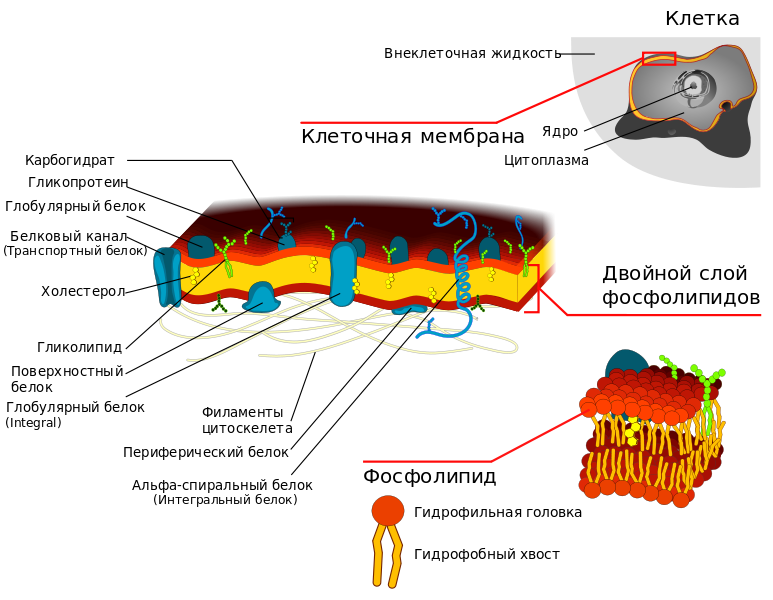
Строение плазматической мембраны клетки
Мы начнем гистологию с изучения эукариотической клетки, являющейся самой простой системой, наделенной жизнью. При исследовании клетки в световом микроскопе мы получаем информацию о ее размере, форме, и эта информация связана с наличием у клеток ограниченных мембраной границ. С развитием электронной микроскопии (ЭМ) наши представления о мембране, как о четко ограниченной линии раздела между клеткой и окружающей средой изменились, ибо оказалось, что на поверхности клетки имеется сложная структура, состоящая из следующих 3-х компонентов:
1. Надмембранный компонент (гликокаликс) (5 — 100 нм);
2. Плазматическая мембрана (8 — 10 нм);
3. Подмембранный компонент (20 – 40 нм).
При этом 1 и 3 компоненты вариабельны и зависят от типа клеток, наиболее статичным представляется строение плазматической мембраны, которую мы и рассмотрим.
Плазматическая мембрана. Изучение плазмолеммы в условиях ЭМ привело к заключению об однотипности ее структурной организации, при которой она имеет вид триламинарной линии, где внутренний и наружный слои электронноплотные, а расположенный между ними – более широкий слой представляется электроннопрозрачным.
кой тип структурной организации мембраны свидетельствует об ее химической гетерогенности. Не касаясь дискуссии по этому вопросу, оговорим, что плазмолемма состоит из трех типов веществ: липидов, белков и углеводов.
Липиды, входящие в состав мембран, обладают амфифильными свойствами за счет присутствия в их составе как гидрофильных, так и гидрофобных групп. Амфипатический характер липидов мембраны способствует образованию липидного бислоя. При этом в фосфолипидах мембраны выделяют два домена:
а) фосфатная– голова молекулы, химические свойства этого домена определяют его растворимость в воде и его называют гидрофильным;
б) ацильные цепи, представляющие собой этерифицированные жирные кислоты – это гидрофобный домен.
Типы мембранных липидов: основным классом липидов биологических мембран являются фосфолипиды, они формируют каркас биологической мембраны. См. рис.1

Рис. 1: Типы мембранных липидов
Биомембраны – это двойной слой амфифильных липидов(липидный бислой). В водной среде такие амфифильные молекулы самопроизвольно образуют бислой, в котором гидрофобные части молекул ориентированы друг к другу, а гидрофильные к воде. См. рис. 2
Рис. 2: Схема строения биомембраны
В состав мембран входят липиды следующих типов:
1. Фосфолипиды;
2. Сфинголипиды — “головки” + 2 гидрофобных “хвоста”;
3. Гликолипиды.
