Складки внутренней мембраны митохондрий называются
Строение
Митохондрии – округлые или вытянутые органоиды, размеры которых колеблются от 0,2 до 2 мкм. Органеллы состоят из двух мембран. Внешняя мембрана гладкая, внутренняя образует складки – кристы, отвечающие за клеточное дыхание. Между мембранами находится пространство в 6-10 нм.
Складки внутренней мембраны заполнены жидкостью – матриксом, который содержит рибосомы, белки, ферменты, ДНК, РНК.

Рис. 1. Внутреннее строение митохондрии.
Существует гипотеза, что митохондрии имеют бактериальное происхождение. Прокариоты захватывали посредством фагоцитоза бактерии, которые могли генерировать энергию. Постепенно бактерии внедрились в структуру клетки и стали её органеллой.
Дыхание
Процесс окисления, т.е. клеточного дыхания, происходит в матриксе и на внутренних мембранах митохондрии. При метаболизме сложные вещества расщепляются до мономеров. Крахмал распадается до глюкозы, которая в бескислородной среде цитоплазмы расщепляется до пировиноградной кислоты (ПВК). При этом образуется две молекулы АТФ. В присутствии кислорода ПВК окисляется до углекислого газа и воды, т.е. осуществляется процесс дыхания в митохондриях.
Окисление происходит в два этапа:
- в матриксе – образуется углекислый газ, водород и 2 молекулы АТФ (цикл Кребса);
- на кристах – окисление водорода, образование воды и 36 молекул АТФ.
Дыхание на кристах (транспорт электронов) осуществляется с помощью дыхательной цепи, являющейся частью окислительного фосфорилирования (образования АТФ) и состоящей из двух компонентов:
- белковых комплексов (I, III и IV), встроенных в мембрану;
- белковых молекул-переносчиков (цитохром и убихинон).
Всего образуется 38 молекул АТФ, которые используются в процессах анаболизма. Вот почему митохондрии называют энергетическими станциями клеток.
Рис. 2. Схема дыхания в митохондриях.
Пластиды
Дополнительными органеллами растительной клетки, схожими по строению и функциям с митохондриями, являются пластиды. Они состоят из двух или четырёх мембран и бывают трёх видов:
- лейкопласты;
- хромопласты;
- хлоропласты.
Лейкопласты – бесцветные органеллы, которые зачастую находятся в корнях растений (не на свету). Они накапливают питательные вещества, например, в виде крахмала. На свету в лейкопластах образуется хлорофилл – зелёный пигмент.
Хромопласты содержат пигменты разных цветов (красный, жёлтый, фиолетовый). Они находятся в лепестках цветов и придают окраску венчику для привлечения насекомых.
Хлоропласты содержат пигменты (хлорофилл, каротиноид, ксантофилл), с помощью которых осуществляется процесс фотосинтеза. Внутри содержится студенистое вещество – строма, отвечающее за тёмную фазу фотосинтеза. В строме находятся ДНК, масла, рибосомы, а также мембранные структуры – тилакоиды, которые образуют граны, похожие на стопки монет. Тилакоиды отвечают за световую фазу фотосинтеза. Хлоропласты могут превращаться в лейкопласты или хромопласты.
Рис. 3. Внутреннее строение хлоропласта.
Источник: obrazovaka.ru
|
|
| Астрономия Биология География Дом и сад Другие языки Другое Информатика История Культура Литература Логика Математика Медицина Металлургия Механика Образование Охрана труда Педагогика Политика Право Психология Религия Риторика Социология Спорт Строительство Технология Туризм Физика Философия Финансы Химия Черчение Экология Экономика Электроника |
Внутренняя мембрана митохондрий образует гребневидные впячивания – кристы – разнообразной формы, на поверхности которых есть грибовидные тела – комплексы фермента АТФ-аза. Пространство между внутренней и внешней мембранами заполнено межмембранным матриксом. Кристы могут терять связь с внутренней мембраной и превращаться в замкнутые полости. В этом случае содержимое таких полостей все равно называется межмембранным матриксом. Наличие крист увеличивает поверхность внутренней (активной) мембраны. Внутреннее содержимое митохондрии называется внутренний матрикс, или просто матрикс. В матриксе содержатся: митохондриальные ДНК, РНК, рибосомы и включения. Таким образом, митохондрии обладают собственным белоксинтезирующим аппаратом. Дополнительные функции митохондрий: регуляция водного режима, хранение питательных веществ, хранение части генетической информации и биосинтез некоторых белков. Форма митохондрий зависит от таксономической принадлежности организмов, от тканевой принадлежности клеток и от физиологического состояния клеток. Крупные разветвленные митохондрии могут дробиться на множество мелких, а затем вновь сливаться. За счет этих преобразований число митохондрий в клетке может изменяться от 1 до десятков тысяч. В соматических клетках млекопитающих обычно содержится 500-1000 митохондрий. У аэробных прокариот и мезокариот митохондрий нет. Их функции выполняют мезосомы. Роль межмембранного матрикса играет пространство между плазмалеммой и клеточной стенкой. Митохондрии также отсутствуют у некоторых анаэробных паразитических Одноклеточных. У анаэробных паразитических червей (например, у аскариды) митохондрии выполняют запасающие функции. Пластиды Пластиды – специализированные полуавтономные двумембранные органоиды растений, выполняющие разнообразные функции. Пластиды описал А. ван Левенгук (1676), но их подробное изучение связано с развитием биохимии и электронной микроскопии. Пластиды различаются по форме, размерам, строению и функциям. В специализированных клетках обычно обнаруживается только один тип пластид. Исходной формой пластид являются пропластиды, или архепластиды. Увеличиваясь в размерах, пропластиды превращаются в лейкопласты. Лейкопласты в своем развитии превращаются или в хлоропласты, или в другие типы пластид: амилопласты содержат крахмал и превращаются в итоге в крахмальные зерна; липидопласты накапливают липиды и, подобно сферосомам, превращаются в масляные