Стадии митоза таблица
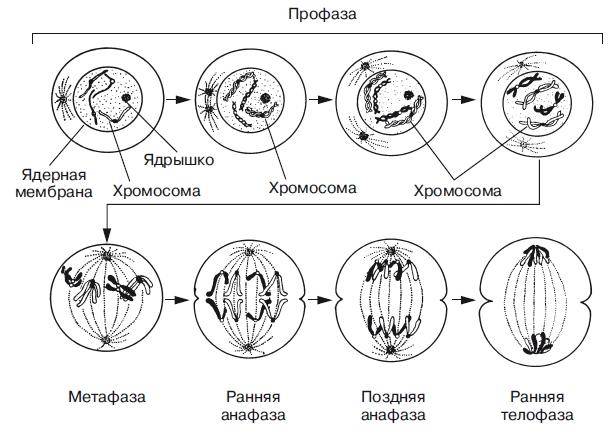
Митоз условно разделяют на четыре фазы: профазу, метафазу, анафазу и телофазу. Профаза. Две центриоли начинают расходиться к противоположным полюсам ядра. Ядерная мембрана разрушается; одновременно специальные белки объединяются, формируя микротрубочки в виде нитей. Центриоли, расположенные теперь на противоположных полюсах клетки, оказывают организующее воздействие на микротрубочки, которые в результате выстраиваются радиально, образуя структуру, напоминающую по внешнему виду цветок астры («звезда»). Другие нити из микротрубочек протягиваются от одной центриоли к другой, образуя веретено деления. В это время хромосомы спирализуются и вследствие этого утолщаются. Они хорошо видны в световом микроскопе, особенно после окрашивания. Считывание генетической информации с молекул ДНК становится невозможным: синтез РНК прекращается, ядрышко исчезает. В профазе хромосомы расщепляются, но хроматиды все еще остаются скрепленными попарно в зоне центромеры. Центромеры тоже оказывают организующее воздействие на нити веретена, которые теперь тянутся от центриоли к центромере и от нее к другой центриоли. Метафаза. В метафазе спирализация хромосом достигает максимума, и укороченные хромосомы устремляются к экватору клетки, располагаясь на равном расстоянии от полюсов. Образуется экваториальная, или метафазная, пластинка. На этой стадии митоза отчетливо видна структура хромосом, их легко сосчитать и изучить их индивидуальные особенности. В каждой хромосоме имеется область первичной перетяжки — центромера, к которой во время митоза присоединяются нить веретена деления и плечи. На стадии метафазы хромосома состоит из двух хроматид, соединенных между собой только в области центромеры.
Рис. 1. Митоз растительной клетки. А — интерфаза; В анафазе вязкость цитоплазмы уменьшается, центромеры разъединяются, и с этого момента хроматиды становятся самостоятельными хромосомами. Нити веретена деления, прикрепленные к центромерам, тянут хромосомы к полюсам клетки, а плечи хромосом при этом пассивно следуют за центромерой. Таким образом, в анафазе хроматиды удвоенных еще в интерфазе хромосом точно расходятся к полюсам клетки. В этот момент в клетке находятся два диплоидных набора хромосом (4n4с).
