Схема развития животного мира
Ход и направление процесса возникновения видов в соответствии с основными положениями теории эволюции Ч. Дарвина подкрепляются данными из различных разделов биологии, в том числе данных из области палеонтологии, которые служат вещественными доказательствами, так как основаны на изучении ископаемых остатков некогда живших организмов. В результате поступательного развития жизни на смену одним группам организмов приходили другие, в то время как третьи изменялись мало, а четвертые вымирали. На основе находок ископаемых форм в отложениях земных пластов удается проследить подлинную историю живой природы. Так были созданы палеонтологические ряды лошади (В. О. Коралевский), слона, некоторых птиц, моллюсков и др.- от наиболее примитивных исходных форм до современных их представителей. Применение радиоизотопного метода позволяет с большой точностью определить возраст пород в местах залегания палеонтологических остатков и возраст ископаемых организмов.
На основе данных палеонтологии всю историю жизни на Земле подразделяют на эры и периоды.
Таблица 1. Геохронологическая шкала
| Эры | их длительность, млн. лет | Животный и растительный мир |
| название и длительность, млн. лет | возраст, млн. лет |
|
| Кайнозойская (новой жизни) 60-70 | 60-70 | ген 1,5-2 Животный и растительный мир принял современный облик |
| Верхнетретичный (неоген) 25 Нижнетретичный (палеоген) 41 Господство млекопитающих, птиц. Появление лемуров и дол-гопятов — низкоорганизованных приматов, позднее — парапитеков, дриопитеков. Расцвет насекомых. Продолжается вымирание крупных пресмыкающихся. Исчезают многие группы головоногих моллюсков. Господство покрытосеменных растений. Сокращение флоры голосеменных растений |
||
| Мезозойская (средней жизни) 173 | 240±10 | Меловой 70 Юрский 58 Триасовый 45 Появление высших млекопитающих и настоящих птиц, хотя еще распространены зубастые птицы. Преобладают костистые рыбы. Резко сокращается численность папоротников и голосеменных. Появление и распространение покрытосеменных Господство пресмыкающихся. Появление археоптерикса. Расцвет головоногих моллюсков. Господство голосеменных Начало расцвета пресмыкающихся. Появление первых млекопитающих, настоящих костистых рыб. Исчезновение семенных папоротников |
| Палеозойская (древней жизни) 330
|
570
|
Пермский 45 Каменноугольный (карбон) 55-75 Быстрое развитие пресмыкающихся. Возникновение звероподобных пресмыкающихся. Вымирание трилобитов. Исчезновение лесов каменноугольного периода. Появление и развитие голосеменных. Расцвет земноводных. Возникновение первых пресмыкающихся. Появление скорпионов, пауков, летающих форм насекомых. Сокращение численности трилобитов. Развитие высших споровых и семенных папоротников. Преобладание древних плаунов и хвощей. Развитие грибов |
| Девон ский 50-70 Расцвет щитковых. Появление кистеперых рыб и стегоцефалов. Возникновение грибов. Развитие, а затем вымирание псилофитов. Распространение на суше высших споровых |
||
| Силурий ский 30 Пышное развитие кораллов, три |
||
| Ордовик- ский 60 Расцвет морских беспозвоночных, трилобитов, моллюсков, археоциат . |
||
| Кемб- рийский 70 Широкое распространение водорослей |
||
| Протеро зойская (ран ней жизни) 2000 |
2600 + 100 |
Представлены все типы беспозвоночных. Появление первичных хордовых — подтипа бесчерепных |
| Архейская (самая древ няя) 900 |
3500 | Следы жизни незначительны. Обнаружены остатки бактерий и одноклеточных водорослей |
1. Архейская эра — древнейший этап в истории Земли, когда в водах первичных морей возникла жизнь, которая была представлена первоначально доклеточными ее формами и первыми клеточными организмами. Aнализ осадочных пород этого возраста показывает, что в водной среде обитали бактерии и синезеленые.
2. Протерозойская эра. На грани архейской и протерозойской эры произошло усложнение строения и функции организмов: возникли многоклеточность, половой процесс, который усилил генетическую неоднородность организмов и дал обширный материал для отбора, более разнообразными стали фотосинтезирующие растения. Многоклеточность организмов сопровождалась повышением специализации клеток, их объединением в ткани и функциональные системы.
