В мембранах эукариот
Взимоотношения между структурой и функцией в клетках прокариотических и эукариотических микроорганизмов.
Эукариотическая клетка (истинноядерная) является сложной структурной единицей у растений, многоклеточных животных, простейших, грибов и всех групп, которые обычно относят к водорослям (кроме цианобактерий).
Прокариотическая клетка (доядерная) является менее сложной структурной единицей бактерий, цианобактерий, актиномицетов.
Бактериальная клетка окружена клеточной стенкой. Цитоплазма обильно насыщена рибосомами. Молекула ДНК обычно расположена в центре клетки. Цитоплазма эукариотической клетки окружена цитоплазматической мембраной (ЦПМ), включает митохондрии, вакуоли, шероховатую эндоплазматическую сеть с рибосомами, гладкую эндоплазматическую сеть, запасные гранулы и ядро.
Для эукариотической клетки характерно наличие внутри нее множества систем элементарных мембран по своей структуре и топологии отличных от плазматической мембраны. Они служат для обособления ряда функциональных компонентов эукариотической клетки в специализированных и частично замкнутых участках, которые обмениваются веществами главным образом путем мембранного транспорта.
Благодаря развитию молекулярной биологии, стало очевидным, что структурные различия между эукариотическими и прокариотическими клетками отражают весьма важные различия в механизмах осуществления ряда жизненных функций клетки. Речь идет, прежде всего, о передаче и проявлении генетической информации, об энергетическом обмене и о механизме поглощения и выделения веществ клеткой.
1. Размер микроорганизмов.
Линейные размеры микроорганизмов в среднем находятся в пределах 0,5-3 мкм, но есть свои гиганты и карлики: н-р клетки нитчатой бактерии Beggiatoa alba имеют диаметр до 500 мкм. Самые мелкие из известных прокариот – микоплазмы, диаметр их клеток 0,1 – 0,15 мкм
У микроорганизмов, в силу их малых размеров, очень велико отношение площади поверхности клетки к ее объему, что создает благоприятные условия для активного обмена с внешней средой. Метаболическая активность микроорганизмов в расчете на единицу биомассы намного выше, чем у более крупных клеток эукариот.
Одной из наиболее существенных особенностей микроорганизмов является высокая пластичность их метаболизма, что приводит к легкости приспособления к меняющимся условиям окружающей среды.
азанное свойство также связано с малыми размерами клеток. Клетки микроорганизмов не могут вместить в себя большое количество белковых молекул. Поэтому не нужные в данных условиях существования ферменты не могут в клетках микроорганизмов содержаться про запас. Они синтезируются только тогда, когда соответствующее питательное вещество (субстрат) появляется в среде. Такие ферменты называются индуцибельными,они могут составлять до 10% общего белка, содержащегося в клетке в данный момент времени. Таким образом, для микроорганизмов характерно большее разнообразие ферментных систем и более мобильные способы регуляции обмена веществ, чем для макроорганизмов.
Другим следствием благодаря высокой пластичности метаболизма микроорганизмов является, по определению В.И.Вернадского, их «всюдность». Их можно обнаружить в арктических областях, в горячих источниках, в высоких слоях атмосферы, в шахтах с высоким содержанием сероводорода и т.д., чем они отличаются от практически всех эукариот.
Мембранные структуры прокариот и эукариот.
Растворенные вещества с небольшими молекулами проникают в эукариотическую клетку через ее поверхностную мембрану. Более крупные молекулы и частицы проникают внутрь путём эндоцитоза. Работа сократительной вакуоли многих простейших, не обладающих клеточными стенками — одна из разновидностей экзоцитоза, который служит активным механизмом осморегуляции. Прокариоты не имеют органелл, выполняющих функцию сократительной вакуоли, и потому не способны активно поддерживать осмотическое равновесие в гипотоничной среде.
связи с этим они могут избежать опасности осмотического лизиса только одним способом — синтезировать достаточно прочную клеточную стенку, способную противостоять тургорному давлению протопласта. Клетки большинства прокариот окружены клеточной стенкой, значительно более толстой, чем мембрана; ее нет только у представителей группы микоплазм. Они чувствительны к осмотическому давлению, и их можно выращивать только в высокоосмотичных средах. Отсутствие клеточной стенки может быть временным явлением, возникающим под влиянием каких-либо внешних факторов (ферменты, антибиотики). Такие бактерии называются L-формы, способны размножаться, образовывать колонии на уплотнённых питательных средах. Или они могут ревертировать в исходную форму.
В состав клеточной стенки прокариот почти всегда входит полимер определенного типа, называемый пептидогликаном (или муреином) и обеспечивающий необходимую механическую прочность. Способность синтезировать полимер такого типа присуща исключительно прокариотам; это одна из биохимических особенностей, отличающая прокариот от эукариот. Исключение составляют архебактерии. У метанобразующих архебактерий клеточная стенка состоит из другого пептидогликана – псевдомуреина. У галобактерий, ацидофильно-термофильных архебактерий и большинства метанобразующих бактерий клеточная стенка построена из белка.
Развитие прочной клеточной стенки также явилось причиной повсеместного распространения микроорганизмов, в том числе в экстремальных условиях обитания.
Цитоплазматическая мембрана прокариот служит гораздо более избирательным барьером между внутренним пространством клетки и внешней средой, чем мембрана эукариот. Самые крупные частицы, способные проходить через этот барьер, имеют размеры молекул: это фрагменты ДНК и белки с относительно низким молекулярным весом (например, внеклеточные ферменты, секретируемые клеткой). Явления экзоцитоза и эндоцитоза совершенно неизвестны у прокариот, даже у тех (группа микоплазм), у которых нет клеточной стенки и поэтому нет механических препятствий для переноса каких-либо частиц или капелек жидкости через поверхность клетки. В результате у прокариот отсутствуют биологические свойства, связанные со способностью к эндоцитозу, в частности способность к внутриклеточному пищеварению и способность, иметь клеточных (невирусных) эндосимбионтов.
У многих прокариот плазматическая мембрана играет определенную роль в энергетическом обмене, чего никогда не бывает в клетках эукариот. У аэробных бактерий дыхательная система переноса электронов «вмонтирована» в клеточную мембрану. У эукариот эта часть механизма дыхания находится во внутренней мембранной системе митохондрий.
