Эукариоты представители
К прокариотическим организмам относятся бактерии — прежде всего бактерии в традиционном смысле этого термина, затем сине-зеленые водоросли (цианобактерии) и недавно открытые зеленые водорослеподобные организмы (хлороксибактерии), а также некоторые многоклеточные организмы, такие как актинобактерии (актиномицеты) и миксобактерии, образующие плодовые тела.
Все это — микробы. Название «прокариоты» происходит от греческих слов pro (перед) и karyon (семя, ядро). Прокариотические клетки в большинстве своем меньше эукариотических. Прокариотическую структуру, несущую гены, иногда неправильно называемую бактериальной хромосомой, следует называть генофором. Это кольцевая цепь ДНК, которая не находится в ядре, окруженном мембраной; в электронном микроскопе генофор выглядит как сравнительно прозрачная область, которую называют нуклеоидом. В эукариотической клетке носителями генов служат хромосомы, находящиеся в ядре, ограниченном мембраной. В исключительно тонких, прозрачных препаратах можно видеть живые хромосомы с помощью светового микроскопа; чаще же их изучают в фиксированных и окрашенных клетках (в отличие от генофора прокариот хромосомы окрашиваются в красный цвет реактивом Фёльгена).
омосомы построены из ДНК, которая находится в комплексе с пятью гистонами-белками, богатыми аргинином и лизином и составляющими у большинства эукариот значительную часть массы хромосом (более половины). Гистоны придают хромосомам ряд характерных свойств — упругость, способность к компактной укладке и окрашиваемость. Однако они непричастны к способности хромосом передвигаться, за которую ответственны митотическое веретено или сходные с ним системы микротрубочек.
Все широко известные организмы — водоросли, простейшие, плесени, высшие грибы, животные и растения — состоят из эукариотических клеток. Клетки этих организмов (за исключением некоторых протоктистов) делятся путем митоза — так называемым непрямым делением, при котором хромосомы продольно «расщепляются» и расходятся двумя группами к противоположным полюсам клетки. Слово митоз в этой книге будет употребляться в классическом смысле — только тогда, когда речь идет о хромосомах и митотическом аппарате; в это понятие не входит точное прямое распределение генов, составляющих группу сцепления (генофор) у бактерий. Прокариотические клетки могут делиться перетяжкой на равные части или почкованием на неравные части, но они никогда не делятся путем митоза.
Прокариоты обычно размножаются бесполым способом.
многих из них половой процесс вообще неизвестен и потомство имеет только одного родителя (в этой книге под половым размножением понимается любой процесс, при котором каждый потомок имеет более чем одного родителя — обычно двух). У прокариот, способных к половому размножению, половые системы однонаправленны в том смысле, что клетки-доноры («мужские») передают свои гены клеткам-реципиентам («женским»). Число передаваемых генов варьирует от одной конъюгации к другой: гены образуют длинную молекулу ДНК, и обычно передается лишь небольшая часть генома (но иногда — почти весь геном). При конъюгации бактерий не происходит слияния цитоплазмы клеток, как это бывает у всех животных, у грибов (при слиянии гиф) и у многих растений и протоктистов. Новый прокариотический организм, называемый рекомбинантом, состоит из самой клетки-реципиента, в которой некоторые гены замещены генами донора. Таким образом, у прокариот родители почти никогда не вносят равного вклада. С другой стороны, у эукариотической клетки, возникающей половым путем (зиготы), вклад родителей одинаков или почти одинаков: новая эукариотическая особь обычно получает половину генов и некоторое количество нуклеоплазмы и цитоплазмы от каждого родителя.
Хромосомы построены из ДНК и белков, но препараты изолированных хромосом часто содержат также значительную примесь РНК из других областей ядра. Эта РНК, вероятно, как информационная, так и рибосомная — легко прилипает к изолированным хромосомам. Эукариотическое ядро содержит также ядрышки, состоящие из предшественников цитоплазматических рибосом — из цепей РНК различной длины и большого числа белков. Другие органеллы, свойственные только эукариотическим клеткам, — это митохондрии, пластиды, центриоли и кинетосомы с их ундулиподиями. За исключением микротрубочек, которые встречаются как внутри, так и вне ядра, все эти органеллы лежат с наружной стороны от ядерной мембраны.