Проследить в деталях эволюцию животных и растений в протерозойскую эру довольно трудно из-за перекристаллизации осадочных пород и уничтожения органических остатков. В отложениях этой эры обнаружены лишь отпечатки бактерий, водорослей, низших типов беспозвоночных и низших хордовых. Крупным шагом в эволюции было появление организмов с двусторонней симметрией тела, дифференцированного на передний и задний отделы, левую и правую стороны, выделение спинной и брюшной поверхности. Спинная поверхность у животных служила защитой, а на брюшной располагались рот и органы захвата пищи.
3. Палеозойская эра. Животный и растительный мир достиг большого разнообразия, стала развиваться наземная жизнь.
В палеозое различают шесть периодов: кембрийский, ордовикский, силурийский, девонский, каменноугольный, пермский. В кембрийском периоде жизнь была сосредоточена в воде (она покрывала значительную часть нашей планеты) и представлена более совершенными многоклеточными водорослями, имевшими расчлененное слоевище, благодаря которому они активнее синтезировали органические вещества и явились исходной ветвью для наземных листостебельных растений. Широкое распространение в морях получили беспозвоночные, в том числе плеченогие моллюски, а из членистоногих — трилобиты. Самостоятельным типом двухслойных животных того периода были археоциаты, формировавшие рифы в древних морях.
и вымерли, не оставив потомков. На суше обитали лишь бактерии и грибы.
В ордовикском периоде климат был теплым даже в Арктике. В пресных и солоноватых водах этого периода пышного развития достигли планктонные водоросли, разнообразные кораллы из типа кишечнополостных, существовали представители почти всех типов беспозвоночных в том числе трилобиты, моллюски, иглокожие. Широко представлены были бактерии. Появляются первые представители бесчелюстных позвоночных — щитковые.
В конце силурийского периода в связи с горообразовательными процессами и сокращением площади морей часть водорослей оказалась в новых условиях среды — в мелких водоемах и на суше. Многие из них погибли. Однако в результате разнонаправленной изменчивости и отбора отдельные представители приобрели признаки, способствовавшие выживанию в новых условиях. Появились первые наземные споровые растения — псилофиты. Они имели цилиндрический стебель около 25 см высоты, вместо листьев — чешуйки. Важнейшие приспособления у них — возникновение покровной и механической тканей, корнеподобных выростов — ризоидов, а также элементарной проводящей системы.
В девоне численность псилофитов резко сократилась, на смену им пришли их преобразованные потомки, высшие растения — плауновидные, моховидные и папоротниковидные, у которых развиваются настоящие вегетативные органы (корень, стебель, лист).
зникновение вегетативных органов повысило эффективность функции отдельных частей растений и их жизненность как гармонически целостной системы. Выход на сушу растений предшествовал выходу животных. На Земле растения накапливали биомассу, а в атмосфере — запас кислорода. Первыми обитателями суши из беспозвоночных были пауки, скорпионы, многоножки. В девонских морях было много рыб, среди них — челюстные панцирные, имевшие внутренний хрящевой скелет и внешний прочный панцирь, подвижные челюсти, парные плавники. Пресные водоемы населяли кистеперые рыбы, у которых было жаберное и примитивное легочное дыхание. С помощью мясистых плавников они перемещались по дну водоема, а при пересыхании переползали в другие водоемы. Группа кистеперых рыб явилась предками древних земноводных — стегоцефалов. Стегоцефалы обитали в болотистой местности, выходили на сушу, но размножались только в воде.
В каменноугольном периоде распространились гигантские папоротникообразные, которые в условиях теплого влажного климата расселились повсеместно. В этот период достигли расцвета древние земноводные.
В пермский период климат стал более сухим и холодным, что привело к вымиранию многих земноводных.
концу периода число видов земноводных стало резко сокращаться, и до наших дней сохранились лишь мелкие земноводные (тритоны, лягушки, жабы). На смену древовидным споровым папоротникообразным пришли семенные папоротники, давшие начало голосеменным растениям. Последние имели развитую стержневую корневую систему и семена, оплодотворение у них проходило в отсутствие воды. Вымерших земноводных сменила более прогрессивная группа животных, произошедшая от стегоцефалов,- пресмыкающиеся. У них были сухая кожа, более плотные ячеистые легкие, внутреннее оплодотворение, запас питательных веществ в яйце, защитные яйцевые оболочки.
