Пищевая цепочка в море
Как общее правило, можно считать, что при свободном (не паразитическом) питании в пищу различным группам идут организмы, имеющие более высокую способность к размножению, чем их потребители. Таким образом обеспечивается постоянное снабжение кормом даже при интенсивном питании. Рыбы, питающиеся животной пищей, в среднем требуют в течение года удесятерённое количество пищи по отношению к собственному весу. Конечно, от этого имеется множество отклонений в обе стороны. Количество животной пищи, потребляемой другими животными, обычно больше, чем у рыб; у низших групп, видимо, выше, чем у высших. Некоторые кишечнополостные (например, актинии, гидра) требуют пищи в течение года, в сотни раз превышающей их собственный вес. Быстро растущие и размножающиеся животные требуют больше пищи, чем медленно растущие, неподвижные меньше, чем подвижные, и т. д. Те пищевые ресурсы, которые животное потребляет, идут частично на поддержание жизни, на процессы роста, движения и размножения. Все эти функции по-разному осуществляются различными организмами, поэтому так несходны их потребности в пище. Кроме того, все эти процессы идут разным темпом в зависимости от температуры.
В общей схеме можно различать четыре основных звена в пищевых цепях в море (рис. 102).
Первую группу составляют водоросли и бактерии; ко второй можно отнести организмы, питающиеся ими и в свою очередь поедаемые другими животными — звеном третьего порядка. Обе последние группы организмов также идут в пищу животным, составляющим четвёртое звено, т. е. тем, которые из-за тяжёлых раковин, особенностей скелета, крупных размеров и т. д. уже никем в море не поедаются. Звено второго порядка — это мелкие формы зоопланктона, бесчисленные личинки донных животных и рыб. К звену третьего порядка относятся более крупные ракообразные, мелкая рыба (хамса, сардина, сельдь), мелкие головоногие и т. п. Конечное звено составляют большинство морских млекопитающих, а также крупные рыбы, моллюски и ракообразные.
Указанная группировка звеньев пищевой цепи может быть охарактеризована также показателем продуктивности. Двигаясь по нашей схеме слева направо, мы переходим от групп с большой к группам с малой относительной продукцией. Удобнее всего продукцию (Р), т. е. прирост живой массы за определённый отрезок времени, отнести к исходной массе (В) — биомасса (табл. 26).
| Группа | В Баренцовом море | В Каспийском море |
|---|---|---|
| Фитопланктон | 50 | 300 |
| Зоопланктон | 1 | 30 |
| Зообентос | 1/3 | 4 |
| Рыба | 1/6 | 1/2 |
Не лишено интереса, что подобного типа соотношение между различными звеньями пищевой цепи по продуктивности характерно только при явлении свободного, не паразитарного питания живыми организмами. Совсем другие соотношения, как бы зеркально противоположные, устанавливаются при паразитарном питании: крупные, медленно растущие животные служат пищевой базой менее крупным и быстро растущим и, наконец, и те и другие идут в пищу наиболее быстро размножающимся микроорганизмам, причём и здесь эти последние могут, минуя промежуточные звенья, питаться наиболее медленно растущими и крупными организмами (бактерии — позвоночные). Обычно же связи более короткие — цестоды паразитируют в позвоночных, грегарины в беспозвоночных, ракообразные на рыбах, низшие ракообразные на высших и т. п. При питании трупами эта система соотношений расстраивается, и в использовании их равно участвуют все группы.
Среди животных, населяющих море, особенное кормовое значение имеют ракообразные, двустворчатые моллюски и кольчатые черви. Очень мало животных, питающихся губками, кишечнополостными и иглокожими, либо из-за массивного скелета, либо из-за свойственных им иных мер защиты — стрекательного аппарата кишечнополостных, химических веществ тела губок, издающих отталкивающий запах. Выше мы упоминали отом, что жгутиковое одноклеточное — феоцистис иногда образует на обширных пространствах моря цветение, отпугивая своим отвратительным запахом сельдь.
Так как мелкий планктон, в основной части фитопланктон, представляет собой наиболее легко восстанавливающуюся часть морского населения, то на него ориентировано питание очень многих групп морских животных. Кишечнополостные, некоторые группы червей, большинство морских звёзд и офиур, часть брюхоногих и двустворчатых моллюсков, головоногие моллюски, часть полихет — хищники; губки, мшанки, плеченогие, часть полихет, двустворчатые моллюски (в том числе наиболее распространённые мидии и устрицы), часть брюхоногих, низшие ракообразные, некоторые голотурии и оболочниковые питаются наннопланктоном (преимущественно фитопланктоном). Большинство голотурий, некоторые ежи и звёзды, офиуры, часть полихет питаются грунтом, вернее, органическим веществом, и частично бактериями, диатомовыми и другими организмами, которые в нём содержатся. Некоторые беспозвоночные всеядны, как, например, многие морские ежи, часть полихет, некоторые брюхоногие моллюски и часть высших ракообразных.