Все двигательные органеллы эукариотической клетки имеют толщину около 0,25 мкм; из них более длинные (от 10 до 15 мкм) и представленные у каждой клетки в небольшом числе по традиции называют жгутиками, а более короткие и многочисленные-ресничками. Электронная микроскопия выявила поразительное структурное сходство всех эукариотических ресничек и жгутиков: на поперечном разрезе во всех случаях можно видеть одно и то же расположение белковых микротрубочек (9 + 2), диаметр каждой из которых составляет около 0,024 мкм. Эти органеллы гораздо сложнее бактериальных жгутиков и имеют совершенно иную структуру и иной белковый состав. Пришло время, чтобы их названия отразили новые сведения; поэтому в нашей книге для ресничек, жгутиков и родственных им органелл эукариот (например, для осевой нити в хвосте спермия, для структурных единиц цирруса у инфузорий и других структур типа 9 + 2 и их производных, развивающихся из кинетосом, которые сами имеют на поперечном разрезе структуру 9 + 0) используется термин ундулиподия. Название жгутик оставлено для тонких бактериальных жгутиков и гомологичных им структур, таких как аксиальные фибриллы спирохет; обычно жгутики слишком малы, чтобы их можно было видеть с помощью обычного светового микроскопа. Эта менее двусмысленная терминология основана на соображениях Т. Яна и его коллег.
Общеизвестные прокариоты и эукариоты
|
Прокариоты |
Эукариоты |
|
|
Одноклеточные гетеротрофы |
Настоящие бактерии: сероводородные бактерии, Е. coli, псевдомонады, некоторые железобактерии, бациллы, метанообразующие бактерии, азотфиксирующие бактерии, спирохеты, микоплазмы, риккетсии, Chlamydia, Bedsonia |
Протисты: амебы, радиолярии, фораминиферы, инфузории, споровики, некоторые динофлагелляты. Некоторые дрожжи |
|
Автотрофы |
Сине-зеленые и зеленые прокариотические водоросли (т. е. цианобактерии и хлороксибактерии), другие фотосинтезирующие бактерии, хемоавтотрофные бактерии |
Водоросли: красные, бурые, харовые, диатомовые; некоторые динофлагелляты, хлореллы, Cyanidium. Растения: мхи, печеночники, папоротники, цикадовые, хвойные, цветковые |
|
Мицелиальные и многоклеточные организмы |
Актинобактерии (актиномицеты), некоторые скользящие и почкующиеся бактерии |
Водные плесени, хитриды, шляпочные грибы, дождевики, аскомицеты, слизевики. Растения. Животные: губки, гребневики, кишечнополостные, плеченогие, мшанки, аннелиды, брюхоногие моллюски, членистоногие, иглокожие, оболочники, рыбы, млекопитающие |
Различия между прокариотами и эукариотами
|
Признаки |
Прокариоты |
Эукариоты |
|
Размеры клеток |
Клетки большей частью мелкие (1-10 мкм); некоторые больше 50 мкм |
Клетки большей частью крупные (10-100 мкм); некоторые больше 1 мм |
|
Общие особенности |
Исключительно микроорганизмы. Одноклеточные или колониальные. Морфологически наиболее сложны нитчатые или мицелиальные формы с «плодовыми телами». Нуклеоид без пограничной мембраны |
Некоторые — микроорганизмы; большинство-крупные организмы. Одноклеточные, колониальные, мицелиальные или многоклеточные. Морфологически наиболее сложны позвоночные животные и покрытосеменные растения. Все имеют ядро с пограничной мембраной |
|
Клеточное деление |
Немитотическое, прямое, чаще всего путем расщепления надвое или почкования. Генофор содержит ДНК, но не содержит белка; не дает реакции Фёльгена. Центриолей, митотического веретена и микротрубочек нет |
Различные формы митоза. Обычно имеется много хромосом, содержащих ДНК, РНК и белки и дающих ярко-красную окраску по Фёльгену. У многих форм имеются центриоли, а также. митотическое веретено или упорядоченно расположенные микротрубочки |
|
Системы пола |
У большинства форм отсутствуют; если имеются, то осуществляют однонаправленный перенос генетического материала от донора к реципиенту |
У большинства форм имеются; равное участие обоих родителей в оплодотворении |