Таблица 1. Митотический цикл и митоз
В телофазе хромосомы раскручиваются, деспирализуются. Из мембранных структур цитоплазмы образуется ядерная оболочка. В это время восстанавливается ядрышко. На этом завершается деление ядра (кариокинез), затем происходит деление тела клетки (или цитокинез). При делении животных клеток на их поверхности в плоскости экватора появляется борозда, постепенно углубляющаяся и разделяющая клетку на две половины — дочерние клетки, в каждой их которых имеется по ядру. У растений деление происходит путем образования так называемой клеточной пластинки, разделяющей цитоплазму: она возникает в экваториальной области веретена, а затем растет во все стороны, достигая клеточной стенки (т.е. растет изнутри кнаружи). Клеточная пластинка формируется из материала, поставляемого эндоплазматической сетью. Затем каждая из дочерних клеток образует на своей стороне клеточную мембрану и, наконец, на обеих сторонах пластинки образуются целлюлозные клеточные стенки. Особенности протекания митоза у животных и растений приведены в таблице 2. Таблица 2. Особенности митоза у растений и у животных
Так из одной клетки формируются две дочерние, в которых наследственная информация точно копирует информацию, содержавшуюся в материнской клетке. Начиная с первого митотического деления оплодотворенной яйцеклетки (зиготы) все дочерние клетки, образовавшиеся в результате митоза, содержат одинаковый набор хромосом и одни и те же гены. Следовательно, митоз — это способ деления клеток, заключающийся в точном распределении генетического материала между дочерними клетками. В результате митоза обе дочерние клетки получают диплоидный набор хромосом. Весь процесс митоза занимает в большинстве случаев от 1 до 2 часов. Частота митоза в разных тканях и у разных видов различна. Например, в красном костном мозге человека, где каждую секунду образуется 10 млн эритроцитов, в каждую секунду должно происходить 10 млн. митозов. А в нервной ткани митозы крайне редки: так, в центральной нервной системе клетки в основном перестают делиться уже в первые месяцы после рождения; а в красном костном мозге, в эпителиальной выстилке пищеварительного тракта и в эпителии почечных канальцев они делятся до конца жизни. Регуляция митоза, вопрос о пусковом механизме митоза. Факторы, побуждающие клетку к митозу точно не известны. Но полагают, что большую роль играет фактор соотношения объемов ядра и цитоплазмы (ядерно-плазменное соотношение). По некоторым данным, отмирающие клетки продуцируют вещества, способные стимулировать деление клетки. Белковые факторы, отвечающие за переход в фазу М, первоначально были идентифицированы на основе экспериментов по слиянию клеток. Слияние клетки, находящейся в любой стадии клеточного цикла, с клеткой находящейся в М фазе, приводит к вхождению ядра первой клетки в М фазу. Это означает, что в клетке находящейся в М фазе существует цитоплазматический фактор способный активировать М фазу. Позднее этот фактор был вторично обнаружен в экспериментах по переносу цитоплазмы между ооцитами лягушки, находящимися на различных стадиях развития, и был назван "фактором созревания" MPF (maturation promoting factor). Дальнейшее изучение MPF показало, что этот белковый комплекс детерминирует все события М-фазы. На рисунке показано, что распад ядерной мембраны, конденсация хромосом, сборка веретена, цитокинез регулируются MPF. Митоз тормозится высокой температурой, высокими дозами ионизирующей радиации, действием растительных ядов. Один из таких ядов называется колхицин. С его помощью можно остановить митоз на стадии метафазной пластинки, что позволяет подсчитать число хромосом и дать каждой из них индивидуальную характеристику, т. е. провести кариотипирование. Амитоз (от греч. а – отриц. частица и митоз)-прямоеделение интерфазного ядра путем перешнуровывания без преобразования хромосом. При амитозе не происходит равномерное расхождение хроматид к полюсам. И это деление не обеспечивает образование генетически равноценных ядер и клеток. По сравнению с митозом амитоз более кратковременный и экономичный процесс. Амитотическое деление может осуществляться несколькими способами. Наиболее распространенный тип амитоза – это перешнуровывание ядра на две части. Этот процесс начинается с разделения ядрышка. Перетяжка углубляется, и ядро разделяется надвое. После этого начинается разделение цитоплазмы, однако это происходит не всегда. Если амитоз ограничивается только делением ядра, то это приводит к образованию дву- и многоядерных клеток. При амитозе может также происходить почкование и фрагментация ядер. Клетка, претерпевшая амитоз, в последующем не способна вступить в нормальный митотический цикл. Амитоз встречается в клетках различных тканей растений и животных. У растений амитотическое деление довольно часто встречается в эндосперме, в специализирующихся клетках корешков и в клетках запасающих тканей. Амитоз также наблюдается в высокоспециализированных клетках с ослабленной жизнеспособностью или дегенерирующих, при различных патологических процессах, таких как злокачественный рост, воспаление и т. п.