Если взять какое-нибудь отдельное животное, то можно видеть, что оно всегда является отдельным звеном пищевой цепи, иногда короткой, иногда длинной, причём для всего населения моря можно установить несколько основных типов пищевых связей: бактерии и водоросли поедаются мелкими ракообразными; эти последние — крупными ракообразными и мелкой рыбой, которые идут в пищу крупной рыбе, части китообразных и морским птицам, и, наконец, рыба и морские млекопитающие вылавливаются промыслом.
Эта пищевая цепь состоит из пяти основных звеньев. Такова система питания в пелагиали, для пелагических организмов (рис. 103 и 104).
Очень сходны с этим и пищевые связи у донных рыб, с тем отличием, что у них существенное место в питании занимает, помимо указанных выше групп, также и бентос — черви, моллюски, ракообразные.
Во всех морях продукция планктона превышает продукцию бентоса во много раз, и поэтому планктон представляет собой большую по мощности и более эластичную пищевую базу, нежели бентос. В полном согласии с этим находится как бы «повёрнутость» всей массы рыб в питании в основном на пелагические организмы (рис. 105).
Происходит изменение питания и с возрастом. Так, например, взрослая океаническая сельдь питается более крупными планктонными ракообразными и мелкой рыбой. Сельдь размерами 1,5–13 см потребляет мелких ракообразных, а личинки и мальки размерами 7–12 мм питаются самыми мелкими компонентами планктона — одноклеточными водорослями, инфузориями, мелкими личинками многоклеточных животных и т. п.
Такая же возрастная смена в питании имеет место и у донных рыб. Количество организмов, потребляемых крупными морскими животными, громадно. Кишечник одной крупной сельди может содержать более 60 000 мелких ракообразных. Содержимое желудка большого беззубого кита может весить несколько тонн, а один килограмм его пищи содержит много миллионов поедаемых им ракообразных. Поэтому совершенно понятно, что в тех районах Антарктики, где летом имеются большие скопления ракообразных, будут держаться и кормящиеся ими киты (рис. 106).
Подобным же образом может быть установлена и косвенная зависимость между количеством света и изобилием скумбрии, так как первый обусловливает развитие фитопланктона, а за ним следует обилие зоопланктона, идущего в пищу скумбрии (рис. 107).
Мы указывали, что одному киту для существования в течение одного дня требуются многие миллиарды экземпляров ракообразных — копепод, каждый из которых сам в течение дня поедает свыше 100 000 мелких диатомовых водорослей. Таким образом, для обеспечения существования одного кита в течение одного дня нужны миллиарды экземпляров ракообразных и в сто тысяч раз большее количество диатомовых. Эта своеобразная «пирамида выедания» должна компенсироваться обратной «пирамидой продуктивности» — повышением темпа размножения компонентов низших звеньев пищевой цепи (рис. 108).
Многие рыбы являются типичными бентофагами (питаются бентосом). Типичный бентофаг — морская камбала —питается главным образом червями и моллюсками, пикша — моллюсками, червями и иглокожими.
У большинства рыб в период подготовки к нересту питание ослабляется или совсем приостанавливается. Такая пауза имеется у трески, камбалы, пикши. С другой стороны, некоторые рыбы, например хамса, питаются и во время нереста.
У донных рыб также наблюдается возрастная смена питания, как у пелагических; так, значение моллюсков с возрастом камбалы возрастает, а ракообразных падает. Другие группы существенных изменений по значению в питании камбалы не дают.
