосомы образуются из структур комплекса Гольджи либо непосредственно из эндоплазматической сети. Функции лизосом: внутриклеточное переваривание пищевых веществ, разрушение структуры самой клетки при её отмирании в ходе эмбрионального развития, когда происходит замена зародышевых тканей на постоянные, и в ряде других случаев. Центриоли. Клеточный центр состоит из 2-х очень маленьких телец цилиндрической формы, расположенных под прямым углом друг к другу. Эти тельца называются центриолями. Стенка центриоли состоит из 9-ти пар микротрубочек. Центриоли способны к самосборке и относятся к самовоспроизводящимся органоидам цитоплазмы. Центриоли играют важную роль в клеточном делении: от них начинается рост микротрубочек, образующих веретено деления. Ядро. Ядро — важнейшая составная часть клетки. Оно содержит молекулы ДНК и поэтому выполняет две главные функции: 1) хранение и воспроизведение генетической информации, 2) регуляция процессов обмена веществ, протекающих в клетке. Клетка утратившая ядро, не может существовать. Ядро также неспособно к самостоятельному существованию. Большинство клеток имеет одно ядро, но можно наблюдать 2-3ядра в одной клетке, например в клетках печени. Известны многоядерные клетки с числом ядер в несколько десятков. Формы ядер зависят от формы клетки. Ядра бывают шаровидные, многолопастные. Ядро окружено оболочкой, состоящей из двух мембран, имеющих обычное трёхслойное строение. Наружная ядерная мембрана покрыта рибосомами, внутренняя мембрана гладкая. Главную роль в жизнедеятельности ядра играет обмен веществ между ядром и цитоплазмой. Содержимое ядра включает ядерный сок, или кариоплазму, хроматин и ядрышко. В состав ядерного сока входят различные белки, в том числе большинство ферментов ядра, свободные нуклеотиды, аминокислоты, продукты деятельности ядрышка и хроматина, перемещающиеся из ядра в цитоплазму. Хроматин содержит ДНК, белки и представляет собой спирализованные и уплотненные участки хромосом. Ядрышко представляет собой плотное округлое тельце, располагающееся в ядерном соке. Число ядрышек колеблется от 1 до 5-7 и более. Ядрышки есть только в неделящихся ядрах, во время митоза они исчезают, а после завершения деление образуются вновь. Ядрышко не является самостоятельным органоидом клетки, оно лишено мембраны и образуется вокруг участка хромосомы, в котором закодирована структура рРНК. В ядрышке формируются рибосомы, которые затем перемещаются в цитоплазму. Хроматином называют глыбки, гранулы и сетевидные структуры ядра, интенсивно окрашивающиеся некоторыми красителями и отличные по форме от ядрышка. Читайте по теме: Хромосомы. |
Источник: mirbiologa.ru
Ядро. Строение и функции.
В клетке выделяют ядро и цитоплазму. Клеточное ядросостоит из оболочки, ядерного сока, ядрышка и хроматина. Функциональная рольядерной оболочкизаключается в обособлении генетического материала (хромосом) эукариотической клетки от цитоплазмы с присущими ей многочисленными метаболическими реакциями, а также регуляции двусторонних взаимодействий ядра и цитоплазмы. Ядерная оболочка состоит из двух мембран, разделенных околоядерным (перинуклеарным) пространством. Последнее может сообщаться с канальцами цитоплазматической сети.
Ядерная оболочка пронизана порожу диаметром 80—90нм. Область поры или поровый комплекс с диаметром около 120нм имеет определенное строение, что указывает на сложный механизм регуляции ядерно-цитоплазматических перемещений веществ и структур. Количество пор зависит от функционального состояния клетки. Чем выше синтетическая активность в клетке, тем больше их число. Подсчитано, что у низших позвоночных животных в эритробластах, где интенсивно образуется и накапливается гемоглобин, на 1мкм2ядерной оболочки приходится около 30пор. В зрелых эритроцитах названных животных, сохраняющих ядра, на 1мк»г оболочки остается до пяти пор, т.е. в 6 раз меньше.