Источник: studopedia.info Митоз — это деление клетки, при котором дочерние клетки генетически идентичны материнской и между собой. То есть при митозе хромосомы удваиваются и распределяются между дочерними клетками так, чтобы каждая получила по одной хроматиде каждой хромосомы. В митозе выделяют несколько стадий (фаз). Однако самому митозу предшествует длительная интерфаза. Митоз и интерфаза вместе составляют клеточный цикл. В процессе интерфазы клетка растет, в ней образуются органоиды, активно идут процессы синтеза. В синтетическом периоде интерфазы редуплицируется, т. е. удваивается, ДНК. После удвоения хроматид они остаются соединенными в области центромеры, т. е. хромосома состоит из двух хроматид. В самом митозе обычно выделяют четыре основные стадии (иногда больше).
Первая стадия митоза — профаза. В эту фазу хромосомы спирализуются и приобретают компактную скрученную форму. Из-за этого становятся невозможны процессы синтеза РНК. Исчезают ядрышки, а значит, рибосомы также не образуются, т. е. синтетические процессы в клетке приостанавливаются. Центриоли расходятся к полюсам (в разные концы) клетки, начинает образовываться веретено деления. В конце профазы распадается ядерная оболочка. Прометафаза — это стадия, которую не всегда выделяют отдельно. Процессы, происходящие в ней, могут относить к поздней профазе или ранней метафазе. В прометафазе хромосомы оказываются в цитоплазме, беспорядочно перемещаются по клетке пока в районе центромеры не соединятся с нитью веретена деления. Нить представляет собой микротрубочку, построенную из белка тубулина. Она нарастает присоединяя новые тубулиновые субъединицы. При этом хромосома движется от полюса. Со стороны другого полюса к ней также присоединяется нить веретена и также толкает ее от полюса. Вторая стадия митоза — метафаза. Все хромосомы располагаются в экваториальной области клетки рядом. К их центромерам прикреплено по две нити веретена деления. В митозе метафаза самая длительная стадия. Третья стадия митоза — анафаза. В этой фазе хроматиды каждой хромосомы отделяются друг от друга и за счет тянущих их нитей веретена деления отходят к разным полюсам. Микротрубочки теперь не нарастают, а разбираются. Анафаза достаточно быстрая фаза митоза. При расхождении хромосом органоиды клетки примерно в равных количествах также расходятся ближе к полюсам. Четвертая стадия митоза — телофаза — во многом обратна профазе. Хроматиды собираются у полюсов клетки и раскручиваются, т. е. деспирализуются. Вокруг них формируются ядерные оболочки. Образуются ядрышки, начинается синтез РНК. Веретено деления начинает разрушаться. Далее происходит деление цитоплазмы — цитокинез. В клетках животных это происходит за счет впячивания мембраны внутрь и образования перетяжки. В клетках растений мембрана начинает формироваться внутри в экваториальной плоскости и идет к периферии.