Источник: www.bruo.ru
|
Морская пищевая цепь представляет собой сложную систему, в которой мелкие организмы потребляются более крупными. В нижней части пищевой цепи находятся микроскопические растения, а в верхней части — известные хищники, такие как акулы и морские птицы. В зависимости от их размера и места в пищевой цепи, рыба служит различным целям и помогает сбалансировать экосистему разными способами. Производители фитопланктона Основной производитель пищевой цепи называется фитопланктон. Производители создают свою еду. Эти одноклеточные микроскопические растения плавают на поверхности океана, получают энергию от солнца и используют ее для превращения углекислого газа и других питательных веществ в углеводы, которые питают другие виды океана. Фитопланктон, такой как диатомовые водоросли и водоросли, является основой пищевой цепи океана. Они составляют 95 процентов первичных производителей на земле. Зоопланктон Зоопланктон — это маленькие, плавающие животные. Они включают личинок рыб, медуз, микроскопических копепод и мелких донных животных. Они дрейфуют через океан и питаются фитопланктоном. Веслоногие ракообразные составляют большинство зоопланктона. Они составляют большую часть массы животных океана и являются наиболее важной связью между основными производителями и многими крупными животными, питающимися планктоном, такими как мелкая сельдь. Почти все рыбы, обитающие в умеренных или полярных водах, едят веслоногих, чтобы выжить в какой-то момент своей жизни. Маленькие Хищники Следующий уровень в пищевой цепи состоит из мелких хищников, которые питаются рачками и другим планктоном, который они добывают из воды. Моллюски, мелкие ракообразные, такие как креветки и криль, и мелкие рыбы, такие как сардины и сельдь, питаются большим количеством зоопланктона. Крупные стаи мелкой рыбы могут быстро уменьшить популяции планктона, но только временно. Лучшие хищники Крупные хищники, такие как акулы, тунец, кальмар и осьминог, а также морские млекопитающие, такие как тюлени и некоторые киты, образуют вершину пищевой цепи. Птицы и люди также включены в эту группу. Крупные хищники питаются большим разнообразием мелких рыб. Пищевая цепочка начинается снова Пища, которую тратят эти крупные хищники, дрейфует на дно океана, где его питают омары и другие обитатели дна. Часть пищи также разлагается бактериями и возвращается в почву, где растения могут использовать ее питательные вещества. Трата китов и морских черепах, существ, у которых нет непосредственных хищников, также разрушается бактериями. |
|
| Категория: Наука и Техника | Добавил: fantast (26.03.2019) | |
| Просмотров: 5605 | Рейтинг: 4.5/2 |
Источник: www.winstein.org
Пищевые цепи моря
Курсовая работа
Гопаны в нефтях и методика их изучения
Оглавление
Введение
Происхождение гопанов
Структура гопанов
Общие сведения о гопанах
Методика изучения гопанов
Применение гопанов
Заключение
Список литературы
Введение
Целью написания данной курсовой работы является рассмотрение одной из групп углеводородов-биомаркеров — группы гопанов и методики их изучения, в частности хромато-масс-спектрометрии.
Что же такое углеводороды-биомаркеры?
Высокомолекулярные насыщенные углеводороды, обнаруженные в нефтях, унаследовавшие липидную часть живого вещества называются биомаркёрами. В составе исходного живого вещества, участвующего в нефтеобразовании, доминирующая роль принадлежит липоидиновым компонентам (липидам и липоидам). В ходе диагенетических и катагенетических преобразований липиды превращаются в насыщенные алкановые и циклоалкановые, а также моно-, ди-, тетрациклические и полициклические ароматические углеводороды.
Биомаркеры используются, чтобы коррелировать нефти друг с другом и с их нефтематеринскими породами, таким образом, углубляется понимание строения резервуаров, путей миграции и возможных подходов при проведении поисково-разведочных работ. Биомаркеры могут использоваться, чтобы оценить степень преобразованности и (или) биодеградации, таким образом, предоставляя важнейшую информацию, необходимую чтобы правильно оценить распределение и продуктивность бассейнов. Биомаркеры дают важнейшую информацию относительно региональных изменений типов нефтей и нефтематеринских пород, контролирующихся генотипом органического вещества и условиями обстановок осадконакопления. Наконец, биомаркеры предоставляют возможность лучше понять кинетику нефтеобразования и теплового режима бассейна. [4]
В составе биомаркеров в наибольших концентрациях встречаются: нормальные алканы, стераны, трициклические и тетрациклические терпаны, гопаны и некоторые другие углеводороды. В настоящее время определено более 600 биомаркёров, наиболее изученными из которых являются стераны, изопренаны и терпаны.
Происхождение гопанов
Гопаны достаточно распространены в осадках и нефти, потому что гопаноидные предшественники являются важными компонентами мембран в живых клетках, устойчивыми к биодеградации в течение диагенеза.
Основным стуктурным предшественником гопанов является гопаноидный спирт-бактериогопантетрол (рис.1), имеющий 35 атомов углерода в молекуле, и некоторые другие биогопаноиды, которые в процессе диа-, катагенетической эволюции осадков теряют гидроксильные группы (в случае спиртов) или декарбоксилируются (в случае кислот) и в древних осадках, углях, горючих сланцах и нефтях присутствуют в виде углеводородов гомологического ряда гопана. Гопаны в растениях и грибах достаточно редки. Их обнаружили в некоторых тропических деревьях и травах, а также в папоротниках. У бактерий, синезеленых водорослей и лишайников синтез гопаноидов занимает ведущее место в изопреноидном метаболизме. Они входят в состав клеточных мембран и выполняют функции, подобные растительным и животным стеринам.