|
Развитие |
Отсутствует многоклеточное развитие, начинающееся от диплоидных зигот; нет выраженной дифференциации тканей. Только одиночные или колониальные формы. Нет сложных межклеточных соединений. Метаморфоз редок |
Гаплоидные формы образуются в результате мейоза, диплоидные развиваются из зигот; у многоклеточных — далеко идущая дифференциация тканей. Плазмодесмы, десмосомы и другие сложные межклеточные соединения. Метаморфоз обычен |
|
Устойчивость к кислороду |
Строгие или факультативные анаэробы, микроаэрофилы или аэробы |
В основном аэробы. Исключения — явно вторичные модификации |
|
Обмен веществ |
Различные схемы метаболизма; нет специализированных, ограниченных мембранами органелл с ферментами, предназначенными для окисления органических молекул (нет митохондрий) |
Во всех царствах одинаковая схема окислительного метаболизма: имеются мембранные органеллы (митохондрии) с ферментами окисления трикарбоновых органических кислот |
|
Фотосинтез (если он имеется); липиды и др. |
Ферменты фотосинтеза связаны с клеточными мембранами (хроматофоры), а не упакованы в виде отдельных органелл. Встречается анаэробный и аэробный фотосинтез с выделением серы, сульфата или кислорода. Донорами водорода могут быть H2, H2O, H2S или (H2CO)n. Липиды: вакциновая и олеиновая кислоты, гопаны; стероиды крайне редки. Образуют аминогликозидные антибиотики |
Ферменты фотосинтеза находятся в пластидах, ограниченных мембранами. Большей частью фотосинтез с выделением кислорода; донором водорода всегда служит H2O. Липиды: обычны линолевая и линоленовая кислоты, стероиды (эргостерол, циклоартенол, холестерол). Обычны (особенно у растений) алкалоиды, флавоноиды, ацетогенины и другие вторичные метаболиты |
|
Двигательные приспособления |
Некоторые имеют простые бактериальные жгутики, состоящие из флагеллина; другие передвигаются путем скольжения. Внутриклеточное движение встречается редко или отсутствует; нет фагоцитоза, пиноцитоза и циклоза |
У большинства имеются ундулиподии: «жгутики» или реснички типа 9 + 2. Структуры 9 + 0 или 6 + 0 представляют собой эволюционные модификации схемы 9 + 2. Обычны псевдоподии, содержащие актиноподобный белок. Характерно внутриклеточное движение (пиноцитоз, фагоцитоз, циклоз), осуществляемое с помощью специализированных белков — актина, миозина, тубулина |
|
Клеточная стенка |
Гликопептиды — производные диаминопимелиновой и мурамовой кислот; гликопротеиды редки или отсутствуют; аскорбиновая кислота не требуется |
Хитин или целлюлоза; обычны гликопротеиды с гидроксилированными аминокислотами; необходима аскорбиновая кислота |
|
Споры |
Устойчивы к высыханию; теплоустойчивые эндоспоры содержат дипиколинат кальция; актиноспоры |
Сложные, варьируют в зависимости от типа; нет дипиколината кальция; в спорах спорополленин; эндоспор нет |
Источник: Л. Маргелис. Роль симбиоза в эволюции клетки. Пер. В.Б. Касинова, Е.В. Кунина. Под ред. Б.М. Медникова. Издательство «Мир». Москва. 1963
Источник: www.activestudy.info
СТРОЕНИЕ И ХИМИЧЕСКИЙ СОСТАВ КЛЕТОК
Живую природу делят на несколько царств. Наиболее распространены два подхода: деление на царства прокариот и эукариот, а также на царства протистов, растений и животных. Особое царство Vira составляют вирусы и фаги. Это бесклеточные формы, занимающие промежуточное положение между живой и неживой материей. Любые организмы размером не более 100-500 мкм называют микроорганизмами. Их положение в системах деления живого мира показано на рис. 1.1.
По строению клеток выделяют две группы организмов: прокариоты и эукариоты.
Прокариоты не имеют окруженного мембраной ядра. ДНК в виде замкнутой в кольцо молекулы свободно располагается в цитоплазме, отсутствуют цитоплазматические органеллы, окруженные элементарной мембраной, рибосомы меньше, чем у эукариот.