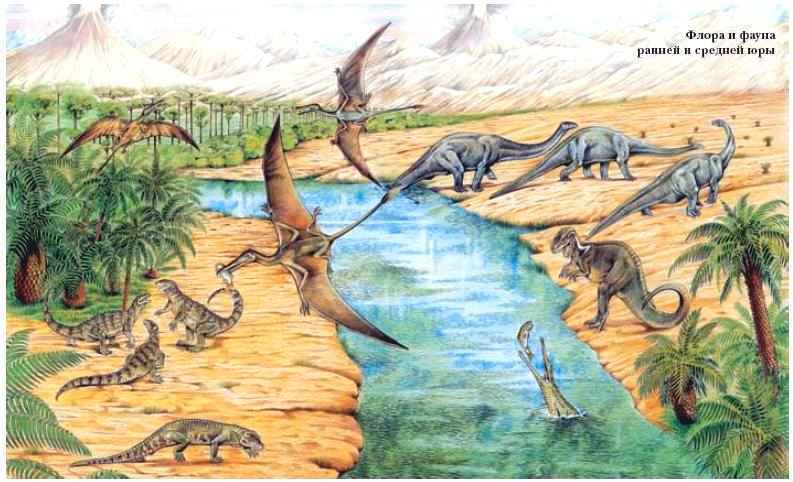
4. Мезозойская эра включает три периода: триасовый, юрский, меловой.
В триасе широко распространились голосеменные растения, особенно хвойные, занявшие господствующее положение. Одновременно широко расселились пресмыкающиеся: в морях обитали ихтиозавры, плезиозавры в воздухе — летающие ящеры, разнообразно были пpeдставлены пресмыкающиеся и на земле. Гигантские пресмыкающиеся (бронтозавры, диплодоки и др.) вскоре вымерли. В самом начале триаса от пресмыкающихся отделилась группа мелких животных с более совершенным строением скелета и зубов. Эти животные npиобрели способность к живорождению, постоянную температуру тела, у них было четырехкамерное сердце и целый ряд других прогрессивных черт организации. Это были первые примитивные млекопитающие.
В отложениях юрского периода мезозоя o6наружены также останки первоптицы — археоптерикса. Он сочетал в своем строении признаки птиц и пресмыкающихся.
В меловом периоде мезозоя от голосеменных отделилась ветвь растений, имевших орган семенного размножения — цветок. После оплодотворения завязи цветка превращается в плод, поэтому развивающиеся семена внутри плода защищены мякотью и оболочками от неблагоприятных условий среды. Многообразие цветков различных приспособлений для опыления и распространения плодов и семян позволило покрытосеменным (цветковым) растениям широко распространиться в природе и занять господствующее положение. Параллельно с ними развивалась группа членистоногих — насекомых которые, будучи опылителями цветковых растений в большой мере способствовали их прогрессивной эволюции. В этом же периоде появились настоящие птицы и плацентарные млекопитающие. Признаки высокой степени организации у них — постоянная температура тела| полное разделение артериального и венозного тока крови, повышенный обмен веществ, совершенная терморегуляция, а у млекопитающих, кроме того, живорождение, вскармливание детенышей молоком, развитие коры головного мозга — позволили этим группам также занять господствующее положение на Земле.
5. Кайнозойская эра подразделяется на три периода: палеоген, неоген и четвертичный.
В палеогене, неогене и начале четвертичного периода цветковые растения благодаря приобретению многочисленных частных приспособлений заняли большую часть суши и представляли субтропическую и тропическую флору.
связи с похолоданием, вызванным наступлением ледника, субтропическая флора отступила к югу. В составе наземной растительности умеренных широт стали преобладать листопадные деревья, приспособленные к сезонному ритму температур, а также кустарники и травянистые растения. Расцвет травянистых приходится на четвертичный период. Большое распространение получили теплокровные животные:
птицы и млекопитающие. В ледниковое время обитали пещерные медведи, львы, мамонты, шерстистые носороги, которые после отступления ледников и потепления климата постепенно вымирали, а животный мир приобрел современный облик.