Рис.1. Происхождение гопанов в нефти из бактериогопантетрола (1) найденный в липидных мембранах прокариотических организмов. Стереохимия обозначена пустой (а) и черной (в) точками (водород направлен в и из страницы. Соответственно). “биологическая” конфигурация [17β (н), 21β (н), 22r] наложена на бактериогопантетрол, и непосредственно насыщает продукт (2) ферментами живых организмов, неустойчива в течение катагенеза и подвергается изомеризации до “геологических” конфигураций (3,4,5). 17β,21α (н) углеводороды (з) называются моретанами, в то время как все остальные гопанами(2,4,5).[4]
Биологическая конфигурация почти плоская, хотя плойчатость связей углерод-углерод в кольцах приводит к трехмерной форме. Подобно стеролам, бактериогопантетрол является амфипатическим, потому что содержит как полярные, так и неполярные концы (рис.2). Неполярный конец гидрофобен, и таким образом, нелегко связывается с водой или другими полярными растворителями. Полярный конец, однако, является гидрофильным, и легко растворим в воде. В клеточной мембране гидрофобные концы липидов расположены внутрь к центру липидного двойного слоя, и полярные концы обращены к водной среде окружающей клетку или цитоплазму внутри клетки. [4]
Рис.2. Бактериогопантетрол [4].
Плоская конфигурация и амфипатический характер необходимы бактериогопантетролу, чтобы встраиваться в липидную мембранную структуру. Так как это стереохимическое расположение термодинамически неустойчиво, диагенез и катагенез бактериогопантетрола приводят к преобразованию 17β (Н), 21β (Н) предшественников в 17α (Н), 21β (Н)-гопаны и 17β (Н), 21α (Н)-моретаны. [1]
Структура гопанов
Структура цилопентанопергидрохризена — основа гопанов, то есть гопан состоит из четырёх сжатых циклогексиловых колец, каждое из которых в форме кресла и сжатого циклопентилового кольца (рис.3).
Рис.3. Структура гопанов [2].
Изопреновые звенья служат основой в структуре (Изопрен (метилбутадиен) — основной структурный блок, состоящий из пяти атомов углерода, который найден во всех биомаркерах). Между изопреновыми звеньями связь в гопанах «голова к хвосту» и именно поэтому они являются регулярными соединениями. Гомологи гопановых углеводородов отличаются всего лишь величиной алкильного заместителя(CH3-С8H17), присоединенной к боковой цепи на Е-кольце. Углеводороды ряда гопана имеют 21 атом углерода в циклах, шесть метильных заместителей в цикле, из которых четыре являются ангулярными. [4]
Общие сведения о гопанах
Собственно говоря, название “гопан” применимо лишь к углеводороду состава С30. Углеводород состава С29 называют норметилгопаном, или адиантаном, состава С27 (отсутствует заместитель при С21)-трисноргопаном. Углеводороды состава С31, С32 и так далее называются соответственно гомогопаном, бисгомогопаном и так далее.
Гопаны с 30 и менее атомами углерода имеют ассиметричные центры в С-21 и на всех кольцевых швах (С-5, С-8, С-9, С — 10, С-13, С-14, С-17, и С-18). Гопаны с более 30 атомами углерода называются гомогопанами, где приставка гомо — касается дополнительных групп метилена в исходной молекуле, гопане. Обычные гомогопаны имеют расширенную боковую цепочку с дополнительным центром в С22 который приводит к двум пикам для каждого гомолога (22R и 22S) на масс-спектограммах этих соединений.
Пара С27 — гопанов (17αН-22,29,30-трисноргопан и 18αН-22,29,30-триноргопан) обычно называют Tm и Ts (соответственно). Считается, что Tm-гопан-биологически образованная структура; Ts-гопан — образовался из него либо посредством диагенетических, либо термальных процессов, или же посредством тех и других.
Гораздо более вероятным является расщепление одиночной связи углерод-углерод между позициями 21 и 22 (приводящие к С27 гопану) или расщепление любой из двух других связей углерод-углерод, присоединенных к С22 приводящему к С29 гопану), чем последовательное расщепление двух связей углерод-углерод.
В стереохимическом плане в гопановых углеводородах наиболее стабильной частью является правая часть молекулы, а именно хиральные центры у С17 и С22 атомов. Гопаны состоят из трех стереоизомерных рядов, а именно: 17α(Н),21β(Н), 17β(Н),21α(Н), 17β(Н),21β(Н)-гопанов. Соединения в ряду βα называются моретанами. (рис. 1)
Обозначения альфа (α) и бета (β) указывают, что водородные атомы ниже или выше плоскости колец.
Существуют два основных типа гопанов: 17αН, 21βН — нефтяной и 17βН, 21βН — биологический. В нефтях, как правило, присутствуют лишь следы гопанов, имеющих “биологическую” конфигурацию. Главная масса гопанов представлена углеводородами 17α,21β ряда. Биогопаны как весьма неустойчивые соединения найдены лишь в торфе, бурых углях, молодых сланцах и рассеянном органическом веществе начальных этапов катагенеза.