капли; протеинопласты накапливают белки и становятся их хранилищем. Хромопласты представляют собой последнюю стадию существования хлоропластов: в них происходит разрушение зеленых пигментов, но длительное время сохраняются желтые и красные пигменты. Происхождение пластид в филогенезе и онтогенезе остается неясным. Твердо установлено, что пластиды никогда не образуются заново; механизмы увеличения числа пластид в клетке, вероятно, разнообразны: деление, дробление, почкование. В изолированной культуре пластиды не могут существовать длительное время. Хлоропласты – пластиды, в которых протекают все реакции фотосинтеза: фотофосфорилирование и фиксация углекислого газа. Форма и количество хлоропластов в клетках относительно постоянны и зависят от таксономической принадлежности организмов, а также от уровня полиплоидии (у полиплоидов пластид больше). Обычно в клетке содержится несколько десятков хлоропластов. Впервые участие хлоропластов в процессах фотосинтеза показал Т. Энгельман (1883), хотя еще Ч. Дарвин считал, что хлорофилл – самое интересное из созданных природой органических веществ. Способность хлоропластов осуществлять все процессы фотосинтеза тесно связана с их строением. Внутреннее содержимое хлоропластов называется строма. Строма содержит пластидные ДНК, РНК, рибосомы и включения. Таким образом, хлоропласты обладают собственным белоксинтезирующим аппаратом. Впячивания внутренней мембраны образуют тилакоиды, заполненные матриксом. Одиночные тилакоиды называются ламеллы (или фреты), комплексы (стопки) тилакоидов – граны. Мембраны тилакоидов содержат комплексы пигментов (фотосистемы) и аналоги грибовидных тел (фермент АТФ-аза). Дополнительные функции пластид (в том числе, и хлоропластов) – те же, что и у митохондрий: регуляция водного режима, хранилище питательных веществ, хранение части генетической информации и биосинтез некоторых белков. У фотосинтезирующих пурпурных и зеленых бактерий пластиды отсутствуют. Их функции выполняют разнообразные мезосомы: прокариотические тилакоиды, ламеллы и хроматофоры. У цианобактерий в цитоплазме имеются ламеллы, по структуре сходные с ламеллами высших растений. У водорослей пластиды обычно называются хроматофоры. Форма хроматофоров: париетальные (постенные), чашевидные, кольцеобразные, цилиндрические, спиральные, звездчатые. Число тилакоидов в составе одной ламеллы – от 1 до 3. У зеленых водорослей имеются граны. Количество оболочек (мембран) различно: 2 (красные и зеленые водоросли), 3 (эвгленовые и пирофитовые) и 4 (золотистые, желто-зеленые, диатомовые и бурые). При наличии 4 мембран внешняя мембрана постепенно переходит в мембраны эндоплазматической сети и ядерной оболочки. В состав хроматофоров входит специфическая белковая структура – пиреноид. Вокруг пиреноида откладывается крахмал. Пиреноиды имеются также в пластидах некоторых моховидных. Задания 1. Заполните таблицу 15 «Сравнительная характеристика митохондрий и хлоропластов». При наличии признака поставьте в соответствующую ячейку знак +. Сделайте вывод о причинах сходства и причинах различий митохондрий и хлоропластов. 2. Анализ «слепых» препаратов.
|
Источник: mykonspekts.ru
Немецкие ученые выяснили, как происходит формирование сложной структуры митохондриальных мембран. Выяснилось, что помимо образования изгибов, необходимо действие еще по крайней мере двух белков. Один из них «раскрывает» второй, после чего второй присоединяется к месту сочленения внутренней и внешней мембраны, а также меняет форму внутренней мембраны, создавая тубулоподобные структуры, которые в результате становятся кристами. Работа опубликована в Nature Communications.
Митохондрия — двумембранная органелла эллипсоидной формы. Митохондрии являются «энергетической станцией» эукариот, так как в ее мембранах происходит окисление органических соединений и использование высвобождающейся при этом энергии для генерации электрического потенциала, тепла и синтеза «батареек» — АТФ.
В соответствии с широко распространенной теорией симбиогенеза, когда-то давно митохондрии были самостоятельными бактериальными организмами. Симбиоз с ними, начавшийся около двух миллиардов лет назад, обеспечил эукариотам налаженную систему энергосбережения, а им, вероятно, дал взамен защиту и питание. У митохондрий до сих пор осталась собственная ДНК, кодирующая белки, но многие гены они утратили, и существовать самостоятельно они не могут.
Наружная мембрана митохондрий гладкая, имеет толщину около семи нанометров и наполовину состоит из липидов. В ней также имеются порины и ряд ферментов. Она служит, в основном, в качестве ограничивающего от цитоплазмы барьера, а также для транспорта веществ и, например, контакта с эндоплазматическим ретикулумом.
жду внешней и внутренней мембраной имеется межмембранное пространство толщиной 10-20 нанометров. Внутренняя мембрана на три четверти состоит из белков, не имеет поринов и несет элементы дыхательной цепи, АТФ-синтазные комплексы и транспортные комплексы. Она образует многочисленные складки — кристы, и за счет этого ее площадь примерно в пять раз больше, чем площадь наружной мембраны. Внутри этой мембраны располагается митохондриальный матрикс.
Структуру двух мембран можно представить как многопальцевую перчатку, вшитую внутрь варежки. До сих пор было не вполне ясно, как именно происходит формирование такого лабиринта. Правильная структура важна не только для работы энергетических систем, но и, например, для деления митохондрий, и нарушения ее ведут к разрыву митохондрий, причем содержимое их может отравлять клетку.
Места, в которых кристы прилегают к внешней мембране, называются «сочленениями крист». Такое сочленение содержит комплекс MICOS, который помимо митохондриальных мембран включает по крайней мере семь белков. Именно он обеспечивают правильное формирование крист.