Плазматическая мембрана имеет также специальные участки для прикрепления ДНК прокариотической клетки, и именно рост мембраны обеспечивает разделение геномов после завершения их репликации. Это еще одна функция, которую, конечно, никогда не выполняет плазматическая мембрана у эукариот у них разделение геномов происходит путем митоза.
Липидный состав клеточных мембран. Липиды, относящиеся к стеринам, обязательно входят в состав клеточной мембраны эукариот, но не содержатся в значительных количествах в клеточной мембране прокариот, за исключением группы микоплазм. Представители этой группы не способны синтезировать эти вещества, но они включают в клеточную мембрану экзогенные стерины из культуральной среды. Среди жирных кислот, входящих в состав мембранных липидов всех эукариот, имеются полиненасыщенные кислоты (т.е. жирные кислоты, содержащие более одной двойной связи). У большинства прокариот встречаются только насыщенные или мононенасыщенные жирные кислоты; исключение составляют лишь некоторые цианобактерии, способные синтезировать полиненасыщенные жирные кислоты.
Прокариотическая клетка характеризуется отсутствием внутренней компартментализации, создаваемой системами элементарных мембран. Плазматическая мембрана у подавляющего большинства прокариот — единственная мембранная система клетки. Цианобактерии — единственное исключение из этого правила. У этих организмов фотосинтезирующий аппарат находится на уложенных рядами уплощенных мембранных мешках, или тилакоидах, сходных по структуре и функции с тилакоидами хлоропластов. Однако у цианобактерий тилакоиды не заключены в особую органёллу, а лежат непосредственно в цитоплазме.
2. Наследственный материал прокариотов и эукариотов.
В эукариотической клетке, ядро служит основным, но не единственным местом хранения наследственной информации. Небольшая в количественном отношении, но функционально важная часть клеточного генома находится в митохондриях и в хлоропластах (у фотосинтезирующих организмов). ДНК органелл определяет некоторые свойства соответствующих органелл и содержат собственные специфические механизмы транскрипции и трансляции. Митохондрии и хлоропласты, ограниченные мембранами, являются структурами, ответственными за дыхание и фотосинтез (у фотосинтезирующих эукариот).
На электронных микрофотографиях большинства прокариот можно видеть внутри клетки две структурно различающиеся области: цитоплазму и нуклеоплазму. Цитоплазма имеет вид мелкозернистой массы, так как содержит рибосомы. Это всегда так называемые 70S-рибосомы, которые меньше цитоплазматических рибосом эукариот, но сходны по размеру с рибосомами их органелл. Нуклеоплазма имеет неправильные контуры, но оначетко отграничена от цитоплазмы, хотя эти две области никогда не бывают разделены мембраной. Наследственная информацияпрокариотической клетки содержится в нуклеоплазме в структуре, называемой бактериальной хромосомой. Это просто двухцепочечная молекула ДНК, которая имеет кольцевую форму.
Бактериальная хромосома в структурном отношении сходна не с ядерными хромосомами эукариотических клеток, а скорее с ДНК, содержащейся в митохондриях и хлоропластах. Возможно, что эти органеллы произошли от каких-то древних прокариот, которые в процессе эволюции проникли в эукариотическую клетку как симбионты и, в конце концов, утратили способность существовать независимо от организма-хозяина.
Многие бактерии могут также включать небольшие внехромосомные молекулы ДНК кольцевой формы, способные к автономной репликации, так называемые плазмиды. Изученные до сих пор плазмиды несут детерминанты таких фенотипических свойств, как устойчивость к лекарственным веществам и другим антибактериальным препаратам, а также информацию о ферментах некоторых второстепенных метаболических путей. Количество ДНК в плазмиде в 20—1000 раз меньше, чем в бактериальной хромосоме; плазмиды могут быть утрачены клеткой без ущерба для ее жизнеспособности.
Гены прокариот не содержат интронов. Это говорит о том, что прокариоты – это тупиковая ветвь эволюции, так как набор изменений в геноме, так или иначе пределен, наличие интронов у экариот позволяет им подвергаться изменениям бесконечно.
Двигательная активность. Направленное движение цитоплазмы, характерное для большинства эукариотических клеток, у прокариот не наблюдается. Тем не менее, многие прокариоты, обладающие клеточной стенкой, могут активно передвигаться. Одна из разновидностей активного движения скольжение — проявляется только при контакте клетки с твердым субстратом; оно осуществляется без участия каких-либо специальных локомоторных органелл. Скольжение свойственно многим цианобактериям, а также некоторым группам бактерий, не способных к фотосинтезу. Вторая разновидность движения – активное плавание – свойственно клеткам, находящимся в жидкой среде, и осуществляется с помощью жгутиков.
Заключение.
Если прокариоты в течение миллиардов лет развивались сами по себе, то эукариоты никогда не оставались одни. Им приходилось всё время противостоять прокариотам. Они предоставляли последним новые экологические ниши, защиту и были их жертвами. Многоклеточные организмы своими высокоразвитыми и защитными и иными приспособлениями отчасти обязаны агрессивности прокариот. С другой стороны, эукариоты научились извлекать пользу из тесной ассоциации с прокариотами и поставили их себе на службу в качестве эктосимбионтов (в кишечном тракте, рубце, на коже), и эндосимбионтов (для фиксации азота).
Источник: megaobuchalka.ru
Происхождение
Существует три теории происхождения эукариот:
- симбиогенез;
- инвагинагенез;
- химерная теория.
Согласно симбиотической теории происхождения эукариоты возникли путём поглощения прокариот более крупными прокариотами. Этим объясняется нахождение наполовину автономных органелл (содержат ДНК) – митохондрий и пластид.
Инвагинационная теория предполагает, что эукариоты возникли путём впячивания мембраны внутрь прокариотической клетки. Из отделившихся пузырьков сформировались различные органеллы.
Химерное образование эукариот – слияние нескольких прокариот. Слившиеся клетки обменивались генетической информацией.
Мембрана
Снаружи находится плазматическая мембрана эукариотической клетки или плазмалемма, которая осуществляет выборочную взаимосвязь органелл с внешней средой. Поверхностная мембрана имеет жидко-мозаичную структуру, образованную:
- двумя слоями липидов (внешним и внутренним);
- белками (60 % мембраны).