В области перового комплекса начинается так называемая плотная пластинка —белковый слой, подстилающий на всем протяжении внутреннюю мембрану ядерной оболочки. Эта структура выполняет прежде всего опорную функцию, так как при ее наличии форма ядра сохраняется даже в случае разрушения обеих мембран ядерной оболочки. Предполагают также, что закономерная связь с веществом плотной пластинки способствует упорядоченному расположению хромосом в интерфазном ядре.
Основу ядерного сока,илиматрикса,составляют белки. Ядерный сок образует внутреннюю среду ядра, в связи с чем он играет важную роль в обеспечении нормального функционирования генетического материала. В составе ядерного сока присутствуютнитчатые,илифибриллярные, белки, с которыми связано выполнение опорной функции: в матриксе находятся также первичные продукты транскрипции генетической информации — гетероядерные РНК (гя-РНК), которые здесь же подвергаются процессингу, превращаясь в м-РНК (см. 3.4.3.2).
Ядрышкопредставляет собой структуру, в которой происходит образование и созреваниерибосомальныхРНК (рРНК). Гены рРНК занимают определенные участки (в зависимости от вида животного) одной или нескольких хромосом (у человека 13—15и 21—22пары) — ядрышковые организаторы, в области которых и образуются ядрышки. Такие участки в метафазных хромосомах выглядят как сужения и называютсявторичными перетяжками. Спомощью электронного микроскопа в ядрышке выявляют нитчатый и зернистый компоненты. Нитчатый (фибриллярный) компонент представлен комплексами белка и гигантских молекул РНК-предшественниц, из которых затем образуются более мелкие молекулы зрелых рРНК. В процессе созревания фибриллы преобразуются в рибонуклеопротеиновые зерна (гранулы), которыми представлен зернистый компонент.
Хроматиновые структуры в виде глыбок,рассеянных в нуклеоплазме, являются интерфазной формой существования хромосом клетки
цитоплазма
В цитоплазмеразличают основное вещество (матрикс, гиалоплазма), включения и органеллы.Основное вещество цитоплазмызаполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Обычный электронный микроскоп не выявляет в нем какой-либо внутренней организации. Белковый состав гиалоплазмы разнообразен. Важнейшие из белков представлены ферментами гаиколиза, обмена Сахаров, азотистых оснований, аминокислот и липидов. Ряд белков гиалоплазмы служит субъединицами, из которых происходит сборка таких структур, как микротрубочки.
Основное вещество цитоплазмы образует истинную внутреннюю среду клетки, которая объединяет все внутриклеточные структуры и обеспечивает взаимодействие их друг с другом. Выполнение матриксом объединяющей, а также каркасной функции может быть связано с выявляемой с помощью сверхмощного электронного микроскопа микротрабекулярной сети, образованной тонкими фибриллами толщиной 2—3нм и пронизывающей всю цитоплазму. Через гиалоплазму осуществляется значительный объем внутриклеточных перемещений веществ и структур. Основное вещество цитоплазмы следует рассматривать так же, как сложную коллоидную систему, способную переходить из золеобразного (жидкого) состояния в гелеобразное. В процессе таких переходов совершается работа. О функциональном значении таких переходов см. разд. 2.3.8.
Включениями(рис. 2.5)называют относительно непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген), продуктами, подлежащими выведению из клетки (гранулы секрета), балластными веществами (некоторые пигменты).
Органеллы —это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции.
Выделяют органеллы общего значенияиспециальные.Последние в значительном количестве присутствуют в клетках, специализированных к выполнению определенной функции, но в незначительном количестве могут встречаться и в других типах клеток. К ним относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки, транспортирующие вещества —переносчики нервного возбуждения с одной нервной клетки на другую или клетку рабочего органа, миофибриллы, от которых зависит сокращение мышцы. Детальное рассмотрение специальных органелл входит в задачу курса гистологии.
К органеллам общего значения относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра. В растительных клетках выделяют также хлоропласта, в которых происходит фотосинтез.