Два белка, Mic10 и Mic60, по-видимому, являются ключевыми элементами этого процесса, поскольку удаление их приводило к разрушению структуры внутренней мембраны. Было известно, что Mic10 принимает форму изогнутого каркаса, и движется вдоль мембраны, к которой он прикреплен. Именно он, по всей видимости, обеспечивает изгибание крист. Однако роль Mic60 до сих пор оставалась неясна.
Ученые выделили белки комплекса MICOS и рассматривали их взаимодействие с липидными мембранами (липосомами) отдельно от клетки. Оказалось, что Mic60 в свободной форме не прикреплен к мембране, а свернут таким образом, что соответствующее место крепления (сайт) его оказывается недоступным. Этот сайт консервативен у мышей, людей, бактерий, дрожжей, птиц и других организмов, что свидетельствует о его значительном возрасте и, соответственно, эволюционной значимости. При взаимодействии с комплексом MICOS, а именно, с белком Mic19, белок меняет конформацию, и сайт оказывается снаружи. Белок прикрепляется к внутренней мембране в месте ее сочленения с внешней мембраной.
Выяснилось, что помимо межмембранного контакта, он при этом выполняет еще важнейшую роль в формировании структуры крист. Он стабилизирует их изгибы, закрепляя мембраны в пространстве в виде длинных тубул. Такие белки можно представить как резинки, надетые на пальцы перчатки и держащие их на определенном расстоянии друг от друга. В отсутствии Mic19 этого не происходит. Таким образом, система Mic19–Mic60 оказывается необходимой для правильного формирования мембранной структуры митохондрий.
А о том, как митохондриями обмениваются нервные клетки, можно прочитать здесь.
Анна Казнадзей
Источник: nplus1.ru
Каждая митохондрия состоит из наружной и внутренней мембран, между которыми находится межмембранное пространство (Рис.7). Внутренняя мембрана образует складки — кристы, обращенные внутрь митохондрии. Пространство, ограниченное внутренней мембраной, заполнено митохондриальным матриксом, — мелкозернистым материалом различной электронной плотности.
Рис.7.
Наружная мембрана митохондрий содержит много молекул специализированных транспортных белков (например, порин), что обеспечивает её высокую проницаемость, а также белки-рецепторы, распознающие белки, которые переносятся через обе мембраны митохондрий в особых точках их контакта – зонах слипания.
Внутренняя мембранамитохондрий образует складки – кристы, благодаря чему значительно увеличивается внутренняя поверхность митохондрий. В состав внутренней мембраны входят транспортные белки; ферменты дыхательной цепи и сукцинатдегидрогеназа; комплекс АТФ-синтетазы. На кристах имеются элементарные частицы (оксисомы, или F1-частицы), состоящие из округлой головки (9 нм) и цилиндрической ножки. Именно на них происходит сопряжение процессов окисления и фосфорилирования (АДФ → АТФ).
Чаще всего кристы располагаются перпендикулярно длинной оси митохондрий и имеют пластинчатую (ламеллярную) форму. В клетках,синтезирующих стероидные гормоны, кристы имеют вид трубочек или пузырьков — тубулярно-везикулярные кристы. В этих клетках ферменты стероидного синтеза частично локализуются на внутренней мембране митохондрий
Число и площадь крист отражает функциональную активность клеток: наибольшая площадь крист характерна, например, для митохондрий клеток сердечной мышцы, где потребность в энергии постоянно очень велика.
Митохондриальный матрикс – мелкозернистое вещество, заполняющее полость митохондрии. Матрикс содержит несколько сотен ферментов: ферменты цикла Кребса, окисления жирных кислот, белкового синтеза. Здесь иногда встречаются митохондриальные гранулы, а также локализуются митохондриальные ДНК, иРНК, тРНК, рРНКи митохондриальные рибосомы.Митохондриальные гранулы – частицы высокой электронной плотности диаметром 20-50 нм, содержащие ионы Са и Мg.
Митохондриальная ДНК имеет кольцевую форму и включает 37 генов. Генетическая информация митохондриальной ДНК обеспечивает синтез около 5-6% белков митохондрий (ферменты электрон-транспортной системы). Синтез других митохондриальных белков контролируется ДНК ядра. Наследование митохондриальной ДНК происходит только по материнской линии.
Повреждения митохондриальной ДНК в результате мутаций могут привести к развитию ряда патологий — митохондриальных цитопатий (синдромы Барта, Патерсона, МERRF (красных разорванных волокон) и др.).
ЛИЗОСОМЫ – мембранные органеллы, которые обеспечивают внутриклеточное переваривание (расщепление) макромолекул внеклеточного и внутриклеточного происхождения, и обновление компонентов клетки.
Морфологически лизосомы представляют собой округлые пузырьки, ограниченные мембраной и содержащие большое количество различных гидролаз (более 60 ферментов). Наиболее характерными ферментами лизосом являются: кислая фосфатаза (маркёр лизосом), протеазы, нуклеазы, сульфатазы, липазы, гликозидазы. Все литические ферменты лизосом представляют собой кислые гидролазы, т.е. оптимум их активности проявляется при рН≈5.
Мембрана лизосом (около 6 нм толщиной) обладает протонным насосом, вызывающим закисление среды внутри органелл, обеспечивает диффузию низкомолекулярных продуктов переваривания макромолекул в гиалоплазму и препятствует утечке литических ферментов в гиалоплазму.
Повреждение мембраны приводит к разрушению клетки вследствие самопереваривания.
Лизосомы присутствуют во всех клетках. Особенно много лизосом в тех клетках, где активно протекают процессы фагоцитоза с последующим перевариванием захваченного материала (например, в нейтрофильных гранулоцитах, макрофагах, остеокластах).
Лизосомы подразделяются на первичные (неактивные)ивторичные (активные).