Главное событие этой эры — формирование человека. К концу неогена в лесах обитали небольшие хвостатые млекопитающие — лемуры и долгопяты.От них произошли древние формы обезьян — парапитеки, ведшие древесный образ жизни и питавшиеся растениями и насекомыми. Их далекие потомки — ныне живущие гиббоны, орангутанги и вымершие мелкие древесные обезьяны — дриопитеки. Дриопитеки дали начало трем линиям развития, которые привели к шимпанзе, горилле, а также вымершему австралопитеку. От австралопитеков в конце неогена произошел человек разумный.
Источник: www.examen.ru
Рубрики
Источник: kaz-ekzams.ru

В школьные годы изучение различных классов животных воспринималось как тяжкая повинность. То немногое, что было вызубрено в школе, благополучно забылось, и до недавнего времени я вряд ли мог бы причислить улитку или пиявку к тому или иному классу – как, вероятно, и большинство завсегдатаев Хабра и ГТ. Да и зачем вообще это помнить?
Но вот недавно попалась в руки книга Николая Александровича Бернштейна, написанная в 40-е годы XX века и изданная только в конце прошлого столетия. Название книги – «О ловкости и её развитии». Книга полна всевозможных открытий, и я рекомендую прочесть ее всем и каждому.
А в этой небольшой заметке попробую вкратце изложить взгляд автора книги на то, как эволюция животного мира стала результатом целой серии ключевых «технологических прорывов», нацеленных на развитие движений. Очень надеюсь, что этот реферат поможет привлечь внимание к научному наследию блестящего физиолога Н.А.Берштейна.
Итак, начнем все же с классификации животных. Бернштейн делит их на 7 основных классов:
- Простейшие – одноклеточные, микроскопически малые животные.
- Кишечнополостные (например, коралловые полипы, голотурии, губки, морские лилии).
- Иглокожие (например, морская звезда).
- Черви (например, дождевой червь, пиявка, ленточная глиста).
- Мягкотелые, или моллюски (например, улитка, каракатица, устрица).
- Членистоногие (насекомые, раки, пауки, сороконожки).
- Позвоночные (рыбы, лягушки, ящерицы, птицы, звери).
Считается, что простейшие (класс 1) были первыми представителями животного мира, и в далекие времена населяли теплый бульон мирового океана, в котором были растворены многочисленные химические элементы и соединения. Их организмы состояли всего из одной клетки, которая сама пробивала себе дорогу в жизнь, шевеля своими жгутиками или ложноножками и работая «за одну» в отношении и питания, и движения, и самосохранения, и размножения.
Первый апргрейд: дифференциация клеток и химический интерфейс
Следующим шагом в развитии животных стало появление многоклеточных организмов. Группа клеток уже способна «окружить» добычу и совместными усилиями создать химическую среду, которая расщепит пойманные в ловушку одиночные (и не только) клетки на «съедобные» элементы.
Именно так и действуют представители 2-го и 3-го классов животных. Кишечнополостные и иглокожие – округло-симметричные, малоподвижные существа.
Из класса кишечнополостных многие ведут полностью образ жизни растений, всю жизнь произрастая на одном месте. Их пищеварительная полость имеет еще вид мешка, и они пользуются как для питания, так и для испражнения одним и тем же отверстием. У иглокожих уже имеется сквозной пищеварительный канал.
Но с точки зрения эволюции гораздо более важной особенностью многоклеточных организмов является то, что их клетки уже потому не могут остаться равными друг другу, что одни из них находятся в глубинах тела, а другие – на поверхности. Возникает специализация клеток: лежащие на покровах тела приспосабливаются к «несению службы» раздражимости и чувствительности, другие, глубинные,– преимущественно к изменениям формы, к «сократительности», к обеспечению первобытных движений. Эти движения, пусть даже беспорядочные и ни на что не нацеленные, помогали многоклеточным просто потому, что шевелившиеся особи имели лучшие шансы в борьбе за жизнь, чем совершенно неподвижные.
Каждый физиологический процесс связан с какими-нибудь химическими превращениями в клетке. Рецептивные клетки поверхности тела, приобретшие повышенную раздражимость, тоже выделяли из себя во время своей деятельности какие-то химические продукты обмена веществ.
