Сколько мембран у митохондрии
Открыл в 1848 году, Альтман. Есть почти у всех эукариотов. Это подвижные органоиды, могут менять свою морфологию, объединяться, разъединяться с помощью микротрубочек. Различные по форме. Период жизни митохондрий короток. У человека они живут 9 суток. Более 10 суток митохондрии не живут, т.е., высокая степень обновления.
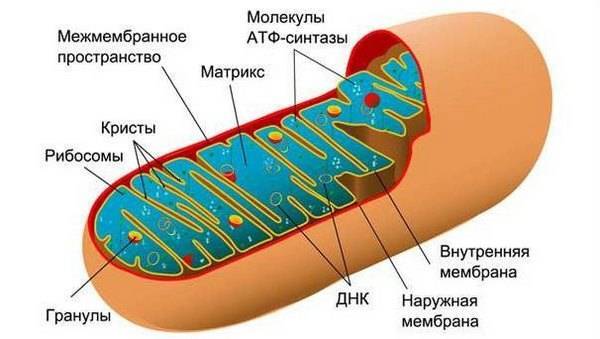
Основная функция митохондрий – синтез АТФ, дающий энергию для всех жизненных процессов. Митохондрии встречаются практически у всех эукариотических клеток, кроме двух видов паразитических амеб. Митохондрии являются подвижными пластичными органоидами, они могут объединяться в гигантские структуры и наоборот – разрушаться. Совокупность всех митохондрий в клетке – хондриом. Размеры варьируют от 1 до 10 мкм. Форма чрезвычайно разнообразна. Но несмотря на разнообразие форм и размеров, для всех митохондрий характерен единый тип строения. Митохондрия состоит из двух мембран, наружной и внутренней, меж которыми находится межмембранное пространство, которое по сути является протонным резервуаром. Внутри – митохондриальный матрикс. Наружная мембрана характеризуется большим количеством транспортных белков. Эти белки имеют специфическое название порины. Они образуют широкие гидрофильные каналы в бислое липидов, которые пропускают молекулы с молекулярной массой до 10000 Дальтон. На наружной мембране имеются ферменты, которые превращают липиды в реакционноспособные соединения. Белков менее 20%. Из физических свойств можно отметить, что наружная мембрана может только необратимо растягиваться. Внутренняя мембрана легко сморщивается и растягивается. Она образует многочисленные складки, которые называются кристы. Площадь внутренней мембраны почти в пять-семь раз больше, чем наружной. Кристы бывают двух типов – пластинчатые в клетках многоклеточных животных и растений; трубчатые в тканях синтезирующих ферменты. Кристы не статистические образования. Расположены параллельно или перпендикулярно длинной оси митохондрий. Степень развития крист, их количество определяется функциональной значимостью клетки. На внутренней мембране очень низок уровень холестерина. Много особого фосфолипида кардиолипида. Благодаря этому мембрана имеет малую проницаемость. При суммарном подсчете и вычислении соотношения белков и липидов получим, что белков 75%, а липидов 25%.
Белки: 1) Цепь переноса электронов; 2) Грибовидные тельца с АТФ-синтетазой; 3) Специфические транспортные белки, регулирующие перенос метаболитов в матрикс митохондрий и из матрикса.