Канальцеваяивакуолярная системыобразованы сообщающимися или отдельными трубчатыми или уплощенными (цистерна) полостями, ограниченными мембранами и распространяющимися по всей цитоплазме клетки. Нередко цистерны имеют пузыревидные расширения. В названной системе выделяютшероховатуюигладкую цитоплазматическую сети(см. рис. 2.3).Особенность строения шероховатой сети состоит в прикреплении к ее мембранам полисом. В силу этого она выполняет функцию синтеза определенной категории белков, преимущественно удаляемых из клетки, например секретируемых клетками желез. В области шероховатой сети происходит образование белков и липидов цитоплазматических мембран, а также их сборка. Плотно упакованные в слоистую структуру цистерны шероховатой сети являются участками наиболее активного белкового синтеза и называютсяэргастоплазмой.
Мембраны гладкой цитоплазматической сети лишены полисом. Функционально эта сеть связана с обменом углеводов, жиров и других веществ небелковой природы, например стероидных гормонов (в половых железах, корковом слое надпочечников). По канальцам и цистернам происходит перемещение веществ, в частности секретируемого железистой клеткой материала, от места синтеза в зону упаковки в гранулы. В участках печеночных клеток, богатых структурами гладкой сети, разрушаются и обезвреживаются вредные токсические вещества, некоторые лекарства (барбитураты). В пузырьках и канальцах гладкой сети поперечно-полосатой мускулатуры сохраняются (депонируются) ионы кальция, играющие важную роль в процессе сокращения.
Рибосома —это округлая рибонуклеопротеиновая частица диаметром 20—30нм. Она состоит из малой и большой субъединиц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называютполисомой.Полисомы свободно располагаются в основном веществе цитоплазмы или прикреплены к мембранам шероховатой цитоплазматической сети. В обоих случаях они служат местом активного синтеза белка. Сравнение соотношения количества свободных и прикрепленных к мембранам полисом в эмбриональных недифференцированных и опухолевых клетках, с одной стороны, и в специализированных клетках взрослого организма —с другой, привело к заключению, что на полисомах гиалоплазмы образуются белки для собственных нужд (для «домашнего» пользования) данной клетки, тогда как на полисомах гранулярной сети синтезируются белки, выводимые из клетки и используемые на нужды организма (например, пищеварительные ферменты, белки грудного молока).
Пластинчатый комплекс Голъджиобразован совокупностью диктиосом числом от нескольких десятков (обычно около 20)до нескольких сотен и даже тысяч на клетку.
Диктиосома(рис. 2.6,А) представлена стопкой из 3—12уплощенных дискообразных цистерн, от краев которых отшнуровываются пузырьки (везикулы). Ограниченные определенным участком (локальные) расширения цистерн дают более крупные пузырьки (вакуоли). В дифференцированных клетках позвоночных животных и человека диктиосомы обычно собраны в околоядерной зоне цитоплазмы. В пластинчатом комплексе образуются секреторные пузырьки или вакуоли, содержимое которых составляют белки и другие соединения, подлежащие выводу из клетки. При этом предшественник секрета (просекрет), поступающий.в диктиосому из зоны синтеза, подвергается в ней некоторым химическим преобразованиям. Он также обособляется (сегрегируется) в виде «порций», которые здесь же одеваются мембранной оболочкой. В пластинчатом комплексе образуются лизосомы. В диктиосомах синтезируются полисахариды, а также их комплексы с белками (гликопротеины) и жирами (гликолипиды), которые затем можно обнаружить в гликокаликсе клеточной оболочки.
Митохондрии(рис. 2.6, Б) —это структуры округлой или палочко-видной, нередко ветвящейся формы толщиной 0,5мкм и длиной обычно до 5—10мкм. В большинстве животных клеток количество митохондрий колеблется от 150до 1500,однако в женских половых клетках их число достигает нескольких сотен тысяч. В сперматозоидах нередко присутствует одна гигантская митохондрия, спирально закрученная вокруг осевой части жгутика. Одна разветвленная митохондрия обнаружена в клетке такого паразита человека, как трипаносома.
Оболочка митохондрий состоит из двух мембран, различающихся по химическому составу, набору ферментов и функциям. Внутренняя мембрана образует впячивания листовидной (кристы) или трубчатой (тубулы) формы. Пространство, ограниченное внутренней мембраной, составляет матриксорганеллы. В нем с помощью электронного микроскопа обнаруживаются зерна диаметром 20—40нм. Они накапливают ионы кальция и магния, а также полисахариды, например гликоген.