Холестерин (ХЛ)– находится в мембране в основном в срединной зоне бислоя, он амфифилен и гидрофобный (за исключением одной гидроксигруппы). Липидный состав влияет на свойства мембран: отношение белок/липиды близок 1:1, однако миелиновые оболочки обогащены липидами, а внутренние мембраны – белками.
Способы упаковки амфифильных липидов:
1. Бислои (липидная мембрана);
2. Липосомы — это пузырек с двумя слоями липидов, при этом как внутренняя, так и наружная поверхности являются полярны;
3. Мицеллы – третий вариант организации амфифильных липидов – пузырек, стенка которого образована одним слоем липидов, при этом их гидрофобные концы обращены к центру мицеллы и их внутренняя среда является не водной, агидрофобной.
Наиболее распространенной формой упаковки молекул липидов является образование ими плоского бислоя мембран. Липосомы и мицеллы – это скорые транспортные формы, обеспечивающие перенос веществ в клетку и из нее. В медицине липосомы используют для переноса водорастворимых, а мицеллы – для переноса жирорастворимых веществ.
Белки мембраны
1. Интегральные (включены в липидные слои);
2. Периферические. См. рис. 3
Интегральные (трансмембранные белки):
1. Монотопные– (например, гликофорин. Они пересекают мембрану 1 раз), и являются рецепторами, при этом их наружный – внеклеточный домен – относится к распознающей части молекулы;
2.Политопные– многократно пронизывают мембрану – это тоже рецепторные белки, но они активизируют путь передачи сигнала внутрь клетки;
3.Мембранные белки, связанные с липидами;
4. Мембранные белки, связанные с углеводами.
Рис. 3: Белки мембраны
Периферические белки:
Не погружены в липидный бислой и не соединены с ним ковалентно. Они удерживаются за счет ионных взаимодействий. Периферические белки ассоциированы с интегральными белками в мембране за счет взаимодействия — белок-белковые взаимодействия.
1. Спектрин, который расположен на внутренней поверхности клетки;
2.Фибронектин, локализован на наружной поверхности мембраны.
Белки – обычно составляют до 50% массы мембраны. При этом интегральные белкивыполняют следующие функции:
а) белки ионных каналов;
б) рецепторные белки.
А периферические мембранные белки (фибриллярные, глобулярные) выполняют следующие функции:
а) наружные (рецепторные и адгезионные белки);
б) внутренние – белки цитоскелета (спектрин, анкирин), белки системы вторых посредников.
Ионные каналы– это сформированные интегральными белками каналы, они формируют небольшую пору, через которую по электрохимическому градиенту проходят ионы. Наиболее известные каналы – это каналы для Nа, К, Са, Сl.
Существуют и водные каналы – это аквопорины (эритроциты, почка, глаз).
Надмембранный компонент – гликокаликс, толщина 50 нм. Это углеводные участки гликопротеинов и гликолипидов, обеспечивающие отрицательный заряд. Под ЭМ – это рыхлый слой умеренной плотности, покрывающий наружную поверхность плазмолеммы. В состав гликокаликса помимо углеводных компонентов входят периферические мембранные белки (полуинтегральные). Функциональные участки их находятся в надмембранной зоне — это иммуноглобулины. См. рис. 4
Функция гликокаликса:
1. Играют рольрецепторов;
2. Межклеточное узнавание;
3. Межклеточные взаимодействия (адгезивные взаимодействия);
4. Рецепторы гистосовместимости;
5. Зона адсорбции ферментов (пристеночное пищеварение);
6.Рецепторы гормонов.
Рис. 4: Гликокаликс и подмембранные белки
Подмембранный компонент — самая наружная зона цитоплазмы, обычно обладает относительной жесткостью и эта зона особенно богата филаментами (d = 5-10 нм). Предполагают, что интегральные белки, входящие в состав клеточной мембраны, прямо или косвенно связаны с актиновыми филаментами, лежащими в подмембранной зоне. При этом экспериментально доказано, что при агрегации интегральных белков, находящийся в этой зоне актин и миозин также агрегируют, что указывает на участие актиновых филамент в регуляции формы клетки.
| <== предыдущая лекция | | | следующая лекция ==> |
| РЫХЛАЯ ВОЛОКНИСТАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ | | |
Источник: helpiks.org
Функция и роль клеточной мембраны
Ее функция заключается в том, чтобы защитить целостность внутренней части клетки, впуская некоторые необходимые вещества в клетку, и не позволяя проникать другим.