Межмембранное пространство или протонный резервуар. По химическому составу почти идентичен цитозолю. Содержится только несколько ферментов для обеспечения реакции фосфорилирования. Матрикс – высококонцентрированная смесь, более чем из ста компонентов, большинство из которых – это ферменты цикла Кребса (карбоновых кислот). Здесь же располагаются кольцевые молекулы, митохондриальная ДНК, тРНК, иРНК и митохондриальные рибосомы с константой седиментации от 55 до 75S Белки цепи переноски электронов. Это крупные ферментные комплексы, по которым электроны опускаются на все более низкие энергетические уровни. Конечным акцептором является молекулярный кислород. В цепи выделяют электрон-собирающие и электрон-распределяющие участки и последовательные ряды ферментов, с которыми взаимодействуют электроны, можно разделить на три главных ферментативных комплекса: 1) НАДН+-дегидрогеназный комплекс. Состоит из двенадцати полипептидных цепочек. Главные из этих компонентов является НАДН+ и убихинон (ко Q). НАДН+-дегидрогеназа первая принимает 2 протона и 2 электрона.Убихинон – небольшой переходный переносчик, который тоже принимает и протоны и электроны. Переносит протоны через мембрану на наружную поверхность сопрягающей мембраны. 2) Комплексу B=С Этот комплекс состоит из 8 полипептидных цепей. Содержит два цитохрома (В и С), которые передают только электроны, а протоны сбрасываются в межмембранное пространство. При этом, цитохром С, принимающий электроны, передает их на 3 комплекс. 3) Третий компонент – цитохромоксидазный комплекс, который состоит из семи различных полипептидов. Он может передавать и электроны и протоны в межмембранное пространство. Особую роль играют Fe содержащие полипептиды. Они различаются между собой в степени сродства к электронам, которое выражается через окислительно-восстановительный потенциал молекулы. Чем выше этот потенциал, тем больше сродство окисленной молекулы к электрону. Электроны переходят по цепи переноса. Их коферменты восстанавливаются и окисляются. Цитохромидаза передает электрон с комплекса цитохрома на молекулярный кислород. Кислород активируется и связывается с электронами. Это важная реакция, в ней кислород приобретает отрицательный заряд, благодаря чему водород соединяется с ним, и образуются вода. Электроны перемещаются в толще внутренней мембраны, при этом через внутреннюю мембрану они не проходят и остаются на внутренней стороне мембраны. Протоны обратно не возвращаются. Они остаются на внешнем мембранном пространстве. На каждую исходную молекулы C6H12O6 64 протона. В результате работы дыхательной цепи на мембранах создается разных электрических потенциалов. Внутренняя сторона мембраны заряжается отрицательно за счет скопления на ней электронов, а наружная сторона внутренней мембраны заряжается положительно. Кроме того, поток протонов создает между двумя сторонами внутренней мембраны разность значения рН. Таким образом, мы имеем кроме мембранного потенциала еще и градиент рН. Вместе они составляют электрохимический протонный градиент. Величина электрохимического протонного градиента, который стимулирует синтез, равно 200 мВ. Заствляет ионы двигаться внутрь матрикса. Заряд – источник протон-движущей силы. Грибовидное тело. Белковый комплекс, который состоит из двух основных частей: ножки (протон-движущий канал) и головки (сопрягающего фактора). Ножка находятся в толще внутренней мембраны, имеет гидрофобную природу и по сути является протон-проводящим канал, по которому протоны из межмембранного пространства возвращаются в матрикс митохондрий Головка тела называется сопрягающий фактор. Здесь располагается фермент АТФ-синтетаза, который превращает энергию движения протона в энергию органического синтеза, синтезируя АТФ из молекулы АДФ и неорганического фосфата (реакция фосфорилирования). АТФ занимает 15% всех белков и образуется при прохождении 1-2 протонов. Действие АТФ-синтетазы обратимо. Субстратами для дыхания служат неорганические соединения. Большинство клеток используют углеводы. Вовлекаются в процесс расщепления только после гидролиза до моносахаридов. Крахмал