Липиды имеют гидрофильные головки и гидрофобные хвостики, которые обращены внутрь мембраны. Липиды плотно прилегают друг к другу, что обеспечивает мембране эластичность. Жёсткость придаёт встроенный в хвостики холестерин. Липиды защищают и ограничивают клетку.
Белки могут находиться на поверхности мембраны или быть интегрированными в неё.
В зависимости от вида белки осуществляют различные функции:
- транспортную;
- ферментативную;
- рецепторную.
Рис. 1. Строение плазмалеммы.
Органеллы
Структурно-функциональная организация растительной и животной клеток гомологична, т.е. похожа. Однако клетки отличаются специфичными органеллами.
Рис. 2. Строение клеток животных и растений.
Основные компоненты эукариотической клетки и их описание представлены в таблице.
Деление
Основным способом деления эукариот является митоз. Это непрямое деление клетки, включающее две стадии:
- кариокинез – распределение ядерного содержимого между двумя клетками;
- цитокинез – разделение органелл между дочерними клетками.
Деление начинается с удвоения центросомы и распада ядерной мембраны. Из хроматина образуются хромосомы, которые выстраиваются на клеточном экваторе. Прикреплённые микротрубочки веретена деления оттягивают части хромосом в разные стороны, где вокруг них образуется новая ядерная оболочка. Затем распределяются органеллы.
Рис. 3. Митоз.
Источник: obrazovaka.ru
Тип урока: комбинированный.
Методы:
- Словесный
- Наглядный
- Практический
- Проблемно–поисковый
Цели урока:
Образовательная: развивать знания учащихся о строении клеток эукариот и применять их на практических занятих.
Развивающая:
1. Совершенствовать у учащихся умения работать с дидактическим материалом.
2. Развивать мышление учащихся, предлагая задания для сравнения клетки растений и клетки животных с выявлением схожих и отличительных признаков.
Обеспечение урока:
- Плакат “Строение цитоплазматической мембраны”.
- Карточка–задание к уроку “Строение клеток эукариот”.
- Раздаточный материал: (строение прокариотической клетки, типичная растительная клетка, строение животной клетки).
Межпредметные связи: ботаника, зоология, анатомия и физиология человека.
План урока
- Организационный момент 5 мин.
- Проверка готовности к уроку.
- Проверка списочного состава учащихся.
- Сообщение темы и целей урока.
- Изучение нового материала.
- Разделение организмов на про– и эукариоты. (Словесный метод) 10 мин.
- Строение клеток растений и животных. (Самостоятельная работа с использованием раздаточного дидактического материала. Осуществление наглядного, практического и проблемно–поискового методов). 35 мин.
- Строение оболочки клеток (Словесный и наглядный метод). 20 мин.
- Поступление веществ в клетку (Словесный метод) 10 мин.
- Закрепление изученного материала (Словесный метод) 5 мин.
- Домашнее задание 5 мин.
II. Изучение нового материала
Разделение организмов на про – и эукариоты.
По форме клетки необычайно разнообразны: одни как шарики, другие как звездочки со многими лучами, третьи вытянутые и т.д. Различны клетки и по размеру – от мельчайших, с трудом различимых в световом микроскопе, до прекрасно видимых невооруженным глазом (например, икринки рыб и лягушек). Любое яйцо, в том числе гигантские окаменевшие яйца ископаемых динозавров, которые хранятся в палеонтологических музеях, тоже были когда–то живыми клетками. Зато если вести речь о главных элементах внутреннего строения, все клетки схожи между собой [5]
Отличия Прокариот от Эукариот.
- Эукариоты имеют настоящее ядро: генетический аппарат эукариотной клетки защищен оболочкой, схожей с оболочкой самой клетки.
- Включенные в цитоплазму органоиды окружены мембраной.
- Строение клеток растений и животных.
Прокариоты (от лат. Pro–перед, раньше, вместо и греч. karyon– ядро), организмы, клетки которых не имеют ограниченного мембраной ядра–все бактерии, включая архибактерий и цианобактерии. Общее число видов прокариот около 6000. Аналог ядра– структура, состоящая из ДНК, белков и РНК. Генетическая система прокариот (генофор) закреплена на клеточной мембране и соответствует примитивной хромосоме. Размножаются прокариоты без выраженного полового процесса. Прокариоты способны осуществлять ряд физиологических процессов, например, некоторые прокариоты фиксируют молекулярный азот. [1] После вступительной беседы учащиеся рассматривают строение прокариотической клетки, сравнивая основные особенности строения с типами эукариотической клетки. (Рис.2)

Эукариоты – это высшие организмы, имеющие четко оформленное ядро, которое обладает оболочкой (кариомембраной), эта оболочка отделяет его от цитоплазмы. К эукариотам относятся все высшие животные и растения, а также одноклеточные и многоклеточные водоросли, грибы и простейшие. Ядерная ДНК у эукариот заключена в хромосомах. Эукариоты обладают ограниченными мембраной клеточными органоидами. [1]
Клетка любого организма представляет собой систему. Она состоит из 3–х взаимосвязанных между собой частей: оболочки, ядра и цитоплазмы.
При прохождении ботаники, зоологии и анатомии человека вы уже знакомились со строением различных типов клеток, давайте немножко с вами повторим. (Рис.1;задание 1) [6]
Строение и функции органоидов растительных и животных клеток
Таблица заполняется по раздаточному материалу (Рис. 4), (Рис.3).