Источник: scienceland.info Митоз, его фазы, биологическое значение Важнейшим компонентом клеточного цикла является митотический (пролиферативный) цикл. Он представляет собой комплекс взаимосвязанных и согласованных явлений во время деления клетки, а также до и после него. Митотический цикл — это совокупность процессов, происходящих в клетке от одного деления до следующего и заканчивающихся образованием двух клеток следующей генерации. Кроме этого, в понятие жизненного цикла входят также период выполнения клеткой своих функций и периоды покоя. В это время дальнейшая клеточная судьба неопределенна: клетка может начать делиться (вступает в митоз) либо начать готовиться к выполнению специфических функций. Основные стадии митоза. 1.Редупликация (самоудвоение) генетической информации материнской клетки и равномерное распределение ее между дочерними клетками. Это сопровождается изменениями структуры и морфологии хромосом, в которых сосредоточено более 90% информации эукариотической клетки. 2.Митотический цикл состоит из четырех последовательных периодов: пресинтетического (или постмитотического) G1, синтетического S, постсинтетического (или премитотического) G2 и собственно митоза. Они составляют автокаталитическую интерфазу (подготовительный период). Фазы клеточного цикла: 1) пресинтетическая (G1). Идет сразу после деления клетки. Синтеза ДНК еще не происходит. Клетка активно растет в размерах, запасает вещества, необходимые для деления: белки (гистоны, структурные белки, ферменты), РНК, молекулы АТФ. Происходит деление митохондрий и хлоропластов (т. е. структур, способных к ауторепродукции). Восстанавливаются черты организации интерфазной клетки после предшествующего деления; 2) синтетическая (S). Происходит удвоение генетического материала путем репликации ДНК. Она происходит полуконсервативным способом, когда двойная спираль молекулы ДНК расходится на две цепи и на каждой из них синтезируется комплементарная цепочка. В итоге образуются две идентичные двойные спирали ДНК, каждая из которых состоит из одной новой и старой цепи ДНК. Количество наследственного материала удваивается. Кроме этого, продолжается синтез РНК и белков. Также репликации подвергается небольшая часть митохонд-риальной ДНК (основная же ее часть реплицируется в G2 период); 3) постсинтетическая (G2). ДНК уже не синтезируется, но происходит исправление недочетов, допущенных при синтезе ее в S период (репарация). Также накапливаются энергия и питательные вещества, продолжается синтез РНК и белков (преимущественно ядерных). S и G2 непосредственно связаны с митозом, поэтому их иногда выделяют в отдельный период — препрофазу. После этого наступает собственно митоз, который состоит из четырех фаз. Процесс деления включает в себя несколько последовательных фаз и представляет собой цикл. Его продолжительность различна и составляет у большинства клеток от 10 до 50 ч. При этом у клеток тела человека продолжительность самого митоза составляет 1—1,5 ч, G2-периода интерфазы — 2—3 ч, S-периода интерфазы — 6—10 ч. Стадии митоза. Процесс митоза принято подразделять на четыре основные фазы: профазу, метафазу, анафазу и телофазу (рис. 1–3). Так как он непрерывен, смена фаз осуществляется плавно — одна незаметно переходит в другую. В профазе увеличивается объем ядра, и вследствие спирализации хроматина формируются хромосомы. К концу профазы видно, что каждая хромосома состоит из двух хроматид. Постепенно растворяются ядрышки и ядерная оболочка, и хромосомы оказываются беспорядочно расположенными в цитоплазме клетки. Центриоли расходятся к полюсам клетки. Формируется ахроматиновое веретено деления, часть нитей которого идет от полюса к полюсу, а часть — прикрепляется к центромерам хромосом. Содержание генетического материала в клетке остается неизменным (2n2хр).