гидрализуется в глюкозу. Гликоген гидролизуется примерно до 30000 остатков глюкоз. Ферментативное расщепление этих запасных углеводов составляет первый подготовительный этап энергетического обмена. Вторым этапом энергетического обмена является неполное окисление глюкозы. Оно всегда бескислородное. Всегда происходит непосредственно в цитоплазме клетки и называется гликолизом. Гликолиз – неполное бескислородное расщепление глюкозы или многоступенчатый процесс по превращению углерода в 2 молекулы пировиноградной кислоты (С3Н4О3). В ходе гликолиза выделяется большое количество энергии (200 кДж на моль), 60% этой энергии рассеивается в виде тепла, а 40% используется на синтез АТФ. Кроме двух молекул пирувата, образуются протоны, которые в цитоплазме клетки запасаются в форме переносчика. У животных преобладающим переносчиком является НАДФ и соединяется с протоном. Дальнейшая судьба пирувата имеет три пути: 1) Характерный для дрожжей и клеток растений при недостатке кислорода происходит спиртовое брожение и ПВК восстанавливается до этилового спирта (С2Н5ОН). 2) В клетках животных при недостатке кислорода. Накопление в мышечных волокнах молочной кислоты. Пируват восстанавливается до молочной кислоты (С3Н6О3). 3) При наличии в среде кислорода, продукты гликолиза расщепляются до СО2 и Н2О (полное окисление или клеточное дыхание). Продукты гликолиза переходят для дальнейшего расщепления в матрикс митохондрий, где протекает третий этап энергетического обмена – клеточное дыхание. Пируват, одновременно с потоком протонов, направляется внутрь митохондрий. В матриксе молекулы пирувата взаимодействуют с каталитическими белками и преобразуется в молекулу ацетил-коА.Только он может вступить в цикл Кребса. Он вовлекается в цикл трикарбоновых кислот, который представляет собой цепь последовательных реакций, в ходе которых из одной молекулы ацетил-коА образуются молекулы углекислого газа, 2 АТФ и 4 атома водорода. Таким образом, в результате бескислородного окисления и цикла Кребса, одна молекула глюкозы расщепляется до диоксида углерода, который легко выходит из клетки через мембрану и высвобождающееся при реакции энергия расходуется в небольшом количестве на синтез АТФ, а в основном запасается в нагруженных электронами переносчиков. Белки-переносчики транспортируют протоны и электроны к внутренней мембране митохондрий, где передают их на цепь дыхательных ферментов.
Транспорт протонов и электронов приводит к накоплению в протонном резервуаре протонов. Наружная поверхность внутренней мембраны заряжена положительно. Электроны обеспечивают отрицательный заряд внутренней стороны мембраны. Когда разность потенциалов = 200мВ, открывается канал в ножке грибовидного тела, протоны проталкиваются через ножку, отдавая энергию ферменты АТФ-синтетазе, что обеспечивает реакцию фосфорилирования. Синтез АТФ сопряжен с обратным потоком протонов в матрикс. Протонный градиент через внутреннюю мембрану митохондрий дает примерно 24 кДЖ на моль переносимых протонов. И процесс переноса электронов (окисление) в мембранах связан с образованием АТФ из АДФ (реакция фосфорилирования) и поэтому весь называется окислительное фосфорилирования. Ведущую роль в этих реакциях выполняет внутренняя сопрягающая мембрана митохондрий. В процессе окислительного фосфорилирования перенос каждой пары электронов с переносчика на молекулярный кислород доставляет энергию примерно для образования трех молекул АТФ. При окислении одной молекулы глюкозы образуется в реакциях третьего этапа 36 молекул АТФ. Две молекулы АТФ образуются в бескислородном втором этапе обмена и поэтому общий выход = 38 молекул АТФ на одну полностью окисленную молекулу глюкозы. 55% энергии запасается в виде химических связей, 45% энергии рассеивается в виде теплоты. Сопряжение окисления и фосфорилирование известно как хемиосмотическая гипотеза Митчела. В1978 году он был лауреатом нобелевской премии за исследование процесса переноса энергии в клетках и разработку хемиосмотической теории. Эта теория объясняет механизм преобразования энергии в биологической мембране при синтезе АТФ.