Первичные лизосомы (гидролазные пузырьки) – округлые пузырьки небольшого размера (обычно около 50 нм диаметром), с мелкозернистым, гомогенным, плотным матриксом. Надежная идентификация первичных лизосом возможна только при гистохимическом выявлении характерных ферментов (кислая фосфатаза). Первичные лизосомы – неактивные структуры, еще не вступившие в процессы расщепления субстратов.
Вторичные лизосомы – органеллы, активно участвующие в процессах внутриклеточного переваривания. Диаметр вторичных лизосом обычно составляет 0.5-2 мкм, их форма и структура могут существенно варьировать в зависимости от перевариваемого субстрата, но обычно содержимое вторичных лизосом гетерогенно.
Вторичная лизосома – результат слияния первичной лизосомы с фагосомой или аутофагосомой (Рис.8).
Фаголизосома формируется путем слияния первичной лизосомы с фагосомой — мембранным пузырьком, содержащим материал, захваченный клеткой извне. Процесс разрушения этого материала называется гетерофагией. Гетерофагия играет важную роль в функции всех клеток. Особое значение гетерофагия имеет для клеток, осуществляющих защитную функцию, таких как макрофаги и нейтрофильные лейкоциты, которые захватывают и переваривают болезнетворные микроорганизмы.
Аутофаголизосома образуется при слиянии первичной лизосомы с аутофагосомой — мембранным пузырьком, содержащим собственные компоненты клетки, которые подлежат разрушению. Процесс переваривания внутриклеточного материала называется аутофагией. Аутофагия обеспечивает постоянное обновление клеточных структур благодаря перевариванию митохондрий, полисом, фрагментов мембран.
Рис.8.
Остаточные тельца – лизосомы, содержащие непереваренный материал, которые могут находиться в цитоплазме длительное время. В некоторых долгоживущих клетках (нейроны, кардиомиоциты, гепатоциты) в остаточных тельцах накапливается коричневый эндогенный пигмент липофусцин – «пигмент старения».
Дефицит лизосомальных ферментов может приводить к развитию ряда заболеваний (болезни накопления), вызванных накоплением в клетках непереваренных веществ, которые нарушают функцию клеток. Примерами могут служить: болезнь Хюрлера, при которой из-за отсутствия α-L-идуронидазы фибробласты и остеобласты накапливают дерматан сульфат, а у больных отмечаются множественные дефекты хондро- и остеогенеза и умственное отставание; болезнь Тэя-Сакса (из-за недостаточности гексозаминидазы А происходит накопление гликолипидов в нервных клетках и поражается нервная система); болезнь Гоше (вследствие наследственного дефекта глюкоцереброзидазы гликолипиды накапливаются в макрофагах и поражаются печень и селезенка) и другие.
Пероксисомы– сферические мембранные органеллы диаметром 0.05 – 1.5 мкм, с умеренно плотным гомогенным или мелкозернистым матриксом. Мелкие пероксисомы встречаются во всех клетках, а крупные пероксисомы – в гепатоцитах, макрофагах, в клетках канальцев почки. Матрикс пероксисом содержит до 50 различных ферментов, важнейшие из которых: каталаза (маркёр пероксисом), пероксидаза, оксидазы аминокислот, уратоксидаза.
У некоторых видов животных в пероксисомах выявляется более плотная кристаллическая сердцевина – нуклеоид, состоящая из уратоксидазы. В пероксисомах клеток человека нуклеотида нет, поскольку отсутствует способность метаболизировать ураты.
Функции пероксисом:
• окисление аминокислот и других субстратов;
• защита клетки от действия перекиси водорода, сильного окислителя, образующегося в результате окисления органических соединений, и оказывающего повреждающий эффект на клетку. При этом каталаза пероксисом разлагает перекись водорода на воду и кислород.
• участие в расщеплении жирных кислот;
• участие в обезвреживании ряда веществ (спирт и др.).
Нарушения активности пероксисом вызывает ряд наследственных заболеваний – пероксисомных болезней с тяжелыми нарушениями нервной системы (синдром Целльвегера и др.)
Рис.9.
Цитоскелет – сложная трехмерная сеть немембранных органелл (рис.9):
· микротрубочек;
· микрофиламентов;
· промежуточных филаментов.
Основная функция цитоскелета – опорно-двигательная:
• поддержание и изменение формы клеток;
• перемещение компонентов внутри клетки;
• транспорт веществ внутрь клетки и из клетки;
• обеспечение подвижности клетки
Микротрубочки – наиболее крупные компоненты цитоскелета. Микротрубочки – полые цилиндрические образования различной длины, с диаметром 24-25 нм, с толщиной стенки 5 нм.
Стенка микротрубочки состоит из спирально
расположенных нитей – профиламентов, образованных димерами из глобулярных белковых молекул – α- и β-тубулина.
Стенка микротрубочки образована 13 субъединицами-профиламентами.
Микротрубочки могут располагаться в цитоплазме в виде отдельных элементов, в виде пучков, где они связаны тонкими поперечными мостиками, или могут частично сливаться друг с другом, образуя дуплеты (в аксонеме ресничек и жгутиков) и триплеты (в базальном тельце и центриолях.
Микротрубочки представляют собой лабильную систему, в которой сохраняется равновесие между их постоянной сборкой и диссоциацией.
Центрами организации микротрубочек (ЦОМТ) являются сателлиты – глобулярные белковые структуры, содержащиеся в базальных тельцах ресничек и клеточном центре, а также центромеры хромосом.
Функции микротрубочек:
· поддержание стабильной формы клеток, и порядка распределения её компонентов;
· обеспечение внутриклеточного транспорта, в том числе органелл, пузырьков, секреторных гранул (благодаря некоторым белкам, ассоциированным с микротрубочками);
· образование основы центриолей и ахроматинового веретена деления и обеспечение движения хромосом в процессе митоза;
· образование основы ресничек и жгутиков, а также обеспечение их движения.