| Органоиды клетки |
Строение органоидов |
Функция |
Присутствие органоидов в клетках | |
|
растений |
животных |
|||
| Хлоропласт | Представляет собой разновидность пластид. | Окрашивает растения в зеленый цвет, в нем происходит фотосинтез |
+ |
– |
| Лейкопласт | оболочка состоит из двух элементарных мембран, внутренняя из них, врастая в строму, образует немногочисленные тилакоиды. | Окрашивает растения в желтый цвет, синтезирует и накапливает крахмал. |
+ |
– |
| Хромопласт | пластид с жёлтой, оранжевой и красной окраской, окраска обусловлена пигментами – каротиноидами | Бесцветное окрашивание растения |
+ |
– |
| Вакуоль | Занимает до 90 % объема зрелой клетки, заполнена клеточным соком | Функция питания |
+ |
– |
| Микротрубочки | Состоят из белка тубулина, расположены около плазматической мембраны | Участвуют в отложении целлюлозы на клеточных стенках, участвуют в перемещении в цитоплазме различных органоидов. При делении клетки микротрубочки составляют основу структуры веретена деления |
+ |
+ |
| Плазматическая мембрана | Состоит из билипидного слоя, пронизанного белками, погруженными на различную глубину. | Барьер, транспорт веществ, сообщение клеток между собой |
+ |
+ |
| Гладкий ЭПР | Система плоских и ветвящихся трубочек. | Осуществляет синтез и выделение липидов |
+ |
+ |
| Шероховатый ЭПР | Название получил из–за множества рибосом, находящихся на его поверхности | Синтез белков, их накопление и преобразование для выделения из клетки наружу |
+ |
+ |
| Ядро | Окружено двойной ядерной мембраной, имеющей поры. Наружная ядерная мембрана образует непрерывную структуру с мембраной ЭПР. Содержит одно или несколько ядрышек. | Носитель наследственной информации, центр регуляции активности клетки. |
+ |
+ |
| Клеточная стенка | Состоит из длинных молекул целлюлозы, собранных в пучки, называемые микрофибриллами. | Внешний каркас, или защитную оболочку, обеспечивает тургор растительных клеток |
+ |
+ |
| Плазмодесмы | Мельчайшие цитоплазматические каналы, которые пронизывают клеточные стенки. | Объединяют протопласты соседних клеток |
+ |
– |
| Митохондрии | Содержат ферменты для синтеза АТФ. Внутренняя мембрана митохондрий образует многочисленные складки. | Аккумулятор энергии, осуществляет аэробное дыхание. |
+ |
+ |
| Аппарат Гольджи | Состоит из стопки плоских мешочков, называемых цистернами | Синтезирует полисахариды |
+ |
+ |
| Лизосомы | Пузырьки, содержащие концентрированные гидролитические ферменты, которые становятся активными в кислой среде | Участвуют в растворении веществ, попавших в клетку |
+ |
+ |
| Рибосомы | Состоит из двух неравных субъединиц – большой и малой, на которые может диссоциировать. | Место биосинтеза белка |
+ |
+ |
| Эндоцитозный пузырек | Содержит слишком большие молекулы | Содержит слишком большие молекулы, которые не могут проникнуть через мембрану способами диффузии или активного транспорта |
– |
+ |
| Цитоплазма | Состоит из воды с большим количеством растворенных в ней веществ, содержащих глюкозу, белки и ионы. | В ней расположены другие органоиды клетки |
+ |
+ |
| Микрофиламенты | Волокна из белка актина, обычно располагаются пучками вблизи от поверхности клеток. | Играют важную роль в подвижности клеток |
– |
+ |
| Секреторный пузырек | много в клетках, активно синтезирующих вещества, например, в клетках островков Лангерганса | Выносит вещества за пределы клетки |
– |
+ |
| Центриоли | Могут входить в состав митотического аппарата клетки. В диплоидной клетке содержится две пары центриолей. | Участвуют в процессе деления клетки у животных |
– |
+ |
| Пероксисома | Группа пузырьков, известных как микротела | Важна для замедления старения клеток |
– |
+ |
| Микроворсинки | Выступы плазматической мембраны | Увеличивают наружную поверхность клетки, микроворсинки в совокупности образуют кайму клетки |
– |
+ |
Выводы
1. Растительная клетка в своем составе имеет: клеточную стенку, пластиды и вакуоли, присущие только этому типу клеток.
2 . Клеточный центр, центриоли, микроворсинки присутствуют только в клетках животных организмов.
3. Все остальные органоиды характерны как для растительных, так и для животных клеток.
Строение оболочки клеток.
Клеточная оболочка располагается снаружи клетки, отграничивая последнюю от внешней или внутренней среды организма. Ее основу составляет плазмалемма (клеточная мембрана) и углеводно–белковая составляющая, имеющая различную толщину, в зависимости от царства организма (животная или растительная клетка) и от местонахождения клетки в многоклеточном организме. [2]
Оболочка клетки
|
Наружный слой |
Внутренний слой |
|
| У растений называется клеточной стенкой. | У животных называется гликокаликсом. | Называется плазматической мембраной, одинаковый для животных и растений. |
Функции клеточной оболочки:
- Оболочка клетки поддерживает форму клетки и придает механическую прочность как клетке, так и организму в целом
- Защищает клетку от механических повреждений и попадания в нее вредных соединений
- Осуществляет узнавание молекулярных сигналов
- Регулирует обмен веществ между клеткой и средой
- Осуществляет межклеточное взаимодействие в многоклеточном организме.[2]
Функция клеточной стенки:
- Представляет собой внешний каркас – защитную оболочку.
- Обеспечивает транспорт веществ (через клеточную стенку проходит вода, соли, молекулы многих органических веществ).[3]
Наружный слой поверхности клеток животных, в отличие от клеточных стенок растений очень тонкий, эластичный. Он не виден в световой микроскоп и состоит из разнообразных полисахаридов и белков. Поверхностный слой животных клеток называется гликоликсом, выполняет функцию непосредственной связи клеток животных с внешней средой, со всеми окружающими ее веществами, опорной роли не выполняет.
Под гликокаликсом животных и (растительной) клеточной стенкой растений расположена плазматическая мембрана, граничащая непосредственно с цитоплазмой. В состав плазматической мембраны входят белки и липиды. Они упорядоченно расположены и соединены друг с другом химическими взаимодействиями. Молекулы липидов в плазматической мембране расположены в два ряда и образуют сплошной билипидный слой. Молекулы белков не образуют сплошного слоя, они располагаются в слое липидов, погружаясь в него на разную глубину. Молекулы белков и липидов подвижны.[3]
Функция плазматической мембраны:
- она образует барьер, отграничивающий внутреннее содержимое клетки от внешней среды.
- транспорт веществ.
- обеспечивает вязь между клетками в тканях многоклеточных организмов [4]
2.4 Поступление веществ в клетку.