Обмен веществ между цитоплазмой и матриксом обеспечивается специальными транспортными системами, локализованными во внутренней мембране митохондрии и обеспечивающие перенос веществ разными механизмами активного транспорта. 1) По механизма типа антипорт, когда два вещества переносятся одновременно в противоположных направлениях. Большая часть АТФ, продуцированная митохондриями, выходит в цитоплазму. Этот перенос осуществляется АТФ-АДФтранслоказой 2) Симпорт или сопряженный транспорт. Таким образом, при участии специфического переносчика осуществляется одновременный перенос пирувата и ионов фосфата и протонов. 3) Унипорт. Облегченная диффузия. Только одно вещество переносится через мембрану в одном направлении (ионы кальция внутрь матрикса). С помощью транспортных белков. Функции митохондрий: 1) Биоэнергетическая; 2) Складирование ионов кальция; 3) Синтез липидов; 4) Место накопления желточных гранул. Митохондрия – полуавтономный органоид. Митохондрии увеличиваются в количестве при делении клеток (особенно в период G2), при увеличении функциональной активности и нагрузки клетки. Увеличение их количества происходит путем роста и деления материнских митохондрий. Делятся митохондрии путем перетяжки, или отпочковывания, или возникновения дочерних митохондрий внутри материнских. В матриксе митохондрий содержится собственная генетическая система, необходимая для процессов воспроизведения. Это митохондриальная ДНК, митохондриальные рибосомы. Гистонов нет. Эти структуры были обнаружены в 1963 году Нассом. Митохондриальная ДНК имеет кольцевую природу. Известно, что все митохондрии содержат несколько копий своего генома. От одной копии до десяти. Линейная длина молекулы варьирует от 5 до 30 мкм. Из нуклеотидных оснований преимущественно гуанин и цитозин. Рибосомы вариабельны, с константой седиментации 55 – 75S. В 1981 году была расшифрована нуклеотидная последовательность митохондриального генома человека. Было показано, что в митохондриях человека содержится 2 гена рРНК, 22 гена тРНК и 13 генов, которые кодируют белки. Репликация и транскрипция происходят в матриксе митохондрий, и в этих процессах преобладает контроль со стороны ядерного генома. Делению или репродукции митохондрий обязательно предшествует репликация митохондриальной ДНК. И этот процесс происходит независимо от репликации ядерной ДНК. Но обычно тогда, когда уже ядерная ДНК вся реплицировалась. Происхождение и эволюцию митохондрий объясняют с позиции эндосимбиотической гипотезы. И эта же самая гипотеза представляет современные митохондрии прямыми потомками бактериального симбионта. Это особая группа фотосинтезирующих бактерий, которые утратили, внедрившись в организм хозяина, способность к фотосинтезу, но сохранили дыхательную функцию. Эти пурпурные бактерии, внедрившись в другую эукариотическую клетку, укоренились к ней. Последние наблюдения цитологов после расшифровки генома показали, что митохондрии эволюционировали вместе со всеми эукариотами от одного общего предка (коэволюция – совместная эволюция). Эволюционно, митохондриальный и ядерные компоненты эукариот возникли одновременно.
Источник: studopedia.info Общие сведения
Эукариотические клетки, которые теряют свои митохондрии, не могут их восстановить. Существуют также эукариоты без них, например, некоторые простейшие. Количество данных органоидов на клеточную единицу обычно составляет от 1000 до 2000 при объемной доле в 25 %. Но эти значения могут сильно варьироваться в зависимости от типа клеточной структуры и организма. В зрелой клетке спермы их около четырех-пяти, в зрелой яйцеклетке – несколько сотен тысяч. Митохондрии передаются через плазму яйцеклетки только от матери, что стало причиной исследования материнских линий. В настоящее время установлено, что также через сперму некоторые мужские органоиды импортируются в плазму оплодотворенной яйцеклетки (зиготы). Вероятно, они будут устранены довольно быстро. Однако есть несколько случаев, когда врачи смогли доказать, что митохондрии ребенка были отцовской линии. Заболевания, вызванные мутациями в митохондриальных генах, наследуются только от матери. Схема строения митохондрииРассмотрим особенности строения этих важных структур. Они образованы в результате сочетания нескольких элементов. Оболочка этих органоидов складывается из внешней и внутренней мембраны, они в свою очередь состоят из фосфолипидных бислоев и белков. Обе оболочки отличаются по своим свойствам. Между ними расположено пять различных отсеков: наружная мембрана, межмембранное пространство (промежуток между двумя мембранами), внутренняя, криста и матрикс (пространство внутри внутренней мембраны), в целом – внутренние структуры органоида. На иллюстрациях в учебниках митохондрия преимущественно выглядит как отдельная бобовидная органелла. Так ли это на самом деле? Нет, они образуют трубчатую митохондриальную сеть, которая может проходить и изменять всю клеточную единицу. Митохондрии в клетке способны сочетаться (путем слияния) и повторно делиться (делением). Внешняя мембрана