Профаза К основным событиям профазы относят конденсацию хромосом внутри ядра и образование веретена деления в цитоплазме клетки. Распад ядрышка в профазе является характерной, но не обязательной для всех клеток особенностью. Условно за начало профазы принимается момент возникновения микроскопически видимых хромосом вследствие конденсации внутриядерного хроматина. Уплотнение хромосом происходит за счёт многоуровневой спирализации ДНК. Данные изменения сопровождаются повышением активности фосфорилаз, модифицирующих гистоны, непосредственно участвующие в компоновке ДНК. Как следствие, резко снижается транскрипционная активность хроматина, инактивируются ядрышковые гены, большая часть ядрышковых белков диссоциирует. Конденсирующиеся сестринские хроматиды в ранней профазе остаются спаренными по всей своей длине с помощью белков-когезинов, однако к началу прометафазы связь между хроматидами сохраняется лишь в области центромер. К поздней профазе на каждой центромере сестринских хроматид формируются зрелые кинетохоры необходимые хромосомам для присоединения к микротрубочкам веретена деления в прометафазе. Наряду с процессами внутриядерной конденсации хромосом в цитоплазме начинает формироваться митотическое веретено — одна из главных структур аппарата клеточного деления, ответственная за распределение хромосом между дочерними клетками. В образовании веретена деления у всех эукариотических клеток принимают участие полярные тельца, микротрубочки и кинетохоры хромосом. С началом формирования митотического веретена в профазе сопряжены разительные изменения динамических свойств микротрубочек. Время полужизни средней микротрубочки уменьшается примерно в 20 раз от 5 минут до 15 секунд. Однако скорость их роста увеличивается примерно в 2 раза по сравнению с теми же интерфазными микротрубочками. Полимеризующиеся плюс-концы являются «динамически нестабильными» и резко переходят от равномерного роста к быстрому укорочению, при котором часто деполимеризуется вся микротрубочка. Примечательно, что для правильного функционирования митотического веретена необходим определенный баланс между процессами сборки и деполимеризации микротрубочек, так как ни стабилизированные, ни деполимеризованные микротрубочки веретена не в состоянии перемещать хромосомы. Наряду с наблюдаемыми изменениями динамических свойств микротрубочек, слагающих нити веретена, в профазе закладываются полюса деления. Реплицированные в S-фазе центросомы расходятся в противоположных направлениях за счёт взаимодействия полюсных микротрубочек, растущих навстречу друг другу. Своими минус-концами микротрубочки погружены в аморфное вещество центросом, а процессы полимеризации протекают со стороны плюс-концов, обращенных к экваториальной плоскости клетки. При этом вероятный механизм расхождения полюсов объясняется следующим образом: динеино-подобные белки ориентируют в параллельном направлении полимеризующиеся плюс-концы полюсных микротрубочек, а кинезино-подобные белки в свою очередь расталкивают их в направлении к полюсам деления. Параллельно конденсации хромосом и формированию митотического веретена, во время профазы происходит фрагментация эндоплазматического ретикулума, который распадается на мелкие вакуоли, расходящиеся затем к периферии клетки. Одновременно рибосомы теряют связи с мембранами ЭПР. Цистерны аппарата Гольджи также меняют свою околоядерную локализацию, распадаясь на отдельные диктиосомы, без особого порядка распределенные в цитоплазме. Прометафаза Прометафаза Окончание профазы и наступление прометафазы, как правило, знаменуется распадом ядерной мембраны. Целый ряд белков ламины фосфорилируется, вследствие чего ядерная оболочка фрагментируется на мелкие вакуоли, а поровые комплексы исчезают. После разрушения ядерной мембраны хромосомы без особого порядка располагаются в области ядра. Однако вскоре все они приходят в движение. В прометафазе наблюдается интенсивное, но беспорядочное перемещение хромосом. Поначалу отдельные хромосомы стремительно дрейфуют к ближайшему полюсу митотического веретена со скоростью, достигающей 25 мкм/мин. Вблизи полюсов деления повышается вероятность взаимодействия новосинтезированных плюс-концов микротрубочек веретена с кинетохорами хромосом. В результате такого взаимодействия кинетохорные микротрубочки стабилизируются от спонтанной деполимеризации, а их рост отчасти обеспечивает отдаление соединенной с ними хромосомы в направлении от полюса к экваториальной плоскости веретена. С другой