В матриксе размещен собственный аппарат биосинтеза белка органеллы. Он представлен 2—б копиями кольцевой и лишенной гистонов (как у прокариот) молекулы ДНК, рибосомами, набором транспортных РНК (тРНК), ферментами редупликации ДНК, транскрипции и трансляции наследственной информации. По основным свойствам: размерам и структуре рибосом, организации собственного наследственного материала —этот аппарат сходен с таковым у прокариот и отличается от аппарата биосинтеза белка цитоплазмы эукариотической клетки (чем подтверждается симбиотическая гипотеза происхождения митохондрий; см. § 1.5).Гены собственной ДНК кодируют нуклеотидные последовательности митохондриальных рРНК и тРНК, а также последовательности аминонокислот некоторых белков органеллы, главным образом ее внутренней мембраны. Аминокислотные последовательности (первичная структура) большинства белков митохондрий закодированы в ДНК клеточного ядра и образуются вне органеллы в цитоплазме.
Главная функция митохондрий состоит в ферментативном извлечении из определенных химических веществ энергии (путем их окисления) и накоплении энергии в биологически используемой форме (путем синтеза молекул аденозинтрифосфата —АТФ). В целом этот процесс называетсяокислительным (расформированием.В энергетической функции митохондрий активно участвуют компоненты матрикса и внутренняя мембрана. Именно с этой мембраной связаны цепь переноса электронов (окисление) и АТФ-синтетаза, катализирующая сопряженное с окислением фосфорилирование АДФ в АТФ. Среди побочных функций митохондрий можно назвать участие в синтезе стероидных гормонов и некоторых аминокислот (глутаминовая).
Лизосомы(рис. 2.6,В) представляют собой пузырьки диаметром обычно 0,2—0,4мкм, которые содержат набор ферментов кислых гидролаз, катализирующих при низких значениях рН гидролитическое (в водной среде) расщепление нуклеиновых кислот, белков, жиров, полисахаридов. Их оболочка образована одинарной мембраной, покрытой иногда снаружи волокнистым белковым слоем (на электронограммах «окаймленные» пузырьки). Функция лизосом — внутриклеточное переваривание оазличных химических соединений и структур.
Первичными лизосомами(диаметр 100нм) называют неактивные органеллы,вторичными —органеллы, в которых происходит процесс переваривания. Вторичные лизосомы образуются из первичных. Они подразделяются нагетеролизосомы(фаголизосомы) иаутолизосомы (цитолизосомы). В первых (рис. 2.6,Г) переваривается материал, поступающий в клетку извне путем пиноцитоза и фагоцитоза, во вторых разрушаются собственные структуры клетки, завершившие свою функцию. Вторичные лизосомы, в которых процесс переваривания завершен, называютостаточными тельцами(телолизосомы). В них отсутствуют гидролазы и содержится непереваренный материал.
Микротельца составляют сборную группу органелл. Это ограниченные одной мембраной пузырьки диаметром 0,1—1,5мкм с мелкозернистым матриксом и нередко кристаллоидными или аморфными белковыми включениями. К этой группе относят, в частности,пероксисомы.Они содержат ферменты оксидазы, катализирующие образование пероксида водорода, который, будучи токсичным, разрушается затем под действием фермента пероксидазы. Эти реакции включены в различные метаболические циклы, например в обмен мочевой кислоты в клетках печени и почек. В печеночной клетке число пероксисом достигает70—100.
К органеллам общего значения относят также некоторые постоянные структуры цитоплазмы, лишенные мембран. Микротрубочки(рис.2.6,Д) —трубчатые образования различной длины с внешним диаметром 24нм, шириной просвета 15нм и толщиной стенки около 5нм. Встречаются в свободном состоянии в цитоплазме клеток или как структурные элементы жгутиков, ресничек, митотического веретена, центриолей. Свободные микротрубочки и микротрубочки ресничек, жгутиков и центриолей имеют разную устойчивость к разрушающим воздействиям, например химическим (колхицин). Микротрубочки строятся из стереотипных субьединиц белковой природы путем их полимеризации. В живой клетке процессы полимеризации протекают одновременно с процессами деполимеризации. Соотношением этих процессов определяется количество микротрубочек. В свободном состоянии микротрубочки выполняют опорную функцию, определяя форму клеток, а также являются факторами направленного перемещения внутриклеточных компонентов.