Он также служит основой привязанности к цитоскелету у одних организмов и к клеточной стенке у других. Таким образом, плазматическая мембрана также обеспечивает форму клетки. Еще одна функция мембраны заключается в регулировании роста клеток через баланс эндоцитоза и экзоцитоза.
При эндоцитозе липиды и белки удаляются из клеточной мембраны по мере усвоения веществ. При экзоцитозе везикулы, содержащие липиды и белки, сливаются с клеточной мембраной, увеличивая размер клеток. Животные, растительные и грибковые клетки имеют плазматические мембраны. Внутренние органеллы, например, ядро, также заключены в защитные мембраны.
Структура клеточной мембраны
Плазматическая мембрана в основном состоит из смеси белков и липидов. В зависимости от расположения и роли мембраны в организме, липиды могут составлять от 20 до 80 процентов мембраны, а остальная часть приходится на белки. В то время как липиды помогают придать мембране гибкость, белки контролируют и поддерживают химический состав клетки, а также помогают в переносе молекул сквозь мембрану.
Липиды мембран
Фосфолипиды являются основным компонентом плазматических мембран. Они образуют липидный бислой, в котором гидрофильные (притянутые к воде) участки «головы» спонтанно организуются, чтобы противостоять водному цитозолю и внеклеточной жидкости, тогда как гидрофобные (отталкиваемые водой) участки «хвоста» обращены от цитозоля и внеклеточной жидкости. Липидный бислой является полупроницаемым, позволяя только некоторым молекулам диффундировать через мембрану.
Холестерин является еще одним липидным компонентом мембран животных клеток. Молекулы холестерина избирательно диспергированы между мембранными фосфолипидами. Это помогает сохранить жесткость клеточных мембран, предотвращая слишком плотное расположение фосфолипидов. Холестерин отсутствует в мембранах растительных клеток .
Гликолипиды расположены с наружной поверхности клеточных мембран и соединяются с ними углеводной цепью. Они помогают клетке распознавать другие клетки организма.
Белки мембран
Клеточная мембрана содержит два типа ассоциированных белков. Белки периферической мембраны являются внешними и связаны с ней путем взаимодействия с другими белками. Интегральные мембранные белки вводятся в мембрану, и большинство проходит сквозь нее. Части этих трансмембранных белков расположены по обе ее стороны.
Белки плазматической мембраны имеют ряд различных функций. Структурные белки обеспечивают поддержку и форму клеток. Белки рецептора мембраны помогают клеткам контактировать со своей внешней средой с помощью гормонов, нейротрансмиттеров и других сигнальных молекул. Транспортные белки, такие как глобулярные белки, переносят молекулы через клеточные мембраны посредством облегченной диффузии. Гликопротеины имеют прикрепленную к ним углеводную цепь. Они встроены в клеточную мембрану, помогая в обмене и переносе молекул.
Мембраны органелл
Некоторые клеточные органеллы также окружены защитными мембранами. Ядро, эндоплазматический ретикулум, вакуоль, лизосома и аппарат Гольджи являются примерами окруженных мембраной органелл. Митохондрии и хлоропласты покрыты двойной мембраной. Мембраны различных органелл различаются по молекулярному составу и хорошо подходят для выполнения своей роли. Они важны для нескольких жизненно важных функций клеток, включая синтез белка, производство липидов и клеточное дыхание.
Понравилась статья? Поделись с друзьями:
Источник: NatWorld.info
Строение клеточной мембраны
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.
В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков. Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.
Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Большинство погруженных в клеточную мембрану белков выполняют ферментативную функцию (являются ферментами). Часто (особенно в мембранах органоидов клетки) ферменты располагаются в определенной последовательности так, что продукты реакции, катализируемые одним ферментом, переходят ко второму, затем третьему и т. д. Образуется конвейер, который стабилизируют поверхностные белки, т. к. не дают ферментам плавать вдоль липидного бислоя.
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции. Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).
При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO2, O2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану. Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример — натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами). Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз — это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа. При этом могут открываться каналы, и в клетку могут начать поступать определенные вещества или выводиться из нее.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) — синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
Источник: biology.su