Эукариоты имеют истинное ядро, в котором содержится основная часть генома (набора хромосом). Небольшая часть клеточного генома содержится в других органеллах (митохондриях, хлоропластах) в виде кольцевых молекул ДНК. Рибосомы крупнее, чем у прокариот.
Царство животных объединяет многоклеточные эукариотные организмы, которые питаются готовыми органическими веществами (гетеротрофно). Животные осуществляют голозойное питание, заключающееся в захватывании и переваривании твердой пищи. В микромир из царства животных попадают коловратки и черви.
Царство растений включает многоклеточные эукариотные организмы, питающиеся неорганическими веществами (автотрофно), из которых синтезируют органические вещества биомассы, используя энергию солнечного света (фотосинтез).
Царство протистов (самых простых) – все организмы, у которых отсутствует дифференцировка на органы и ткани. Большинство протистов являются одноклеточными организмами. Протисты делятся на две группы: высшие протисты и низшие протисты.
Высшие протисты имеют эукариотные клетки. К ним относятся простейшие, водоросли и низшие грибы.
Простейшие – одноклеточные животные, размножаются простым делением клетки, для окисления потребляемых органических веществ используют молекулярный кислород. Простейшие делятся на 3 класса: саркодовые, жгутиковые, инфузории. По своему размеру (10-200 мкм) все простейшие относят к микроорганизмам.

Рис.1.1. Положение микроорганизмов в схеме живого мира
Водоросли – микроскопические фотосинтезирующие организмы (в основном одноклеточные), которые в отличие от растений не имеют ни корней, ни стеблей, ни листьев. Окраска водорослей зависит от количественного соотношения различных пигментов: зеленого – хлорофилла, синего – фикоциана, бурого – фукоксантина, красного – фикоэритрина, оранжевых – каротина и ксантофилла.
Грибы – одноклеточные и мицелиальные органотрофные организмы, питающиеся растворенными органическими веществами (осмотрофный тип питания). К микроорганизмам относятся низшие грибы (фикомицеты) и дрожжи (почкующиеся грибы).
Низшие протисты – это одноклеточные микроорганизмы, имеющие прокариотный тип клетки. По современной классификации все прокариоты отнесены к бактериям. Следовательно, термины «низшие протисты», «прокариоты», «бактерии» являются тождественными. Большинство бактериальных клеток имеют форму сферы, цилиндра или спирали. Они бывают одиночными или образуют нити и колонии. Бактерии сферической формы называют кокками. Если такие клетки после деления не расходятся, то образуются пары кокков (диплококки), цепочки (стрептококки), пакеты правильной формы (сарцины) или колонии сферической формы (рис. 1.2). Клетки, имеющие форму цилиндра, называются палочковидными. Спиралевидные клетки классифицируются в зависимости от числа витков: у спирилл – от одного до нескольких витков, спирохеты имеют множество витков (свыше трех), вибрионы выглядят как изогнутые палочки (неполный виток спирали). Встречаются бактерии и самой разнообразной формы (рис. 1.2).
Рис. 1.2. Формы бактериальных клеток
1 – кокки; 2 – диплококки; 3 – сарцины; 4 – стрептококки; 5 – колония сферической формы: 6 – палочковидные бактерии (одиночная клетка и цепочка клеток); 7 – спириллы; 8 – вибрион; 9 – бактерии, имеющие форму замкнутого или незамкнутого кольца; 10 – бактерии, образующие выросты (простеки); 11 – бактерия червеобразной формы; 12 – бактериальная клетка в форме шестиугольной звезды; 13 – представитель актиномицетов; 14 – плодовое тело миксобактерии; 15 – нитчатая бактерия рода Caryophanon с латерально расположенными жгутиками: 16 – нитчатая цианобактерия. образующая споры (акинеты) и гетероцисты; 8, 15, 17, 18 – бактерии с разными типами жгутикования; 19 – бактерии, образующая капсулу; 20 – нитчатые бактерии группы Sphaeroillus, заключенные в чехол, инкрустированный гидратом окиси железа; 21 – бактерия, образующая шипы; 22 – Galionella
Для названия микроорганизмов принята бинарная номенклатура: первое слово обозначает род и пишется с прописной буквы, второе пишется со строчной буквы и определяет вид. Название дается на латинском языке, например, Escherichia coli или сокращенно E. coli (кишечная палочка).