стороны хромосому настигают тяжи микротрубочек, идущие от противоположного полюса митотического веретена. Взаимодействуя с кинетохором, они также участвуют в движении хромосомы. В результате сестринские хроматиды оказываются связанными с противоположными полюсами веретена. Усилие, развиваемое микротрубочками от разных полюсов, не только стабилизирует взаимодействие этих микротрубочек с кинетохорами, но также, в конечном счёте, приводит каждую хромосому в плоскость метафазной пластинки. В клетках млекопитающих прометафаза протекает, как правило, в течение 10—20 минут. В нейробластах кузнечика данная стадия занимает всего 4 минуты, а в эндосперме Haemanthus и в фибробластах тритона — около 30 минут. Метафаза Метафаза В завершении прометафазы хромосомы располагаются в экваториальной плоскости веретена примерно на равном расстоянии от обоих полюсов деления, образуя метафазную пластинку. Морфология метафазной пластинки в клетках животных, как правило, отличается упорядоченным расположением хромосом: центромерные участки обращены к центру веретена, а плечи — к периферии клетки. В растительных клетках хромосомы зачастую лежат в экваториальной плоскости веретена без строгого порядка. Метафаза занимает значительную часть периода митоза, и отличается относительно стабильным состоянием. Все это время хромосомы удерживаются в экваториальной плоскости веретена за счёт сбалансированных сил натяжения кинетохорных микротрубочек, совершая колебательные движения с незначительной амплитудой в плоскости метафазной пластинки. В метафазе, также как и в течение других фаз митоза, продолжается активное обновление микротрубочек веретена путём интенсивной сборки и деполимеризации молекул тубулина. Несмотря на некоторую стабилизацию пучков кинетохорных микротрубочек, происходит постоянная переборка межполюсных микротрубочек, численность которых в метафазе достигает максимума. К окончанию метафазы наблюдается чёткое обособление сестринских хроматид, соединение между которыми сохраняется лишь в центромерных участках. Плечи хроматид располагаются параллельно друг другу, и становится отчетливо заметной разделяющая их щель. Анафаза Анафаза Анафаза — самая короткая стадия митоза, которая начинается внезапным разделением и последующим расхождением сестринских хроматид в направлении противоположных полюсов клетки. Хроматиды расходятся с равномерной скоростью достигающей 0,5—2 мкм/мин., при этом они часто принимают V-образную форму. Их движение обусловлено воздействием значительных сил, оценочно 10 дин на хромосому, что в 10 000 раз превышает усилие, необходимое для простого продвижения хромосомы через цитоплазму с наблюдаемой скоростью. Как правило, расхождение хромосом в анафазе состоит из двух относительно независимых процессов называемых анафазой А и анафазой В. Анафаза А характеризуется расхождением сестринских хроматид к противоположным полюсам деления клетки. За их движение при этом отвечают те же силы, что ранее удерживали хромосомы в плоскости метафазной пластинки. Процесс расхождения хроматид сопровождается сокращением длины деполимеризующихся кинетохорных микротрубочек. Причем их распад наблюдается преимущественно в области кинетохоров, со стороны плюс-концов. Вероятно, деполимеризация микротрубочек у кинетохоров либо в области полюсов деления является необходимым условием для перемещения сестринских хроматид, так как их движение прекращается при добавлении таксола или тяжёлой воды, оказывающих стабилизирующее воздействие на микротрубочки. Механизм, лежащий в основе расхождения хромосом в анафазе А, пока остается неизвестным. Во время анафазы В расходятся сами полюса деления клетки, и, в отличии от анафазы А, данный процесс происходит за счёт сборки полюсных микротрубочек со стороны плюс-концов. Полимеризующиеся антипараллельные нити веретена при взаимодействии отчасти и создают расталкивающее полюса усилие. Величина относительного перемещения полюсов при этом, также как и степень перекрывания полюсных микротрубочек в экваториальной зоне клетки сильно варьирует у особей разных видов. Помимо расталкивающих сил, на полюса деления воздействуют тянущие силы со стороны астральных микротрубочек, которые создаются в результате взаимодействия с динеино-подобными белками на плазматической мембране клетки. Последовательность, продолжительность и относительный вклад каждого из двух процессов, слагающих анафазу, могут быть крайне различны. Так в клетках млекопитающих анафаза В начинается сразу вслед за