Поверхность клетки не сплошная. В цитоплазматической мембране есть многочисленные мельчайшие отверстия – поры, через которые, с помощью ферментов, внутрь клетки могут проникать ионы и мелкие молекулы. Кроме того, ионы и мелкие молекулы могут попадать в клетку непосредственно через мембрану. Поступление ионов и молекул в клетку – не пассивная диффузия, а активный транспорт, требующий затрат энергии. Транспорт веществ носит избирательный характер. Избирательная проницаемость клеточной мембраны носит название полупроницаемости. [4]
Путем фагоцитоза внутрь клетки поступают: крупные молекулы органических веществ, например белков, полисахаридов, частицы пищи, бактерии. Участие принимает плазматическая мембрана. В том месте, где поверхность клетки соприкасается с частицей какого-либо плотного вещества, мембраны прогибаются, образуют углубление и окружают частицу, которая в “мембранной упаковке” погружается внутрь клетки. Образуется пищеварительная вакуоль, и в ней перевариваются поступившие в клетку органические вещества. [3]
Путем фагоцитоза питаются: амебы, инфузории, лейкоциты животных и человека.
Лейкоциты поглощают бактерии, а также разнообразные твердые частицы случайно попавшие в организм, защищая его таким образом от болезнетворных частиц. Клеточная стенка растений, бактерий и сине–зеленых водорослей препятствует фагоцитозу, и потому этот путь поступления веществ в клетку у них, практически, отсутствует.
Через плазматическую мембрану в клетку проникают и капли жидкости, содержащие в растворенном и взвешенном состоянии разнообразные вещества.
Поглощение жидкости в виде мелких капель напоминает питье, и это явление было названо пиноцитозом. Процесс поглощения жидкости сходен с фагоцитозом. Капля жидкости погружается в цитоплазму в “мембранной упаковке”. Органические вещества, попавшие в клетку вместе с водой, начинают перевариваться под влиянием ферментов, содержащихся в цитоплазме. Пиноцитоз широко распространен в природе и осуществляется клетками всех животных организмов. [3]
III. Закрепление изученного материала.
- На какие две большие группы разделяются все животные организмы по строению ядра?
- Какие органоиды свойственны только растительным клеткам?
- Какие органоиды свойственны только животным клеткам?
- Чем различается строение оболочки клеток растений и животных?
- Два способа поступления веществ в клетку?
- Значение фагоцитоза для животных?
Список использованной литературы:
- Большой энциклопедический словарь “Биология”, под редакцией М.С. Гилярова, Научное издательство “Большая Российская Энциклопедия”, Москва 1998
- Е. Тупикин “Общая биология с основами экологии и природоохранной деятельности”, Москва ПроОбрИздат, 2001
- Ю.И. Полятинский “Общая биология для 9–10 классов средней школы”
- Захаров В.Б. “Общая биология для 10–11 классов”, Москва “Дрофа”, 2003
- “Энциклопедия для детей, Биология, том 2”, Москва, “Аванта +”, 1999
- Р.А. Петросова “Дидактический материал по общей биологии”, Минск ООО “Белфарпост”, 1997
Источник: urok.1sept.ru
Происхождение эукариот «наизнанку»
В недавно опубликованной в журнале BMC Biology статье [4] Дэвид и Базз Баумы, основываясь на большом количестве филогенетических данных, выдвинули новую гипотезу происхождения эукариотической клетки. Они называют эту гипотезу «моделью наизнанку» (inside-out, изнутри — наружу), в противовес господствовавшей до сих пор гипотезе «снаружи — внутрь» (outside-in). Согласно традиционной теории мембранные органеллы эукариот появились благодаря «впячиванию» своей наружной мембраны. Митохондрии, например, согласно этой гипотезе, были «проглочены» будущими эукариотами с помощью фагоцитоза. Однако со времени появления этой гипотезы накопилось немало данных, которые ей противоречат и указывают на то, что ситуация была противоположной. Вероятно, новые органеллы появились у будущих эукариот более дружелюбным способом — с помощью объятий. «Модель наизнанку» предполагает, что эукариотическое ядро образовалось из основной части предковой клетки, а цитоплазма с митохондриями и другими мембранными органеллами — из выростов этой клетки, которые по началу просто окружали клетки-симбионты (рис. 1). Новую гипотезу поддерживает множество важных фактов. Например, археи (они и были этими предковыми клетками) могут только выпячивать мембрану, а «впячивать» — нет. Несомненно, эта новая гипотеза требует дальнейшей проработки, но специалисты* оценивают ее позитивно: она действительно подтверждается известными данными о морфологии и биохимии прокариот и помогает сделать предсказания, которые можно проверить экспериментально (например, механизмы сборки ядерных пор и филогению белков фагоцитоза).
* — более развернутый комментарий специалиста о «модели наизнанку» можно прочитать на ПостНауке: «Выдвинута новая гипотеза происхождения эукариотической клетки» [5]. — Прим. автора.
Математическое моделирование позволило другой группе ученых лучше разобраться с еще одним важным вопросом: какой была мембрана общего предка архей и бактерий, и как ее строение определило эволюцию этих двух групп прокариот. Об этом рассказывается в их недавней статье, вышедшей в журнале PLoS Biology [6].
Бактерии и археи: единство противоположностей
Все современные живые организмы относятся к одному из трех доменов жизни: бактерии, археи и эукариоты. По более-менее общепринятой гипотезе эукариоты происходят от своеобразного «слияния» двух других групп, которые являются гораздо более древними. Бактерии и археи происходят от общего предка — по-английски он называется LUCA (last universal common ancestor, последний универсальный общий предок). Бактерии и археи имеют много общих черт, включая одинаковый генетический код, механизмы транскрипции и рибосомной трансляции, но при этом отличаются в некоторых ключевых моментах. Они имеют разный химический состав клеточных мембран и стенок, по-разному устроенный гликолиз, ионные насосы и даже разные механизмы репликации ДНК.