Понятно, что те особи, у которых, может быть чисто случайно, мышечные клетки оказались возбудимыми от действия проникавших в них рецептивных веществ, получили серьезное, почти решающее, биологическое преимущество перед другими. В то время, как эти последние были способны только на самопроизвольные шевеления, иногда бывшие просто ни к чему, а иногда бывшие и прямо невпопад, особи новой «марки» могли реагировать на внешние раздражения (например, поворачиваться лицом к добыче или спиной к опасности).
Это новое явление на Земле – реактивность – поначалу было огульным, неизбирательным, расплывчатым, как говорят в физиологии, диффузным. Мы и сейчас можем наблюдать у различных низших организмов подобную диффузную раздражимость и реактивность: пока не трогаешь его, он лежит смирно; прикоснешься – начинаются общие неупорядоченные движения тела, тем более значительные, чем сильнее было раздражение.
Так выявились первые в природе химические возбуждающие мышцу вещества – первобытные посредники между рецептивной поверхностью тела и мышцами. Эти вещества называются в физиологии посредниками – медиаторами, и они и по сию пору у самых высших организмов (в т.ч. у Homo Sapiens) играют очень существенную для движений роль. Каждый раз, как мы при ходьбе или выполнении гимнастических упражнений произвольно напрягаем ту или иную мышцу, у ее нервных окончаний выделяется микроскопически малая капелька вещества, которому 500 миллионов лет.
В последующих поколениях организмов начали мало-помалу обособляться каналы, специально приспособленные для доставки химических медиаторов. Однако не успели еще эти «водные пути сообщения» как следует оформиться и обеспечить хоть какую-то избирательную адресацию медиаторов к тем или иным мышечным группам, произошло другое событие, биологическое значение которого оказалось неизмеримо большим.
Развитие электрического интерфейса
Каждое химическое явление имеет свой электрический «отблеск», сопровождается теми или иными колебаниями электрического потенциала.
Так уж получилось (в порядке случайных прирожденных изменений, или еще как), что у некоторых экземпляров их мышечные клетки оказались возбудимыми не только от прямого химического воздействия медиатора, но уже и от одного только электрического спутника последнего – от того неуловимо малого электрического колебания, которым он всегда сопровождался.
Преимущества электрического способа передачи сигналов перед химическим очевидны. Во-первых, волна электрического импульса имеет гораздо большую скорость, нежели раствор, медленно сочащийся по межтканевым щелям,– значит, она дает возможность ее обладателю реагировать во много раз быстрее. Во-вторых, электрический возбуждающий импульс несет в себе хоть какие-то возможности для его «маршрутизации» в ту или другую мышечную группу, в то время как жидкость, содержащая медиатор, обязательно омывает весь организм. Неудивительно, что вновь открытый природой «телеграфный» принцип передачи возбудительных импульсов начал энергично завоевывать себе командное положение.
Постепенно вычленились волокна, обнаруживавшие лучшую проводимость для биотоков. Такие волокна представляли собою длинные отростки клеток. Специализировавшиеся на передаче импульсов (пора уже начать называть их нервными импульсами) волоконца образовали внутри организма сети, позволяющие передавать сигналы к нужным частям тела.
Но на данном этапе нервные сети отвечали только за передачу данных, как телеграфные столбы.
Отделение рта от ануса
Маловажным на вид обстоятельством оказалось появление на Земле продолговатых, колбасовидных животных форм. Те классы животных (2-й и 3-й по нашей классификации), которые были описываемы до сих пор, имели округло-симметричные формы, с ротовым отверстием посередине.
Очертания тела низших из них, кишечнополостных, менее определенны; это по сути дела, мешки с одним отверстием, что понуждает их заменять естественные отправления тела рвотой. Более продвинутые в своем развитии (сквозной пищеварительный канал) иглокожие имеют лучистое строение, и вокруг центрального рта обладают пятью симметричными отростками (лучами у морских звезд, лимонообразными дольками и у морского ежа и т.д.).
На смену им начинают появляться продолговатые животные классов 4 и 5 (в последующем – черви и моллюски) с пищеварительной трубкой, тянущейся во всю длину их тела, с ротовым отверстием на одном и заднепроходным – на другом конце. В ротовом конце-то и было все дело.