Угнетение самосборки микротрубочек при действии на клетку блокаторов (колхицин и др.) вызывает гибель быстроделящихся клеток вследствие отсутствия митотического веретена деления, нарушения транспортных процессов в клетке (аксонный транспорт в нейронах, секреция), изменения форм клетки, дезорганизацию клеточных органелл (в частности, цистерн ЭПС).
Клеточный центр образован двумя полыми цилиндрическими структурами — центриолями, которые расположены под прямым углом друг к другу.
Каждая центриоль представляет собой короткий цилиндр длиной ~ 0,5 мкм и диаметром ~ 0,2 мкм, состоящий из 9 триплетов частично слившихся трубочек (А, В и С), связанных поперечными белковыми мостиками (рис.10).
Формула строения центриоли описывается как (9 × 3) + 0, так как в центральной части микротрубочки отсутствуют. Каждый триплет центриоли связан с глобулярными белковыми тельцами – сателлитами, от которых отходят микротрубочки, образующие центросферу.
Рис.10.
В неделящейся клетке выявляется одна пара центриолей – диплосома, которая располагается обычно вблизи ядра. Перед делением клетки в S-периоде интерфазы происходит дупликация центриолей: под прямым углом к каждой зрелой (материнской) центриоли пары образуется новая (дочерняя) центриоль.
В ранней профазе митоза пары центриолей расходятся к полюсам клетки и служат центрами образования микротрубочек ахроматинового веретена деления.
Реснички и жгутикиявляются выростами цитоплазмы, обладающие подвижностью. Основу ресничек и жгутиков составляет каркас из микротрубочек, называемый аксонемой (рис.11).
Длина ресничек равна 2-10 мкм, а их количество на поверхности одной клетки может составлять до нескольких сотен.
В организме человека жгутик есть только в одном типе клеток – сперматозоидах. При этом один сперматозоид имеет один жгутик длиной 50-70 мкм.
Рис.11.
Аксонемаобразована 9периферическими парами микротрубочек (микротрубочки А и В) и одной центрально расположенной парой; такое строение описывается формулой (9 × 2) + 2. Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дуплетам расходятся радиальные спицы. Периферические дублеты связаны друг с другом мостиками белка нексина, а от микротрубочки А к микротрубочке В соседнего дублета отходят “ручки” из белка динеина, который обладает АТФ-азной активностью, что необходимо для скольжения соседних дублетов в аксонеме, вызывающих движение (биение) ресничек и жгутиков
Мутации, вызывающих изменения белков ресничек и жгутиков, ведут к различным нарушениям функций клеток. Так, при отсутствии динеиновых ручек (синдром неподвижных ресничек, или синдром Картагенера), больные страдают хроническими заболеваниями дыхательной системы и бесплодием (вследствие неподвижности спермиев и нарушений продвижения яйцеклеток по яйцеводу).
В основании каждой реснички или жгутика лежит базальное тельце, сходное по строению с центриолью. На уровне апикального конца базального тельца микротрубочка С триплета заканчивается, тогда как микротрубочки А и В продолжаются в соответствующие микротрубочки аксонемы реснички. При развитии ресничек или жгутика базальное тельце играет роль матрицы, на которой происходит сборка компонентов аксонемы
Микрофиламенты – тонкие белковые нити диаметром 5-7 нм, расположенные в цитоплазме поодиночке, в виде сетей или упорядоченными пучками (в скелетной и сердечной мышцах). Основной белок микрофиламентов – актин – встречается в клетках как в мономерной форме (глобулярный G-актин), так и в виде полимерного фибриллярного F-актина.
Функции микрофиламентов:
• в мышечных волокнах и клетках актиновые микрофиламенты образуют упорядоченные пучки и при взаимодействии с миозиновыми филаментами обеспечивают их сокращение.
• в немышечных клетках микрофиламенты образуют кортикальную (терминальную) сеть, в которой микрофиламенты сшиты с помощью особых белков (филамин и др.). Кортикальная сеть, с одной стороны, обеспечивает поддержание формы клетки, а с другой — способствует изменениям формы плазмолеммы, обеспечивая, таким образом, функции эндо- и экзоцитоза, миграции клеток, образования псевдоподий.
• микрофиламенты тесно связаны с органеллами, транспортными пузырьками, секреторными гранулами и играют важную роль в их перемещении внутри цитоплазмы.
• микрофиламенты формируют сократимую перетяжку (срединное тельце) при цитотомии, завершающей клеточное деление.
• микрофиламенты участвуют в организации структуры межклеточных соединений (zonula adherens – поясок сцепления).
• микрофиламенты являются основой специальных выростов цитоплазмы – микроворсинок и стереоцилий.
Микроворсинки– пальцевидные выросты цитоплазмы клетки диаметром 0.1 мкм и длиной 1 мкм, основу которых образуют актиновые микрофиламенты (рис.12).
Микроворсинки обеспечивают многократное увеличение площади поверхности клетки. На апикальной поверхности некоторых клеток, активно участвующих в процессах расщепления и всасывания веществ, имеется до несколько тысяч микроворсинок, образующих в совокупности щёточную каемку(эпителий тонкой кишки и почечных канальцев).
Рис.12.
Основа каждой микроворсинки – пучок, содержащий около 40 микрофиламентов, расположенных вдоль её длинной оси. Микрофиламенты имеют поперечные сшивки из белков (фимбрин, виллин), и прикреплены к плазмолемме особыми белковыми мостиками (минимиозин). У основания микроворсинки микрофиламенты пучка вплетаются в терминальную сеть
Стереоцилии – длинные, иногда ветвящиеся микроворсинки, имеющие каркас из микрофиламентов. Они встречаются редко (например, в главных клетках эпителия протока придатка семенника).