Возможно, различия в устройстве клеточной мембраны являются ключевыми в этом списке различий (рис. 2) [7]. Мембраны современных бактерий состоят из фосфолипидов: сложных эфиров глицерина, двух остатков жирной кислоты и одного фосфатного остатка, к которому может быть присоединена дополнительная полярная группа. Гидрофобные хвосты жирных кислот образуют средний слой мембраны, а полярные остатки глицерина, фосфата и вспомогательных полярных групп — наружный и внутренний слои. Мембраны архей устроены в принципе похоже, но на другой химической основе. Вместо жирных кислот их липиды содержат терпеновые спирты, углеводородные цепочки которых несут метильные группы через каждые четыре атома. Моделирование молекулярной динамики мембран показало, что благодаря таким метильным «ответвлениям» мембраны становятся очень прочными, но при этом сохраняют гибкость [8, 9]. Терпеновые спирты простыми эфирными связями присоединяются к глицеринфосфату, фосфатный остаток может дополняться другими полярными головками, такими же, как у бактерий. Сам глицеринфосфат архей тоже отличается от бактериального — у архей используется другой его оптический изомер (глицерин-1-фосфат вместо глицерин-3-фосфата). Получается, что мембрана — важнейший элемент, обеспечивающей существование клетки как самостоятельной единицы, — появилась у бактерий и архей независимо. Из этого удивительного наблюдения некоторые ученые даже делают вывод о том, что у LUCA мембраны вообще не было [10]. Но это крайне маловероятно, учитывая, насколько важной для большинства биохимических процессов является мембрана. Сложно представить, что молекулярные механизмы, протекающие одинаково и у бактерий, и у архей, появились и могли функционировать еще до появления мембраны. Значит, какая-то мембрана у LUCA все-таки была. Группа ученых из Университетского Лондонского колледжа с помощью математического моделирования разработала модель, описывающую, как эта мембрана выглядела, и как из нее появились разные мембраны бактерий и архей [6].
«Протекающая» мембрана
C различным строением мембраны бактерий и архей никак не вязалось то, что производство энергии в клетках обеих групп устроено очень похожим образом. Дело в том, что во всех современных клетках производство энергии (которая запасается в виде молекул АТФ) сопряжено с мембраной. Ключевыми стадиями этого процесса являются создание градиента протонов на мембране (избыток ионов Н+ с наружной стороны мембраны по сравнению с внутренней) и работа АТФ-синтазы за счет этого градиента. При этом протоны проходят через канал в АТФ-синтазе, вызывая тем самым механический поворот части АТФ-ситназного комплекса, который, в свою очередь, обеспечивает катализ синтеза АТФ. Согласно филогенетическим исследованиям, АТФ-синтазы всех организмов имеют общее эволюционное происхождение, и предковая молекула была уже у LUCA. У некоторых бактерий и архей вместо градиента протонов используется градиент ионов натрия, а у некоторых — и тот, и другой. Долгое время считалось, что Na+ выступает в качестве заменителя H+ у организмов, живущих в экстремальных условиях (термальных источниках или в сильнощелочной среде). Однако оказалось, что натрий-специфические ферменты занимают самые нижние ветви филогенетического древа в обоих доменах, что указывает на их древность. Модель функционирования древней мембраны, предложенная британскими учеными, успешно объясняет, как и зачем в процессе эволюции возникла способность АТФ-синтазы использовать ионы натрия. Но, прежде чем ответить на этот вопрос, они должны были разобраться с еще одной проблемой — несмотря на общее происхождение АТФ-синтаз, ионные насосы возникли у бактерий и архей независимо, т.е, вероятно, у LUCA их не было. Как же тогда древняя клетка могла избавляться от протонов, поступающих внутрь при работе АТФ-синтазы, и создавать градиент протонов?
По мнению авторов исследования, единственным объяснением могло быть то, что мембрана LUCA была «протекающей» (leaky), и клетка использовала естественные источники протонного градиента. На основе своих предположений ученые построили математическую модель древней клетки. В этой модели клетка находится на границе между двумя ламинарными потоками — кислотным (pH
По мнению ученых, такая «протекающая» мембрана могла состоять из смеси амфифильных молекул, включая жирные кислоты и изопрены, но никак не могла содержать фосфолипиды, свойственные современным мембранам. Добавление фосфолипидов приводит к снижению проницаемости мембраны для ионов, так как полярные группы не могу проходить через неполярную внутреннюю часть мембраны. Такая мембрана не позволяла бы поддерживать градиент протонов, а значит, и работу АТФ-синтазы. Получается, что для клеток с «протекающей» мембраной не нужны ни фосфолипиды, ни ионные насосы (они никак не буду способствовать более эффективной работе АТФ-синтазы, т.к. все «накачанные» ионы будут утекать через мембрану). Чтобы понять, как произошел переход от «протекающей» мембраны к современным мембранам с ионными насосами, ученые обратились к уже упомянутому факту: некоторые АТФ-синтазы могут использовать не только протоны, но и ионы натрия.
Исследователи предположили, что необходимым шагом для перехода к современной мембране было появление способности использовать для создания энергии градиента ионов натрия. Создавать такой градиент могла бы молекула SPAP (sodium-proton antiporter, антипорт для ионов натрия и протонов), которая переносит один ион натрия в обмен на один протон. SPAP есть у многих представителей как архей, так и бактерий. Именно эта молекула могла бы использовать естественный градиент протонов для создания градиентов ионов натрия. Даже «протекающая» мембрана в шесть раз менее проницаема для ионов натри, чем для протонов, поэтому градиент ионов натрия гораздо более долговечен в таких условиях. Если АТФ-синтаза сможет использовать для производства АТФ и протоны, и ионы натрия, клетка, согласно подсчетам, сможет создавать на 60% больше энергии. Как уже было отмечено, некоторые современные АТФ-синтазы действительно способны использовать оба вида ионов. Другие используют только один тип ионов, но при этом все они отличаются только парой аминокислотных замен (вероятно, это связано со схожестью ионного радиуса и заряда ионов Na+ и H3O+ — форм, в которые этих ионы обычно транспортируются ионными каналами). Получившийся благодаря SPAP и смешанной работе АТФ-синтаз выигрыш в энергии клетки смогли бы использовать для того, чтобы начать занимать новые экологические ниши, в которых естественный градиент протонов был гораздо ниже (до 50 раз ниже) или был непостоянным. Кроме того, наличие SPAP делает выгодным наличие в клетке ионных насосов. Согласно расчетам модели, преимущество в использовании насосов возрастает со снижением проницаемости мембраны, вплоть до значений проницаемости, характерных для современных мембран.