началом расхождения хроматид к противоположным полюсам и продолжается вплоть до удлинения митотического веретена в 1,5—2 раза по сравнению с метафазным. В некоторых других клетках анафаза В начинается только после того как хроматиды достигают полюсов деления. У некоторых простейших в процессе анафазы В веретено удлиняется в 15 раз по сравнению с метафазным. В растительных клетках анафаза В отсутствует. Телофаза Телофаза Телофаза рассматривается как заключительная стадия митоза; за её начало принимается момент остановки разделённых сестринских хроматид у противоположных полюсов деления клетки. В ранней телофазе наблюдается деконденсация хромосом и, следовательно, увеличение их в объёме. Вблизи сгруппированных индивидуальных хромосом начинается слияние мембранных пузырьков, что дает начало реконструкции ядерной оболочки. Материалом для построения мембран новообразованных дочерних ядер служат фрагменты изначально распавшейся ядерной мембраны материнской клетки, а также элементы эндоплазматического ретикулума. При этом отдельные пузырьки связываются с поверхностью хромосом и сливаются воедино. Постепенно восстанавливается наружная и внутренняя ядерные мембраны, восстанавливаются ядерная ламина и ядерные поры. В процессе восстановления ядерной оболочки дискретные мембранные пузырьки, вероятно, соединяются с поверхностью хромосом без распознавания специфических последовательностей нуклеотидов, так как в результате проведенных экспериментов было выявлено, что восстановление ядерной мембраны происходит вокруг молекул ДНК, заимствованных у любого организма, даже у бактериального вируса. Внутри заново сформировавшихся клеточных ядер хроматин переходит в дисперсное состояние, возобновляется синтез РНК, и становятся различимыми ядрышки. Параллельно с процессами образования ядер дочерних клеток в телофазе начинается и заканчивается разборка микротрубочек веретена деления. Деполимеризация протекает в направлении от полюсов деления к экваториальной плоскости клетки, от минус-концов к плюс-концам. При этом дольше всего сохраняются микротрубочки в средней части веретена деления, которые образуют остаточное тельце Флеминга. Окончание телофазы преимущественно совпадает с разделением тела материнской клетки — цитокинезом. При этом образуются две или более дочерние клетки. Процессы, ведущие к разделению цитоплазмы, берут свое начало еще в середине анафазы и могут продолжаться после завершения телофазы. Митоз не всегда сопровождается разделением цитоплазмы, поэтому цитокинез не классифицируется в качестве отдельной фазы митотического деления и обычно рассматривается в составе телофазы. Различают два основных типа цитокинеза: деление поперечной перетяжкой клетки и деление путём образования клеточной пластинки. Плоскость деления клетки детерминируется положением митотического веретена и проходит под прямым углом к длинной оси веретена. При делении поперечной перетяжкой клетки место разделения цитоплазмы закладывается предварительно ещё в период анафазы, когда в плоскости метафазной пластинки под мембраной клетки возникает сократимое кольцо из актиновых и миозиновых филаментов. В дальнейшем, вследствие активности сократимого кольца, образуется борозда деления, которая постепенно углубляется вплоть до полного разделения клетки. По окончании цитокинеза сократимое кольцо полностью распадается, а плазматическая мембрана стягивается вокруг остаточного тельца Флеминга, состоящего из скопления остатков двух групп полюсных микротрубочек, тесно упакованных вместе с материалом плотного матрикса. Деление путём образования клеточной пластинки начинается с перемещения мелких ограниченных мембраной пузырьков по направлению к экваториальной плоскости клетки. Здесь они сливаются, образуя дисковидную, окружённую мембраной структуру — раннюю клеточную пластинку. Мелкие пузырьки происходят в основном из аппарата Гольджи и перемещаются к экваториальной плоскости вдоль остаточных полюсных микротрубочек веретена деления, образующих цилиндрическую структуру, называемую фрагмопластом. По мере расширения клеточной пластинки микротрубочки раннего фрагмопласта попутно перемещаются к периферии клетки, где за счёт новых мембранных пузырьков продолжается рост клеточной пластинки вплоть до её окончательного слияния с мембраной материнской клетки. После окончательного разделения дочерних клеток в клеточной пластинке откладываются микрофибриллы целлюлозы, завершая образование жёсткой клеточной стенки. Источник: StudFiles.net |