Промежуточные филаменты – прочные и устойчивые белковые нити толщиной около 10 нм (что является промежуточнымзначением между толщиной микротрубочек и микрофиламентов). Промежуточные филаменты располагаются в виде трехмерных сетей в различных участках цитоплазмы, окружают ядро, участвуют в образовании межклеточных контактов (десмосом) и поддерживают форму отростков.
Главная функция промежуточных филаментов – поддерживающая и опорная.
Промежуточные филаменты в клетках различных типов различаются по своей химической природе и молекулярному весу. Выделяют 6 основных классов промежуточных филаментов
Цитокератины –промежуточные филаменты, характерные для клеток эпителия. Этот класс включает около 20 близких полипептидов (тонофиламентов). Кератиновые филаменты входят в состав десмосом и полудесмосом, участвуют в образовании рогового вещества в эпителии кожи и являются главным компонентом волос и ногтей.
Десмины – промежуточные филаменты мышечных тканей (за исключением миоцитов сосудов). Десмины играют важную роль в организации миофибрилл в мышечной ткани и обеспечении сократительной функции
Виментины – филаменты, характерные для различных клеток мезенхимного происхождения (фибробласты, макрофаги, остеобласты, эндотелий и гладкие миоциты сосудов).
Нейрофиламенты – промежуточные филаменты нейронов, которые играют важную роль в поддержании формы отростков нервных клеток.
Глиальные клетки содержат глиальный фибриллярный кислый белок и встречаются только в клетках нейроглии (астроциты, олигодендроциты).
Идентификация классов промежуточных филаментов (методами иммуноцитохимии с антителами к данному типу промежуточных филаментов) имеет большое значение в диагностике опухолей, и, следовательно, в прогнозе и выборе противоопухолевого лечения. Так, выявление различных форм кератинов свидетельствует о недифференцированных опухолях эпителиального происхождения, карциномах, аденокарциномах. Десмин является маркёром опухолей мышечного происхождения, а глиальный фибриллярный кислый белок – маркёр опухолей глиального происхождения.
ВКЛЮЧЕНИЯ
В отличие от органелл, включения цитоплазмы – непостоянные компоненты цитоплазмы, возникающие и исчезающие в зависимости от метаболического состояния клеток.
Включения подразделяются на трофические, секреторные, экскреторные и пигментные.
Трофические включения разделяются в зависимости от природы накапливаемого вещества на липидные, углеводные и белковые. Липидные включения – это капли нейтрального жира различного диаметра, которые накапливаются в цитоплазме и служат резервом энергетических субстратов, используемых клеткой. Из углеводных включений наиболее распространены гранулы гликогена (полимер глюкозы), эти включения также используются в качестве источника энергии. Примером белковых включений могут служить запасы белка вителлина в яйцеклетках животных. Они являются источником питания на ранних стадиях развития зародыша.
Секреторные включения имеют вид пузырьков, окруженные мембраной и содержащие биологически активные вещества, которые синтезируются в самой клетке, а затем выделяются (секретируются) во внешнюю среду. К таким включениям относятся секреторные гранулы, содержащие пищеварительные проферменты (зимогеновые гранулы), гормоны, медиаторы и др.
Экскреторные включения по своему строению сходны с секреторными, но в отличие от них, содержат вредные продукты метаболизма, подлежащие удалению из цитоплазмы клеток.
Пигментные включения представляют собой скопления эндогенных (синтезированных клеткой), или экзогенных (захваченных клеткой извне) окрашенных веществ — пигментов. Наиболее распространенными эндогенными пигментами являются гемоглобин, гемосидерин, билирубин, меланин, липофусцин; к экзогенным пигментам относят каротин, различные красители, пылевые частицы и др. Меланин – тёмно-коричневый пигмент, встречающийся в норме в коже, волосах, пигментной оболочке сетчатки в виде меланосом — гранул, окруженных мембраной. Липофусцин – гранулы жёлто-коричневого пигмента из продуктов лизосомного переваривания – накапливается в долгоживущих клетках (нейроны, кардиомиоциты), и поэтому его рассматривают как «пигмент старения».
ГИАЛОПЛАЗМА
Гиалоплазму называют также цитозолем, или клеточным матриксом. Гиалоплазма – сложная коллоидная система, которая может менять своё агрегатное состояние: переходить из более жидкого (золь) в более плотное (гель). Гиалоплазма состоит из гомогенного мелкозернистого вещества с низкой электронной плотностью, в которое погружены органеллы и включения. В составе гиалоплазмы – вода, белки (ферменты), нуклеиновые кислоты, полисахариды, липиды, а также неорганические вещества.
Функции гиалоплазмы:
· создание жидкой микросреды;
· метаболическая: метаболизм белков, жиров, углеводов.
III. ЯДРО. КЛЕТОЧНЫЙ ЦИКЛ. ДЕЛЕНИЕ КЛЕТКИ. КЛЕТОЧНЫЕ ПОПУЛЯЦИИ. ГИБЕЛЬ КЛЕТОК.
Ядро– важнейший компонент клетки, содержащий её генетический аппарат.
Функции ядра:
· хранение генетической информации (в молекулах ДНК, находящихся в хромосомах);
· реализация генетической информации, контролирующей различные процессы в клетке: транскрипция информационных, рибосомальных, транспортных РНК → синтетическая активность; апоптоз и т.д.);
· воспроизведение и передача генетической информации при делении клетки
В ядре неделящейся (интерфазной) клетки выявляются следующие компоненты ядра:
· ядерная оболочка (кариолемма);
· хроматин;
· ядрышко;
· кариоплазма.
Ядерная оболочка (кариолемма, нуклеолемма) на светооптическом уровне практически не определяется. Под электронном микроскопом обнаруживается, что она состоит из двух мембран – наружной и внутренней мембран, разделенных полостью шириной 15-40 нм – перинуклеарной цистерной.