Получатся, что SPAP — это та молекула, которая могла бы обеспечить переход от «протекающей» мембраны к почти непроницаемой современной, параллельно позволяя древним клеткам расширять ареал своего обитания. По мере расселения, в разных популяциях LUCA могли возникать различные типы насосов, поэтому в современном мире бактерий и архей мы наблюдаем такое разнообразие молекул, причем не все они имеют общее происхождение. Исследователи смогли ответить и на вопрос, связанный с принципиальным различием мембран бактерий и архей. Моделирование показало, что только после появления в эволюции ионных насосов клеткам стало выгодно снижать проницаемость мембраны за счет присоединения гидрофильных глицерол-фосфатных головок. Из-за того, что такой синтез фосфолипидов может происходить двумя путями, в зависимости от того, с какой стороны происходит нуклеофильная атака на карбонильный центр, появилось два разных хиральных варианта фосфолипидов у бактерий и архей. Получается, что разные популяции получили разные ионные насосы, а потом каждая из них пошла либо по «архейному» пути, либо по «бактериальному», в зависимости от реакции нуклеофильного замещения.
Заключение
Изучать появление и эволюцию жизни на самых ранних ее этапах — задача сложная и нетривиальная, требующая работы с большими объемами данных и особенных подходов. В последние годы у ученых в руках появляется все больше инструментов для таких исследований, позволяющих им проверять давно сформулированные гипотезы и выдвигать новые предположения. Иногда результаты удивляют и предполагают отказ от уже устоявшихся и давно вошедших в учебники теорий. Одно из новых исследований, например, показало, что стоит отказаться от теории происхождения мембранных органелл путем фагоцитоза, а обратить внимание на противоположную модель — модель расширения мембраны. Другое описанное в этой статье исследование предлагает еще одну достаточно революционную идею. Согласно математической модели британских ученых мембрана LUCA была «протекающей», а переход к современной мембране стал возможен благодаря антипорту протонов и ионов натрия. Эта модель подразумевает, что мембрана древних клеток состояла из жирных кислот и терпенов, хотя ранее такие мембраны считались неподходящими для производства энергии как раз из-за своей склонности к «протечкам».
Благодаря развитию информационных технологий и растущим объемам биологических баз данных ученые могут, хотя только в компьютерных моделях, заглянуть в далекое прошлое. Являются ли эти модели верными, покажут дальнейшие исследования, но уже сейчас они помогают понять многие критические точки в эволюции жизни на Земле.
Источник: biomolecula.ru
Мембранные органоиды клетки. Ядро. Прокариоты и эукариоты
Необходимо запомнить
ВАЖНО!
Органоиды клетки
Органоиды, или Органеллы, – постоянные специфические структуры цитоплазмы, выполняющие определённые функции, необходимые для поддержания жизнедеятельности клетки.
Различают органоиды общего значения и специальные органоиды. Органоиды общего значения имеются во всех клетках и выполняют общие функции. Это – митохондрии, рибосомы, эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы, цитоскелет и клеточный центр.
Органоиды специального значения имеются только в клетках какого-то определённого типа и обеспечивают выполнение функций, присущих только этим клеткам.
Мембранные органоиды:
— ядро;
— эндоплазматическая сеть;
— аппарат Гольджи;
— митохондрии;
— лизосомы;
— пластиды;
— вакуоли.
Эндоплазматическая сеть (ЭПС) открыта К. Портером в 1945 году. ЭПС или ЭПР (эндоплазматический ретикулум) – сеть канальцев и цистерн, сложенных мембранами. Различают гранулярную (шероховатую, зернистую) и гладкую (агранулярную) ЭПС.
Гранулярная ЭПС содержит рибосомы на наружной стороне мембраны. Гладкая ЭПС не содержит рибосомы. В скелетных мышцах ЭПС носит название саркоплазматический ретикулум. ЭПС пронизывает всю клетку. Полость ЭПС сообщается с ядром и цитоплазматической мембраной.
На рибосомах гранулярной ЭПС синтезируются секреторные белки, предназначенные для выведения из клетки, а также белки лизосом и внеклеточного матрикса.
Наряду с секреторными белками на гранулярной ЭПС синтезируется большая часть полуинтегральных и интегральных белков. В гладеой ЭПС происходит также синтез мембраны липидов и осуществляется «сборка» компонентов мембраны.
Кроме того, ЭПС, как считают, участвует в образовании пероксисом. Таким образом, гранулярная ЭПС служит «фабрикой» мембран для плазмалеммы, аппарата Гольджи, лизосом и других мембранных структур клетки.
Агранулярная (гладкая) эндоплазматическая сеть представляет собой замкнутую сеть трубочек, канальцев, цистерн. На цитоплазматической поверхности гладкой ЭПС синтезируются жирные кислоты, большая часть липидов клетки, в том числе почти все липиды, необходимые для построения клеточных мембран. Поэтому гладкую ЭПС нередко называют «фабрикой липидов». Например, в клетках печени с мембранами гладкого эндоплазматического ретикулума связан фермент, обеспечивающий образование глюкозы из глюкозо-6-фосфата. Эта реакция имеет большое значение в поддержании уровня глюкозы в организме человека.
В организме человека эндоплазматическая сеть особенно хорошо развита в клетках, синтезирующих гормоны, в клетках печени.
Комплекс Гольджи (КГ, или аппарат Гольджи, – пластинчатый комплекс, расположен вблизи ядра, между ЭПС и плазмалеммой. Его структурно-функциональная единица – диктиосома – представляет собой стопку из 5–20 плоских одномембранных мешочков (цистерн), имеющих диаметр около 1 мкм, внутренние полости которых не сообщаются друг с другом. Количество таких мешочков в стопке обычно не превышает 5–20, а расстояние между ними составляет 20–25 нм.
Белки, синтезированные на шероховатой эндоплазматической сети, попадают в аппарат Гольджи. Здесь осуществляется химическая модификация транспортируемых белков и их упаковка в специальные пузырьки.
Таким образом, основными функциями комплекса Гольджи являются химическая модификация, накопление, сортировка, упаковка в секреторные пузырьки и транспорт по назначению белков и липидов, синтезированных в ЭПС.
В комплексе Гольджи образуются лизосомы и синтезируются некоторые полисахариды.