Микрофиламентами(рис. 2.6,Е) называют длинные, тонкие образования, иногда образующие пучки и обнаруживаемые по всей цитоплазме. Существует несколько разных типов микрофиламентов.Актиновые микрофиламентыблагодаря присутствию в них сократимых белков (актин) рассматривают в качестве структур, обеспечивающих клеточные формы движения, например амебоидные. Им приписывают также каркасную роль и участие в организации внутриклеточных перемещений органелл и участков гиалоплазмы.
По периферии клеток под плазмалеммой, а также в околоядерной зоне обнаруживаются пучки микрофиламентов толщиной 10нм — промежуточные филстенты.В эпителиальных, нервных, глиальных, мышечных клетках, фибробластах они построены из разных белков. Промежуточные филаменты выполняют, по-видимому, механическую, каркасную функцию.
Актиновые микрофибриллы и промежуточные филаменты, как и микротрубочки, построены из субъединиц. В силу этого их количество зависит от соотношения процессов полимеризации и деполимеризации.
Для животных клеток, части клеток растений, грибов и водорослей характерен клеточный центр,в состав которого входят центриоли.Центриолъ(под электронным микроскопом) имеет вид «полого» цилиндра диаметром около 150нм и длиной 300—500нм. Ее стенка образована 27микротрубочками, сгруппированными в 9триплетов. В функцию центриолей входит образование нитей митотического веретена, которые также образованы микротрубочками. Центриоли поляризуют процесс деления клетки, обеспечивая расхождение сестринских хроматид (хромосом) в анафазе митоза.
Эукариотическая клетка имеет клеточный скелет (цитоскелет) из внутриклеточных волокон (Кольцов) – начало ХХ века, в конце 1970 вновь открыт. Эта структура позволяет клетке иметь свою форму, иногда изменяя ее. Цитоплазма находится в движении. Цитоскелет участвует с процессе переноса органоидов, участвует в регенерации клеток.
Митохондрии – сложные образования с двойной мембраной(0,2-0,7мкм) и разной формой. Внутренняя мембрана имеет кристы. Наружная мембрана проницаема практически для всех химических веществ, внутренняя – только активный транспорт. Между мембранами – матрикс. Митохондрии располагаются там, где необходима энергия. Митохондрии имеют систему рибосом, молекулу ДНК. Возможно возникновение мутаций (более66 заболеваний). Как правило, они связаны с недостаточной энергией АТФ, часто связаны с сердечно-сосудистой недостаточностью, патологиями. Количество митохондрий разное (в клетке трипаносомы- 1 митохондрия). Количество зависит от возраста, функции, активности ткани (печень – более1000).
Лизосомы – тельца, окруженные элементарной мембраной. Содержат 60 ферментов( 40 лизосомальных, гидролитических). Внутри лизосомы – нейтральная среда. Активизируются низкими значениями рН, выходя в цитоплазму (самопереваривание). Мембраны лизосом защищают цитоплазму и клетку от разрушения. Образуются в комплексе Гольджи (внутриклеточный желудок, могут перерабатывать отработавшие свое структуры клетки). Есть 4 вида. 1-первичные, 2-4 – вторичные. С помощью эндоцитоза в клетку попадает вещество. Первичная лизосома (запасающая гранула) с набором ферментов, поглощает вещество и образуется пищеварительная вакуоль (при полном переваривании расщепление идет до низкомолекулярных соединений). Непереваренные остатки остаются в остаточных тельцах, которые могут накапливаться (лизосомные болезни накопления). Остаточные тельца, накапливающиеся в эмбриональном периоде, приводят к гаргалеизму, уродствам, мукополисахаридозам. Аутофагирующие лизосомы уничтожают собственные структуры клетки( ненужные структуры). Могут содержать митохондрии, части комплекса Гольджи. Часто образуются при голодании. Могут возникать при воздействии других клеток (эритроциты).
Источник: alexmed.info