Итак, в биогопане сочленение циклов А/В, В/С и С/D — транс, тогда как в геологических гопанах динамически неустойчивое транс-сочленение циклов D/E меняется на более устойчивое — цис. (Сочленение циклов С/D транс — в биогопане и цис — в нефтяном гопане.) Именно из-за термической неустойчивости транс-сочленения D/E и заслонения связей 21-22 и 17-16 (цис-ориентация замещающего радикала) соединения ряда ββ (биологические гопаны) обнаруживают в нефтях в очень небольших количествах. Гопаны С28 αβ редки или отсутствуют в нефтях, так как их образование требует расщепления не одной, а двух связей углерод-углерод, присоединенных к С22 в С35 в гопаноидном предшественнике.
Следует также отметить, что биогопаны имеют более высокую температуру кипения и на хромато-масс-фрагментограммах иллюстрируются значительно позднее стереоизомерных им нефтяных гопанов (например, биогопан состава С30 иллюстрируется в области нефтяного гопана С31).
Различные конфигурации гопанов хорошо заметно в их масс-спектрах. Главным направлением распада гопанов под влиянием электронного удара является разрыв связи 8-14 с образованием двух фрагментов — постоянного А (m/z=191) и переменного (то есть зависящего от молекулярной массы исходного углеводорода) — Б (m/z=149(С27), 163(С28), 177(С29), 191(С30), 205(С31), 219(С32), 233(С33), 247(С34) и так далее.). Кроме того, для гопанов характерными являются молекулярные массы:370, 384, 398, 412, 426, 440 и так далее.
Для нефтяных (17α) гопанов фрагмент А более интенсивен, чем фрагмент Б. Для биогопанов (17β), напротив, интенсивность фрагмента Б выше, чем интенсивность фрагмента А.
Для углеводородов ряда гопана состава С31 и выше, то есть в тех случаях, когда атом С-22 становится хиральным, возможно существование двух эпимеров, отличающихся R- или S-конфигурацией хирильного центра С-22, в исходных биогопанах состава С31 и выше атом С-22 имеет строго определённую R-конфигурацию. В то же время переход к нефтяным гопанам, кроме уже отмечавшейся эпимеризации С-17, сопровождается эпимеризацией центра С-22, что проявляется в ряде дублетных пиков, характерных для хроматограмм смеси нефтяных гопанов. Следует отметить несколько большую устойчивость эпимера с S-конфигурацией. В условиях равновесия, а также в нефтях соотношение 22S- и 22R-эпимеров обычно равно 1.2-1.4. Изомеры, различающиеся конфигурацией С22, несколько напоминают по- свойствам аналогичные эпимеры стеранов, различающихся конфигурацией углеродного атома С20, особенно в βα-диастеранах. [4]
Говорить о биодеградации нефти мы можем если при m/z=177 наблюдается присутствие углеводородов гомологического ряда 25-норгопанов. Например, при биодеградации гопан С28 мы видим как 25,30-бисноргопан, а гопан С29 как 25-норгопанС29.
Методика изучения гопанов
В 1950-1970 годах были заложены основы современной теории нафтидогенеза, охватывающие как геологические, так и химические стороны этого процесса. Считается, что основная масса углеводородов, образующих скопления нефти и газа, формируется на стадии катагенеза, но их количество и состав в значительной степени предопределены особенностями исходной биомассы, количеством и характером превращений органического вещества в диагенезе. Многие углеводороды и близкородственные им соединения, обнаруженные в нефтях, наследуют структуру молекул липидной части живого вещества, а промежуточные продукты их биогеохимических превращений наблюдаются в органическом веществе современных осадков.
Ранее состав и распределение углеводородов-биомаркёров изучался методом газожидкостной хроматографии.
В настоящее время ведущую роль в изучении состава и распределения биометок нефтей, играет метод хроматомасс-спектрометрии. Типичный хроматомасс-спектрометр, используемый для анализов биометок, состоит из газового хроматографа последовательно соединённого с масс-спектрометром. Изучаемая жидкость вводится в газовый хроматограф, где различные составляющие его компоненты разделяются в соответствии со скоростями их перемещения через газово-хроматографическую колонку. Разделение насыщенных углеводородов производится в соответствии с молекулярным весом и летучести компонентов. Разделённые компоненты смеси покидают друг за другом газовый хроматограф и попадают в масс-спектрометрическую ионизационную камеру, где они анализируются в той же последовательности. Далее каждый компонент, попадающий в масс-спектрометр, бомбардируется пучком электронов высокой энергии, ионизирующих молекулы путём выбивания одного электрона (рисунок 4 [2]).