Ясно, что ротовой конец тела – это его активный конец. Он ищет питания, он первым сталкивается с добычей, первым же – и с опасностью. Он, как правило, движется впереди.
Переднему концу тела важнее, чем какой-либо другой его точке, тонко и своевременно ощутить свойства того, с чем он соприкоснулся, к чему он подполз. Но кроме обострения древних видов чувствительности (осязательная, температурная, вкусовая, химическая), которые можно объединить под общим названием «контактной» или «чувствительности непосредственного соприкосновения», на переднем, ротовом, конце начинают развиваться качественно новые, более совершенные виды органов чувств.
Новым рецепторам удобно присвоить, воспользовавшись широко привившейся в техническом языке приставкой, имя телерецепторов. По аналогии этого слова с такими терминами, как телефон, телеграф, телевидение и т. п., легко понять его смысл: речь идет о дальнодействующих или дальнобойных рецепторах.
Каждый из древних видов контактных рецепторов, видоизменяясь, породил один из усовершенствованных дальнодействующих. Орган химической чувствительности – вкус – дал начало химическому телерецептору – органу обоняния. Осязательная чувствительность переднего конца, утончаясь, обратилась в чувствительность к частым и мелким сотрясениям, или вибрациям, передаваемым издали через окружающую среду: в орган слуха, «слышания» звуков, которые и есть не что иное, как колебания, или вибрации, воды или воздуха.
Наконец, температурная контактная чувствительность преобразовалась сперва в восприимчивость к лучистой теплоте, а затем и к лучистой энергии самого мощного отдела солнечного спектра – световой энергии. Отсюда, таким образом, возникло зрение.
С появлением телерецепторов (воспринимаемый) окружающий мир стал гораздо больше по объему, чем само животное. Теперь можно было узреть-унюхать-услышать добычу или опасность издалека, и, соответственно, двигаться к ней либо от нее.
Обладание телерецепторами породило целый ряд новых потребностей для организма, таких как:
- согласование движений различных частей тела, чтобы можно было двигаться в заданном направлении как единое целое;
- механизм планирования действий или движений, чтобы можно было выработать тактику нападения на, или самообороны от, объекта, обнаруженного на расстоянии (загодя);
- зачатки памяти, хотя бы для того, чтобы запомнить план действий, о котором говорилось выше.
Для удовлетворения указанных выше потребностей у животных сформировались скопления нервных клеток, называемые нервными узлами, или ганглиями, которые взяли на себя функции согласования, планирования и запоминания.
Естественно, что этим центрам всего более подходит помещаться на переднем конце, так сказать, на капитанском мостике всего тела, там, где находятся все телерецепторы, и там, откуда наиболее открытый вид для наблюдения.
Таким путем ротовой конец оказался сперва, по неминуемой логике вещей, передним концом тела, а затем, оснастившись в качестве переднего высокопробными телерецепторами, стал головным концом тела и, наконец, его главным концом.
Как ни парадоксально, именно отделение рта от ануса привело к возникновению глаз, ушей, носа и мозга.
Забегая вперед, отметим, что физиологические особенности рептилий чуть было не привели к возникновению у них второго мозга, расположенного именно в области ануса. Но об этом – читайте далее.
Как анизотропные диски изменили мир
До возникновения анизотропных дисков древний животный мир был медленным и неторопливым. Мышцы животных в те времена были способны только на слабые и медленные движения. Но вот появился новый элемент, который сделал возможным формирование совершенно нового типа мускулатуры.
Анизотропные диски – это особые клетки, которые способны к молниеносным сокращениям под действием электрического тока. Именно благодаря этому крылья комара или мухи способны совершать несколько сот движений в секунду.
Другое свойство анизотропных дисков – это высокая механическая мощность, вырабатываемая при их сокращении. При том же весе, анизотропные диски вырабатывают в тысячи раз большую мощность, чем клетки более древних гладких мышц.
Анизотропные диски настолько успешно решали проблемы быстроты и мощности, что тела организмов, вопреки обычному порядку вещей, стали всячески подстраиваться под использование анизотропных дисков (а не наоборот, т.е. не стали дожидаться, пока анизотропные диски подстроятся под особенности организма).