Внешняя, которая охватывает всю органеллу и не свернута, имеет весовое отношение фосфолипида к белку 1:1 и, таким образом, похожа на эукариотическую плазматическую мембрану. Она содержит множество интегральных белков, поринов. Порины образуют каналы, которые обеспечивают свободную диффузию молекул с массой до 5000 дальтон через оболочку. Более крупные белки могут вторгаться, когда сигнальная последовательность на N-конце связывается с большой субъединицей белка транслоксазы, из которой они затем активно перемещаются по мембранной оболочке. Если трещины возникают во внешней оболочке, белки из межмембранного пространства могут выходить в цитозоль, что может привести к гибели клетки. Наружная мембрана может сливаться с оболочкой эндоплазматического ретикулума, а затем формировать структуру под названием MAM (ER, ассоциированную с митохондрией). Это важно для обмена сигналами между ER и митохондрией, что также необходимо для переноса липидов. Межмембранное пространство Участок представляет собой промежуток посреди внешней и внутренней мембраны. Поскольку внешняя обеспечивает свободное проникновение малых молекул, их концентрация, таких как ионы и сахар, в межмембранном пространстве идентична концентрациям в цитозоле. Однако для больших белков требуется передача специфической сигнальной последовательности, так что состав белков различается между межмембранным пространством и цитозолем. Таким образом, белок, который удерживается в межмембранном промежутке, является цитохромом. Внутренняя мембранаВнутренняя митохондриальная мембрана содержит белки с четырьмя видами функций:
Внутренняя имеет, в частности, двойной фосфолипид, кардиолипин, замещенный четырьмя жирными кислотами. Кардиолипин обычно характерен для митохондриальных мембран и бактериальных плазматических мембран. В организме человека он в основном присутствует в областях с высокой метаболической активностью или высокой энергетической активностью, таких как сократительные кардиомиоциты, в миокарде.
В типичной митохондрии печени, например, внешняя область, в частности кристы, примерно в пять раз превышает площадь наружной мембраны. Энергетические станции клеток, которые имеют более высокие потребности в АТФ, например, мышечные клетки, содержат больше крист, чем типичная митохондрия печени. Внутренняя оболочка охватывает матрикс, внутреннюю жидкость митохондрии. Он соответствует цитозолю бактерий и содержит митохондриальную ДНК, ферменты цитратного цикла и их собственные митохондриальные рибосомы, которые отличаются от рибосом в цитозоле (но также и от бактерий). Межмембранное пространство содержит ферменты, которые могут фосфорилировать нуклеотиды под потреблением АТФ. Функции
Матрикс Это пространство, включенное во внутреннюю митохондриальную мембрану. Содержит около двух третей общего белка. Играет решающую роль в производстве АТФ с помощью синтазы АТФ, включенной во внутреннюю мембрану. Содержит высококонцентрированную смесь сотен различных ферментов (главным образом, участвующих в деградации жирных кислот и пирувата), митохондриально-специфических рибосом, передаточной РНК и нескольких копий ДНК митохондриального генома. Данные органоиды имеют свой собственный геном, а также ферментативное оборудование, необходимое для осуществления собственного биосинтеза белка. Митохондрия Что такое Митохондрия и её функции Строение и функционирование митохондрий ВыводТаким образом, митохондриями называются клеточные электростанции, которые производят энергию и занимают ведущее место в жизни и выживаемости отдельной клетки в частности и живого организма в целом. Митохондрии – это неотъемлемая часть живой клетки, в том числе растительной, которые до конца еще не изучены. Особенно много митохондрий в тех клетках, которым требуется больше энергии. Источник: uchim.guru
|
 Особенно много находится митохондрий в клетках с высоким энергопотреблением. К ним относятся мышечные, нервные, сенсорные клетки и ооциты. В клеточных структурах сердечной мышцы объемная доля этих органоидов достигает 36 %. Они имеют диаметр около 0.5-1.5 мкм и разнообразные формы, от сфер до сложных нитей. Их число корректируется с учетом энергетических потребностей клетки.
Особенно много находится митохондрий в клетках с высоким энергопотреблением. К ним относятся мышечные, нервные, сенсорные клетки и ооциты. В клеточных структурах сердечной мышцы объемная доля этих органоидов достигает 36 %. Они имеют диаметр около 0.5-1.5 мкм и разнообразные формы, от сфер до сложных нитей. Их число корректируется с учетом энергетических потребностей клетки.