Молекулы и фрагментарные ионы, полученные этим способом, различаются по массе, в большинстве своём имеют одинаковый заряд +1. И только благодаря этому их различию в отношениях молекулярной массы к заряду, вызванных отличием масс, они могут быть разделены в магнитном поле или квадрополе. После чего разделенные ионы подходят к детектору ионного захвата, где относительное количество массы каждого иона записывается. Полная запись содержания и масс всех ионов, образованных из начальных компонент, называется их масс спектром. Собственно масс-спектр изображает зависимость интенсивности сигнала (ордината) от сооотношения массы к заряду (m/z, абцисса) для различных заряженных продуктов, образовавшихся при распаде вещества. Постольку поскольку заряд z обычно равен единице, отношение m/z в действительности отвечает массе иона. Различные классы фрагментарных биомаркёров имеют характерные для каждого из них пути в масс-спектрометре, зависящие от их молекулярной структуры. Фактически, в ХМ-МС вначале получают хроматограмму полного ионного тока (как в газо-жидкостной хроматографии). Затем с помощью квадрупольного масспектрометра возможно разделение этого полного ионного тока на фрагменты определённой массы.
Для отдельного хромотографического пика, вымывающегося из колонки, обычно снимают несколько полных масс-спектров (от трёх до десяти). Для этого необходимо очень быстрое сканирование масс (за время от 0.1 до нескольких секунд) и запись их в цифровом виде на компьютер одновременно с фиксацией времени удерживания соединений. В зависимости от количества соединений, введённых в хроматограф, количество сканирований может быть очень большим и достигать нескольких тысяч. Впоследствии возможна компьютерная обработка полученных данных, то есть получение масс спектров для любого пика хроматограммы. Кроме того, компьютерная обработка масс-спектров позволяет различать индивидуальные соединения, имеющие близкие времена удерживания и элюирующиеся на газовой хроматограмме в виде единого пика. Компьютерная обработка также позволяет просканировать всю хроматограмму по определенным фрагментным или молекулярным ионам и получить так называемые масс-фрагментограммы или масс-хроматограммы. Это бывает полезно, если есть необходимость проследить распределение индивидуальных соединений какого-либо гомологического ряда или соединений, обладающих близким структурным строением. Так, например, тритерпановые хорошо иллюстрируются масс-фрагментограммой по — m/z 191. [2]
Хромато-масс-спектрометрические исследования насыщенных углеводородов проводились на системе, включающей газовый хроматограф 6890, имеющий интерфейс с высокоэффективным масс-селективным детектором Agilent 5973N. Хроматограф снабжен кварцевой капиллярной колонкой длиной 30м, а диаметром 0,25 мм, импрегнированной фазой HP-5MS. В качестве газа-носителя служил гелий со скоростью потока 1мл/мин. Температура испарителя 300°С. Ввод пробы проводился без деления потока. Изометрическая ‘”площадка” длительностью 4 мин. Программирование подъема температуры осуществлялось от 100°С до 290°С со скоростью 4°С в мин с последующей изотермой в течение 20 мин. Ионизирующее напряжение источника-70eV, температура источника — 230°С. Идентификация соединений осуществлялась по временам удерживания, которые могут меняться в зависимости от условий съемки, путем сравнения полученных масс-фрагментограмм с уже имеющимися спекторами в библиотеке системы и опубликованными данными.
При компьютерной обработке хроматограмм очень полезно проводить их реконструкции по наиболее характерному для гопанов фрагментарному иону с m/e 191. Схема распада гопана под воздействием электронного удара представлена на рис.5.
Рис.5. Схема распада гопана под воздействием электронного удара [2].
На распечатанной масс-фрагментограмме можно идентифицировать пики гопанов, стеранов или терпанов по аналогии пользуясь типовой масс-фрагментограммой с пронумерованными пиками и таблицей идентификации пиков. Обрабатываемая и типовая масс-фрагментограммы сравниваются и на первой пронумеровываются пики, на тех же позициях, как на типовой. Высоты пиков измеряются линейкой, для этого проводятся базовые линии, проведённых от вершины пика до пересечения с базовой линией параллельно оси игрек. Типовая масс-хроматограмма терпанов и идентификация пиков представлены на рис.6.
Рис.6. Типовая масс-хроматограмма терпанов [2].
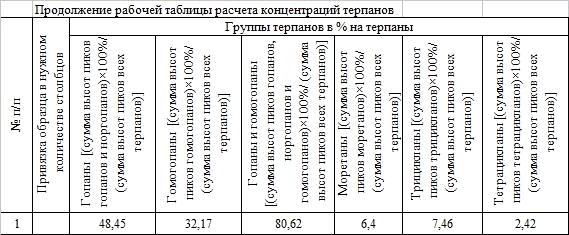
Как пример использования методики хромато-массспектрометрии я приведу лабораторную работу, по идентификации, измерении и расчету содержаний терпанов (m/z 191) (Рис.7).