Лизосомальная система и пероксисомы
Лизосомы – мембранные органеллы клеток животных и грибов, содержащие гидролитические ферменты и осуществляющие гидролитическое расщепление макромолекул (внутриклеточное пищеварение). Лизосомы представляют собой окружённые одинарной мембраной пузырьки, размеры которых в клетках животных колеблются от 0,2 до 0,5 мкм. В лизосомах содержится не менее 60 гидролитических ферментов, которые расщепляют все основные классы органических макромолекул.
Все ферменты лизосом активны лишь в кислой среде при значениях pH, близких 5,0. Количество лизосом в разных клетках варьирует от единичных до нескольких сотен, как например, в фагоцитах.
Завершающие этапы процесса внутриклеточного переваривания веществ, поглощённых клеткой, осуществляются в лизосомах.
Лизосомы с помощью своих ферментов могут разрушать не только отдельные органеллы или клетки, но и целые органы (автолиз). Например, в процессе онтогенеза лягушки с помощью ферментов лизосом лизируются хвост и жабры головастика, а образующиеся при этом продукты распада используются для формирования органов взрослого животного.
Митохондрии – крупные мембранные органоиды клетки, которые можно различить в световой микроскоп. Митохондрии присутствуют во всех эукариотических клетках человека, кроме эритроцитов.
Они имеют обычно округлую, удлиненную или нитевидную формы. Количество митохондрий в клетке колеблется в широких пределах (от 1 до 100 тыс. и более) и зависит от потребностей клетки в энергии. Митохондрии имеют наружную и внутреннюю мембраны.
На внутренней поверхности увеличенного фрагмента кристы видны небольшие выпуклости, обращенные в митохондриальный матрикс, которые содержат ферментные системы, обеспечивающие процессы дыхания. Наружная мембрана гладкая и по своему составу сходна с плазмалеммой.
В матриксе содержатся кольцевая молекула митохондриальной ДНК (мтДНК), различные включения, а также молекулы мРНК, транспортной РНК (тРНК) и рибосомы, сходные по строению с рибосомами бактерий. Здесь же располагаются ферменты, превращающие пируват и жирные кислоты в ацетил-КоА, и ферменты реакций цикла Кребса.
Митохондриальная ДНК имеет не линейную, как в хромосомах ядра, а кольцевую форму. Главная функция митохондрий – синтез АТФ, основного источника энергии для обеспечения жизнедеятельности клетки. Поэтому митохондрии называют «энергетическими станциями» клетки.
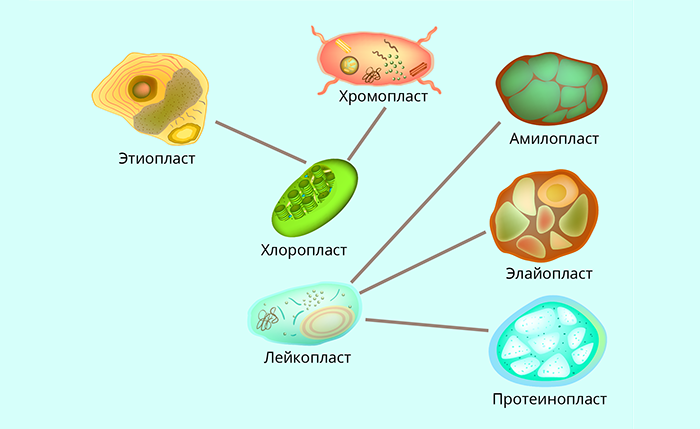
Пластиды
Пластиды – это органоиды клеток растений и некоторых фотосинтезирующих простейших. У большинства животных и грибов пластид нет.
Пластиды делятся на несколько типов: хлоропласты, хромопласты, лейкопласты. Наиболее важный и известный – хлоропласт, содержащий зелёный пигмент хлорофилл, который обеспечивает процесс фотосинтеза.
Все виды пластид связаны между собой общим происхождением или возможным взаимопревращением. Пластиды развиваются из пропластид – более мелких органоидов меристематических клеток.
Строение пластид
Пластиды относятся к двумембранным органоидам, у них есть внешняя и внутренняя мембраны.
Во многих пластидах, особенно в хлоропластах, хорошо развита внутренняя мембранная система, формирующая такие структуры, как тилакоиды, граны (стопки тилакоидов), ламелы – удлинённые тилакоиды, соединяющие соседние граны. Внутреннее содержимое пластид обычно называют стромой. В ней, помимо прочего, находятся крахмальные зёрна.
Считается, что в процессе эволюции пластиды появились аналогично митохондриям – путём внедрения в клетку-хозяина другой прокариотической клетки, способной в данном случае к фотосинтезу. Поэтому пластиды считают полуавтономными органеллами. Они могут делиться независимо от делений клетки, у них есть собственная ДНК, РНК, рибосомы прокариотического типа, т. е. собственный белоксинтезирующий аппарат. Часть генов, управляющая их функционированием, находится как раз в ядре.
Ядро
Ядро – важнейшая часть эукариотической клетки. Оно состоит из ядерной оболочки, кариоплазмы, ядрышек, хроматина.
1. Ядерная оболочка по строению аналогична клеточной мембране, содержит поры. Ядерная оболочка защищает генетический аппарат от воздействия веществ цитоплазмы. Осуществляет контроль за транспортом веществ.
2. Кариоплазма представляет собой коллоидный раствор, содержащий белки, углеводы, соли, другие органические и неорганические вещества. В кариоплазме содержатся все нуклеиновые кислоты: практически весь запас ДНК, информационные, транспортные и рибосомальные РНК.
3. Ядрышко – сферическое образование, содержит различные белки, нуклеопротеиды, липопротеиды, фосфопротеиды. Функция ядрышек – синтез зародышей рибосом.
4. Хроматин (хромосомы). В стационарном состоянии (время между делениями) ДНК равномерно распределены в кариоплазме в виде хроматина. При делении хроматин преобразуется в хромосомы.
Функции ядра: в ядре сосредоточена информация о наследственных признаках организма (информационная функция); хромосомы передают признаки организма от родителей к потомкам (функция наследования); ядро согласует и регулирует процессы в клетке (функция регуляции).
Одномембранные органоиды клетки
Геном, генотип и кариотип
Источник: resh.edu.ru