Наружная мембранасоставляет единое целое с мембранами грЭПС: на её поверхности имеются рибосомы, а перинуклеарная цистерна сообщается с цистерной грЭПС
Внутренняя мембрана –гладкая, её интегральные белки связаны со слоем, состоящим из сети промежуточных филаментов (ламинов), — ламиной, или ядерной пластинкой. Ламина играет большую роль в поддержании формы ядра, укладке хроматина и структурной организации поровых комплексов.
В определенных точках наружная и внутренняя мембрана смыкаются, образуя ядерные поры. Ядерная пора образована двумя параллельными кольцами диаметром 80 нм, содержащих по 8 белковых гранул, от которых к центру поры тянутся фибриллы, формирующие диафрагму толщиной около 5 нм. В середине диафрагмы лежит центральная гранула. Белковые гранулы ядерной поры структурно связаны с белками ламины. Совокупность компонентов, входящих в состав ядерной поры, называется комплексом ядерной поры.
Ядерная оболочка клетки содержит 2000-4000 поровых комплексов. Число поровых комплексов возрастает с увеличением функциональной активности: в клетках с высокой синтетической активностью ядерные поры занимают до 35% поверхности кариолеммы.
Комплекс ядерной поры обеспечивает избирательный транспорт веществ между цитоплазмой и ядром. По каналу, образованному поровым комплексом, движутся мелкие водорастворимые молекулы и ионы; активно переносятся в ядро белки, синтезируемые в цитоплазме (белки с маркировкой в виде с особой последовательности аминокислот – NLS, распознаваемой рецепторами NLS в комплексе поры); из ядра в цитоплазму переносятся субъединицы рибосом.
Хроматин в интерфазной (неделящейся) клетке соответствует хромосомам и состоит из комплекса ДНК и белка. Выраженность спирализации каждой из хромосом неодинакова по длине. Соответственно, различают два вида хроматина: эухроматин и гетерохроматин.
Эухроматин соответствует участкам хромосом, которые деспирализованы и открыты для транскрипции. Эти участки не окрашиваются и не видны в световой микроскоп.
Гетерохроматин соответствует конденсированным сегментам хромосом, что делает их недоступными для транскрипции. Гетерохроматин интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид мелких гранул и глыбок.
По соотношению эу- и гетерохроматина в ядре можно оценить активность процессов транскрипции, и, следовательно, синтетической функции клетки. При её повышении это соотношение изменяется в пользу эухроматина, при снижении – нарастает содержание гетерохроматина. Соотношение эухроматин-гетерохроматин может, например, служить основой для дифференциальной диагностики доброкачественных и злокачественных опухолевых клеток.
При полном подавлении функции ядра в поврежденных и гибнущих клетках, оно уменьшается в размерах и содержит только гетерохроматин. Такое явление называется кариопикнозом.
Половой хроматин (тельце Барра) – скопление гетерохроматина, соответствующее одной из пары Х-хромосом, которая в интерфазе плотно скручена и неактивна.
Выявление полового хроматина используется как диагностический тест для определения генетического женского пола, что существенно при изучении генетических аномалий и, особенно, в спортивной медицине. Обычно анализируют эпителиальные клетки слизистой оболочки полости рта, где, как и в большинстве других клеток, половой хроматин выявляется как крупная глыбка гетерохроматина, лежащая рядом с ядерной оболочки. В нейтрофильных лейкоцитах крови половой хроматин имеет вид маленькой добавочной дольки ядра («барабанной палочки»).
Упаковка хроматина в ядре
В деконденсированном состоянии длина одной молекулы (двойной спирали) ДНК, образующей одну хромосому, составляет около 5 см, а общая длина молекул ДНК в ядре – более 2 м. Такие длинные нити ДНК компактно и упорядоченно упакованы в ядре диаметром всего 5-10 мкм.
Компактная упаковка молекул ДНК осуществляется благодаря связи ДНК со специальными основными белками – гистонами.
Начальный уровень упаковки хроматина – нуклеосома с
диаметром 11 нм.
· Нуклеосома состоит из блока, образованного комплексом из 8 молекул гистонов, на который намотана двойная нить ДНК (цепочка из 166 пар нуклеотидов).
· Нуклеосомы разделены короткими участками свободной ДНК (48 пар
оснований). Нуклеосомная нить имеет вид нитки с бусинами, где каждая бусина – нуклеосома.
· Второй уровень упаковки также обусловлен гистонами и приводит к скручиванию нуклеосомной нити (виток из 6 нуклеосом) с формированием хроматиновой фибриллы диаметром 30 нм.
· Хроматиновые фибриллы образуют петли диаметром 300 нм. При делении клетки в результате еще более компактной укладки и сверхспирализации ДНК появляются хромосомы (диаметр 700 нм), видимые под световым микроскопом.
Компактная упаковка ДНК в ядре обеспечивает упорядоченное расположение очень длинных молекул ДНК в небольшом объеме ядра, а также функциональный контроль активности генов.
Ядрышко выявляется в интерфазном ядре на светооптическом уровне как мелкая (~ 1 мкм в диаметре), плотная сферическая структура, интенсивно окрашивающееся основными красителями. Ядрышко образовано специализированными участками хромосом – ядрышковыми организаторами, на которых происходит синтез рибосомальной РНК, а также её сборка в предшественники рибосомальных субъединиц.
Компоненты ядрышка:
· Аморфный компонент, слабо окрашиваемый, представляет собой участки расположения ядрышковых организаторов: крупные петли ДНК, активно участвующих в транскрипции рибосомальной РНК;
· Фибриллярный компонент состоит из множества нитей диаметром 5-8 нм, преимущественно во внутренней части ядрышка, и представляет собой длинные молекулы рРНК (первичные транскрипты);
· Гранулярный компонентобразован скоплением плотных мелких гранулярных частиц, представляющие собой зреющие субъединицы рибосом. Рибосомальные субъединицы образуется из рРНК, синтезированной в ядрышке, и белков, синтезированных в цитоплазме.