Результаты были занесены в рабочие таблицы (Табл.1, Табл.2, Табл.3, Табл.4, Табл.5, Табл.6, Табл.7) и был произведен расчет концентраций. Исходя из полученных данных можно сделать вывод о том, что ОВ-незрелое (Ts/Tm<1), но так как мы не видим ярко выраженных биогопанов и рассмотрев отношение изомеров 22S/22R (изомеры 22S встречаются в больших концентрациях), можно сделать вывод, что мы находимся в начале стадии нефтегазообразования (не ниже МК1, начале МК2). Относительно генотипа ОВ сказать точно мы не можем, но так как С35<С34 можно предположить, что ОВ континентального происхождения. Преобладает гопан С30, доля гопанов приблизительно 81%, трицикланов мало (из них преобладают С19 и С20), что тоже говорит о континентальном генезисе, но для полной характеристики нужно рассмотреть также стераны, геологию региона и так далее.
Рис.7.Изучаемая масс-хроматограмма.
Таблица 1.
Таблица 3.
Таблица 4.
Таблица 5.
Таблица 6.
Таблица 7.
Применение гопанов
Сопоставление концентрационного распределение гопанов в нефтях и в РОВ (в битумоидах) позволяет определить источники образования тех или иных нефтей.
В качестве индикатора оценки окислительно-восстановительной обстановки в седиментогенезе и диагенезе используется относительное распределение гомогопанов С31-С35. Соотношение гомогопанов C35/(C31-C35) называют гомогопановый индекс. Относительно высокие концентрации гомогопана С35 указывают на морские условия седиментогенеза и восстановительные условия диагенеза. Относительно низкие концентрации С35 — на субокислительные или слабовосстановительные условия.
Еще один показатель условий образования — соотношение С34/С35. По данным В.А. Каширцева в нефтях морских фаций в ряду гомогопанов С34<С35, в нефтях континентального генезиса — С34>С35. [7]
Об информативности такого показателя, как соотношение С27 норметилгопана (адиантана) к гопану, нет единого мнения. О.А. Арефьев считает, что в нефтях терригенно-карбонатного комплекса величина соотношения адиантан/гопан ниже, чем в нефтях галогено-карбонатного (0,6 и 0,9 соответственно). По его данным для пермских нефтей Восточной Сибири, генерированных материнским веществом континентально-болотных фаций, характерно равное соотношение адиантана и гопана. По соотношению концентраций гопанов С27 и С30 четкой диагностики типов ОВ осуществить не удается, но замечено, что в морских анаэробных обстановках он, как правило, имеет значение ниже 0,5, а значения этого показателя, большее, чем 0,8 встречаются только в неморском ОВ.
Гопаны также используются для оценки степени катагенетических преобразований в ОВ и нефтях, так как в них не происходит перестройки основного углеродного скелета под воздействием температуры. С ростом катагенетической превращенности ОВ в гопанах происходит стереохимическая изомеризация, которая приводит к появлению пространственных изомеров с цис-сочленением колец C/D, трансориентацией алифатического радикала у С-17, и стереоизомеров, с цис-сочленением колец D/E и эпимеров с S-конфигурацией у хирального центра С-22. С учетом этого, для оценки степени катагенетического созревания ОВ используются показатели:
) Соотношение гопанов 17α,21β/17β,21β, то есть процент новообразованных нефтяных гопанов.
) Соотношение гопанов 22S/(22R+22S). Стереохимическая перестройка гопанов, приводящая к увеличению S-конфигурации по отношению к R, активизируется со стадии MK1 (мезокатагенеза), достигая на этаже "нефтяного окна" термодинамического равновесия. В нормально зрелых нефтях, как правило, соотношение S/R в С32 составляет 60/40. Стереохимические перестройки этого типа наиболее четко проявляются в гомогопанах С30-С33.
) Соотношение 17β,21α-моретанов С29+С30 и 17α,21β-гопанов С29+С30. Оно уменьшается с увеличением термической зрелости и достигает 0,05 (равновесный уровень) в сильнопреобразованных нефтях.