Действительно, анизотропные диски были настолько мощными и быстрыми, что древние мягкотелые организмы могло просто разорвать при их использовании. Поэтому в первую очередь были предприняты меры к тому, чтобы сгладить взрывной эффект анизотропных дисков.
Во-первых, взрывной эффект от действия анизотропных дисков компенсируется тем, что в мышцах животных и человека анизотропные диски чередуются с упругими клетками сухожильной ткани (изо-элементами), которые играют роль амортизаторов. Длинная цепочка чередующихся анизотропных и изотропных клеток образует мышечные волокна, которые именно благодаря такому чередованию клеток носят название поперечно-полосатой мышцы.
Во-вторых, каждое волоконце двигательного нерва врощено своими разветвлениями в пачку из 10 – 100 мышечных волокон, которые, очевидно, под действием его импульсов могут двигаться не иначе, как все разом и все одинаково. Такая пачка мышечных волокон носит название миона. Каждая мышца нашего тела состоит в зависимости от своей величины из нескольких десятков или сотен мионов.
В-третьих, каждый анизотропный диск сокращается в течение тысячной доли секунды, а потом ему нужно «отдохнуть» в течение времени, которое длится в 3-4 раза дольше, чем сокращение. Поэтому нервная система посылает к мышцам целые серии импульсов возбуждения с частотой 50-200 герц, чтобы можно было продолжить начатое ранее действие мускулатуры.
В-четвертых, все мышечные клетки помещены в густую и вязкую жидкость, называемую саркоплазмой. Эта жидкость в свою очередь обеспечивает плавность мускульных движений. Заметим, что исключительная быстрота движений некоторых представителей фауны (например, тех же мух) связана с ничтожно малым количеством саркоплазмы в их мускулатуре.
Членистоногие против позвоночных
Итак, есть мощный и быстрый источник движущей силы – поперечно-полосатая мышца.
Как пишет Бернштейн, дело пошло так, как будто бы на великом конкурсе, объявленном жизнью на наилучшее оснащение для поперечнополосатой мышцы, первую премию поделили между собой два разных проекта. Оба они по первоначальному рассмотрению как будто одинаково хорошо и остроумно решали поставленную конкурсом задачу, хотя решали ее глубоко различными между собой способами.
Один из проектов шел под девизом Arthropoda (членистоногие), другой – под девизом Vertebrata (позвоночные). Оба проекта исходили из поперечнополосатой мышцы как чего-то «уже данного», и оба объединяли ее с жесткими, суставчато-подвижными скелетами; то и другое входило, очевидно, в «технические условия» конкурса.
Членистоногие создали жесткие внешние скелеты наподобие рыцарских доспехов, и поместили в них поперечно-полосатые мышцы. Все твердые элементы брони – членики – соединены между собой с той или иной степенью подвижности. Отсюда и название класса: членистоногие.
Позвоночные пошли прямо противоположным путем. Они поместили твердый скелет внутри организма, «обмотав» его мускулами со всех сторон. Центральным элементом скелета является состоящий из отдельных сегментов позвоночник. Его структура сохраняется благодаря «обмотке» из мышц, тянущих каждый позвонок одновременно в разные стороны.
Преимущество проекта членистоногих состояло в том, что строение их тела одновременно решало 2 дополнительные (помимо двигательных) задачи: обеспечивало организм броней, и в то же время обеспечивало устойчивость. В самом деле, даже мертвые насекомые сохраняют устойчивое положение тела, потому что для этого не требуется напряжения мышц.
Конструкция позвоночных, очевидно, не обладала указанными преимуществами, поскольку не предусматривала изначально жесткой брони и требовала постоянного напряжения мускулатуры для поддержания устойчивости. Ведь, как известно, любое позвоночное животное, будучи убитым, обязательно падает. Зато эти недостатки с лихвой компенсировались дополнительными степенями свободы в движениях и гибкостью.
По мнению Берштейна, именно ограниченность движений членистоногих завела их в «эволюционный тупик», поскольку у них не было потребности в развитии «управленческого аппарата» для сложных движений. Хотя, конечно же, не могут не удивлять сложные и великолепно организованные формы существования некоторых насекомых, таких как пчелы или термиты.