· Фибриллярный и гранулярный компоненты ядрышка образуют ядрышковую нить – нуклеолонему, которая образует петлистую сеть, выделяющуюся большой плотностью на фоне менее плотного ядерного матрикса
Размеры и объем ядрышек увеличиваются при повышении функциональной активности клетки. Особенно крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также клеток быстрорастущих злокачественных опухолей.
Ядрышко исчезает в профазе митоза, в результате инактивации рибосомных генов при конденсации соответствующих хромосом, и вновь формируется в поздней телофазе.
Ядерный матрикс – компонент ядра, в котором располагаются хроматин и ядрышко. Ядерный матрикс образован кариоплазмой и кариоскелетом. Кариоплазма – жидкий компонент ядра, содержащий РНК, ионы, ферменты, метаболиты, растворенные в воде. Кариоскелет состоит из ламины и других фибриллярных белков.
Клеточный цикл – совокупность процессов, происходящих в клетке между двумя последовательными делениями или между её образованием и гибелью. Клеточный цикл включает в себя собственно митотическое деление и интерфазу – промежуток между делениями
Интерфаза занимает около 90% всего времени клеточного цикла и подразделяется на три периода:
· пресинтетический или постмитотический – G1 (от англ. gap – промежуток);
· синтетический – S;
· постсинтетическийилипремитотический — G2.
Пресинтетический период – G1 – характеризуется активным ростом клетки, синтезом белка и РНК, благодаря чему клетка восстанавливает необходимый набор органелл и достигает нормальных размеров. G1 период длится от нескольких часов до нескольких дней. В течение этого периода синтезируются особые «запускающие» белки – активаторы S периода. Они обеспечивают достижение клеткой точки R (точки ограничения), после которого она вступает в S-период.
Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G0).Клетки некоторых тканей под влиянием определенных факторов способны возвращаться из периода G0 в клеточный цикл, клетки других тканей (кардиомиоциты, нейроны) утрачивают эту способность по мере дифференцировки. Абсолютное большинство дифференцированных клеток организма, выполняющих свои специфические функции, не делятся.
Синтетический период –S- характеризуется репликацией (удвоением содержания) ДНК, синтезом гистонов и других белков. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. S-период длится у большинства клеток 8-12 часов.
Постсинтетический период – G2 — длится 2-4 часа и продолжается вплоть до митоза. В течение этого периода запасается энергия, и синтезируются белки, в частности тубулины, необходимые для процесса деления.
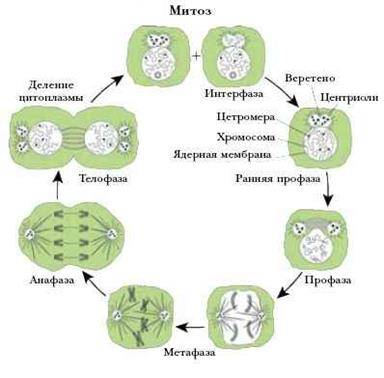
Митоз (кариокинез) является универсальным механизмом деления соматических клеток. Во время митоза родительская клетка делится, и каждая из дочерних клеток получает набор хромосом идентичный родительскому, и, таким образом, происходит равномерное распределение генетического материала. Продолжительность митоза – 1-3 часа.
Митоз условно разделяют на 4 основные фазы:
· профазу;
· метафазу;
· анафазу;
· телофазу.
Профазаначинается сконденсации хромосом, которые становятся видимыми в световой микроскоп как нитевидные структуры. Каждая хромосома состоит из двух параллельно лежащих сестринских хроматид, связанных в области центромеры. Ядерная оболочка распадается на мембранные пузырьки и исчезает к концу профазы, так же как и ядрышко. Кариоплазма смешивается с цитоплазмой. Пары центриолей расходятся к противоположным полюсам клетки и дают начало микротрубочкам митотического веретена.
В метафазе хромосомы выстраиваются в области экватора митотического веретена (в равной удаленности от центриолей противоположных полюсов), и образуют картину экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со сторону полюсов). Сестринские хроматиды к концу этой фазы разделяются щелью, однако удерживаются в области центромеры.
Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды (в области центромеры) и движения дочерних хромосом к противоположным полюсам клеток, происходящего вдоль микротрубочек. Анафаза завершается скоплением на полюсах клетки двух идентичных наборов хромосом, которые образуют картину звезд (стадия дочерних звезд). В конце анафазы начинает образовываться клеточная перетяжка, благодаря сокращению актиновых микрофиламентов, которые концентрируются по окружности клетки.
Телофазахарактеризуется реконструкцией ядер дочерних клеток и завершением их разделения. Ядерная оболочка восстанавливается, хромосомы постепенно деспирализуются, замещаясь картиной хроматина интерфазного ядра, а в конце телофазы вновь появляется ядрышко. Углубление клеточной перетяжки завершается полной цитотомией с формированием двух дочерних клеток. При этом происходит приблизительно равное распределение органелл между дочерними клетками.
Эндомитоз – процесс увеличения числа хромосом внутри ядерной оболочки без последующего деления клетки, что приводит к повышенному содержанию ДНК в ядре – полиплоидии.
Полиплоидные ядра имеют больший объем. Полиплоидные клетки могут также возникнуть вследствие митотического деления без последующей цитотомией. При таком делении образуются двуядерные клетки с увеличенным вдвое набором хромосом. Основной смысл развития полиплоидии заключается в усилении функциональной активности клеток.
Наличие полиплоидных – тетра- (4n, если 1n – гаплоидный набор хромосом) и октаплоидных (8n) клеток – нормальное явление для гепатоцитов (клеток печени), переходного эпителия мочевого пузыря, секреторных клеток поджелудочной и слюнных желез. Уровень полиплоидизации мегакариоцитов красного костного мозга достигает – 16-32n.
По уровню обновления ткани организма подразделяются на три группы – три типа клеточных популяций:
· Обновляющиеся клеточные популяции<
Источник: poznayka.org