Кроме того, очень широко, для оценки степени катагенетической превращенности нефтей используется соотношение Ts/Tm. Пара С27 — гопанов (17α(Н)-22,29,30-трисноргопан и 18α(Н)-22,29,30-триноргопан) обычно называют Tm и Ts(соответственно). Считается, что Tm гопан — биологически образованная структура, а Ts гопан образовался из Tm либо посредством диагенетических, либо термальных процессов, или же посредством тех и других. Таким образом, соотношение Ts/Tm увеличивается с повышением зрелости, и в главной зоне нефтеобразования составляет 1, а на поздних стадиях катагенеза — 5-10. Но в последнее время появился ряд работ, где показаны значительные вариации этого показателя при близкой степени зрелости нефтей, что указывает на влияние в изменении этого соотношения не столько термальных условий преобразования, сколько лито-фациальных условий осадконакопления.
Так, по данным Филпа, Рюлькеттера и Марзи высокие значения соотношения Ts/Tm характерны для нефтей из карбонатных пород, по Робинсону величины этого показателя велики для континентальных нефтей. По мнению Молдована величина этого соотношения контролируется окислительно-восстановительным потенциалом. Значения отношения Ts/Tm увеличиваются при наличии глинистых минералов (катализаторов) в породах, гиперсолёности вод, низких значениях Eh и уменьшаются при более высоких значениях pH среды осадконакопления. Таким образом, применение этого показателя для оценки степени зрелости нефтей желательно проводить только для групп нефтей, генерированных в единых фациальных условиях. [6]
Заключение
гопан углеводород спектрометрия
Изучение биомаркеров в нефтях черезвычайно важно для познания их генезиса и геохимической истории, для повышения достоверности прогноза нефтегазоносности, т.к. сложные структуры биомаркеров дают подробную информацию относительно их происхождения.
В данной курсовой работе была рассмотрена группа гопанов. Структура этих соединений представляет собой четыре сжатых циклогексиловых кольца, каждое из которых в форме кресла и сжатое циклопентиловое кольцо. Основным стуктурным предшественником гопанов является гопаноидный спирт — бактериогопантетрол, имеющий 35 атомов углерода в молекуле. Существуют два основных типа гопанов: 17αН, 21βН — нефтяной и 17βН, 21βН — биологический, а также ряд соединений 17β(Н),21α(Н), которые называются моретанами. (рис. 1)
В нефтях, как правило, присутствуют лишь следы биологических гопанов, а основная масса представлена геогопанами.
Также был раскрыт актуальный сейчас метод хромато-масспектрометрии, приведена типовая хроматограмма и таблица с идентификацией пиков. Для оценки степени катагенетической превращенности нефтей используется соотношение Ts/Tm. Для определения генотипа ОВ мы должны использовать полученные показатели и отношения в совокупности с другими геологическими показателями и геологией региона. Например, показатель условий образования — соотношение С34/С35. В нефтях морских фаций в ряду гомогопанов С34<С35, в нефтях континентального генезиса — С34>С35. Или, например, относительно высокие концентрации гомогопана С35 указывают на морские условия седиментогенеза и восстановительные условия диагенеза, а относительно низкие концентрации С35 — на субокислительные или слабовосстановительные условия.
Гопаны применяются для решения очень многих задач, например для корреляции систем нефть-нефть или для системы нефть-материнские породы. Сопоставление концентрационного распределения гопанов в нефтях и рассеянном органическом веществе позволяет определить источники образования тех или иных нефтей, также они позволяют определить степень преобразованности нефти, и используются в качестве индикатора оценки окислительно-восстановительной обстановки в седиментогенезе и диагенезе, также с помощью изучения гопанов проводятся палеореконструкции в целях определения некоторых факторов: источники, созревание, биодеградация. Относительное распределение гопанов рассматривается как характерный “отпечаток пальцев” нефтей региона и поэтому представляет интерес сводка этих данных по различным месторождениям земного шара.
Отдельно взятый показатель не дает достоверной информации о перспективах нефтегазоносности и продуктивности региона, нужно рассматривать в совокупности все геохимические показатели и учитывать геологию этого региона.
Список литературы
1. В.А. Каширцев, Р.П. Филп, Дж. Аллен, А. Гальвес-Синибальди, И.Н. Зуева, О.Н. Чалая, И.Н. Андреев “Биодеградация биомаркеров в природных битумах Оленекского поднятия” 1993
2. Методическое руководство по хромато-масспектрометрии
. Б. Тиссо, Д.Вельте “Образование и распространение нефти” Москва, “Мир”, 1981
4. Peters K.E., Moldowan J.M. The biomarker guide. — Prentice Hall: Englewood Cliffs. New Jersy, 1993, 353p
5. Семинары по предмету “Введение в геохимию нефти и газа”
. Костырева Е.А. Геохимия и генезис палеозойских нефтей юго-востока Западной Сибири // Новосибирск: Изд-во СО РАН. Филиал "Гео", 2005, 183 с
7. Каширцев В.А. Органическая геохимия нафтидов востока Сибирской платформы. Якутск: СО РАН, ЯФ, 2003. — 160 с.
Источник: www.BiblioFond.ru