В то же время, новые степени свободы в движениях, появившиеся у позвоночных, создали предпосылки для развития у них высшей нервной системы.
Как бы то ни было, оба проекта были успешно имплементированы, и в настоящее время представляют доминирующие формы жизни на нашей планете.
Куда делись динозавры?
Заключительная часть нашего реферата посвящена любопытной точке зрения Берштейна на причину исчезновения динозавров.
Как известно, первыми позвоночными были древние рыбы. Их представителей до сих пор можно встретить в океанах – акулы, скаты и т.д. От более поздних рыб их отличает отсутствие костных скелетов, присущих ершам, щукам и окуням.
Амфибии и земноводные (лягушки, тритоны) были первыми среди тех позвоночных, которые выбрались на сушу. Ясное дело, для них потребовался еще один «апгрейд»: развитие конечностей, без которых по суше не очень то и поползаешь.
От амфибий и земноводных впоследствии произошли рептилии, которые царствовали на суше чрезвычайно долго, достигнув расцвета в Юрский период. Когда-то пресмыкающиеся существовали на земном шаре в огромном количестве отрядов и видов, владея и поверхностью воды, и сушей, и воздухом.
В наше время от всего этого обилия гадов уцелели только остатки, всего четыре отряда: ящерицы, черепахи, змеи и крокодилы.
Как известно, динозавры так и не смогли развить головной мозг. Причиной этого, по-видимому, явились особенности их физиологии и размеры.
Известно и точно измерено, что скорость, с которой распространяется вдоль нерва электрохимический сигнал возбуждения – нервный импульс, очень различна у холоднокровных и у теплокровных животных. Волна возбуждения бежит по нерву лягушки со скоростью 8 – 10 метров в секунду, а по нерву кошки или человека – со скоростью 100-120 метров в секунду.
А теперь сделаем несложный расчет.
Представим себе, что кто-то укусил тридцатиметрового великана-ящера за заднюю лапу, и он, почувствовав боль, отдернул лапу или ударил ею обидчика. Для пути ощущения боли имеем: лапа 6 метров, туловище 10 метров, шея еще 10 метров, итого 26 метров, т. е. три секунды в один конец. Положим столько же для ответного двигательного приказа от головного мозга к ножным мышцам; к этому нужно прибавить еще хоть секунду на скорость реакции в самом мозгу. В итоге получается, что от момента укуса до начала ответного движения пройдет семь секунд – срок очень немалый, если посмотреть на секундную стрелку своих ручных часов и «вытерпеть» семь секунд, внимательно следя за ней.
Измерение сохранившихся скелетов юрских и меловых гигантских ящеров показывает, что у них на длинной мощной шее сидела крохотная голова, так же подходившая им по пропорциям, как нам подошла бы мышиная. И в этой-то непомерно малой голове большая ее часть была занята лицевым скелетом – зубастою пастью, на долю же головного мозга оставалось совсем тесное, ничтожное вместилище. Мы легче поймем это, если учтем, что животное, которое было бы вынуждено «спрашиваться» обо всех своих движениях у головного мозга и дожидаться ответов по семи секунд, было бы нежизнеспособным.
Очевидно, преобладающая часть их двигательных реакций протекала под управлением одного только спинного мозга. Это давало очень значительное укорочение нервного маршрута: секунд до двух-трех. И действительно, у очень многих из этих ящеров в позвоночном канале – вместилище спинного мозга – имеется вздутие в области поясницы и крестца, там, где начинаются нервы задних лап. Это вздутие говорит о том, что в этом месте спинной мозг был расширен, и очень значительно: он был здесь даже больше головного.
Так вот, по мнению Н.А.Берштейна, именно скорость передачи данных сыграла решающую роль в определении исхода схватки между вновь появившимся классом млекопитающих и рептилиями.
Новый, молодой класс млекопитающих теплокровных, темпераментных, с очень обогащенными как мозгом, так в связи с этим и двигательными средствами, – оказался для рептилий неодолимым противником. Некрупные, ловкие хищники накинулись на эти медлительные, как будто нарочно для них заготовленные горы мяса, и своим действительно хищническим хозяйствованием быстро вывели их из употребления.
Другими словами, динозавры исчезли не в результате падения метеорита или наступления ледников. Их просто сожрали теплокровные!
Источник: habr.com