Основной единицей в систематике микроорганизмов является вид. Вид – это совокупность особей одного генотипа, обладающих ярко выраженным фенотипическим (внешним) сходством. В генетике микроорганизмов существует также понятие «штамм» и «клон». Штамм – это генетически однородная культура, поддерживаемая с помощью отбора наследственно однородных клеток. Клон – это генетически однородное потомство одной клетки, имеющей одно ядро (для эукариот) или один нуклеоид (для прокариот).
При классификации микроорганизмов основные единицы распределяют по группам более высокого порядка: виды объединяют в роды, роды – в семейства, семейства – в порядки, порядки – в классы, классы – в отделы.
Источник: studopedia.su
Строение эукариотической клетки
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органеллы (другое название, реже употребляемое в научной литературе[комм. 1], — органоиды), из которых многие отделены от цитоплазмы одной или несколькими мембранами (в прокариотических клетках внутренние органоиды, окруженные мембраной, встречаются редко). Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
Деление на царства
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных. Затем было выделено царство грибов, которые, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств из-за биохимических особенностей. Также некоторые авторы выделяют царства протистов или простейших и хромистов. Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалье-Смита, все эукариоты подразделяются на два монофилетических таксона (поддомена) — униконтов (Unikonta) и биконтов (Bikonta). Положение таких эукариот, как Collodictyon и Diphylleia rotans, на данный момент не определено.
Отличия эукариот от прокариот
Важнейшая, основополагающая особенность эукариот связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой. ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие, среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать достаточно прочную клеточную стенку из одной молекулы. Это обстоятельство могло заставить их использовать для клеточной стенки иной материал. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие — строение жгутиков. У бактерий жгутиками являются полые нити диаметром 15—20 нм из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотических жгутиков жгутики эукариот изгибаются или извиваются.
Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5—10 мкм, когда тот же показатель у эукариот составляет 10—100 мкм. Объём такой клетки в 1000—10 000 раз больше, чем прокариотической.
Рибосомы прокариот мелкие (70S-типа). Клетки эукариот содержат как более крупные рибосомы 80S-типа, находящиеся в цитоплазме, так и 70s-рибосомы прокариотного типа, расположенные в митохондриях и пластидах.
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд лет назад, от них около 1,2 млрд лет назад произошли эукариотические организмы.
Примечания
Комментарии
Источники
Литература
На русском языке:
- Галицкий В. А. Возникновение эукариотических клеток и происхождение апоптоза // Цитология, 2005, т. 47, вып. 2, с. 103—120.
- Биологический энциклопедический словарь / под редакцией М. С. Гилярова. — М., 1989.
- Мирабдуллаев И. М. Проблема происхождения эукариот // Успехи совр. биол. 1989а. Т. 107. С. 341—356.
- Марков А. В. Проблема происхождения эукариот // Палеонтологический журнал 2 (2005): 3—12.
- Подборка статей по проблеме происхождения эукариот
- Вся Биология — Эукариоты
- Медников Б. М. Биология: формы и уровни жизни. — М.: Просвещение, 1995.
- Тейлор Д., Грин Н., Стаут У. Биология. В трёх томах
- Малахов В. В. Основные этапы эволюции эукариотных организмов. — 2003
- Федонкин М. А. Сужение геохимического базиса жизни и эвкариотизация биосферы: причинная связь. — 2003
- Шестаков С. В. О ранних этапах биологической эволюции с позиции геномики. — 2003
- Марков А. В. Проблема происхождения эукариот
- Марков А. В, Куликов А. М. Происхождение эвкариот: выводы из анализа белковых гомологий в трёх надцарствах живой природы
- Заварзин Г. А. Эволюция микробных сообществ.
- Колчанов Н. А. Эволюция регуляторных генетических систем.
- Розанов А. Ю., Федонкин М. А.. Проблема первичного биотопа эвкариот. — 1994.
- Богданов Ю. Ф., Дадашев С. Я., Гришаева Т. М. Сравнительная геномика и протеомика дрозофилы, нематоды Бреннера и арабидопсиса. Идентификация функционально сходных генов и белков синапсиса мейотических хромосом
- Ермилова Е. В., Залуцкая Ж. М., Лапина Т. В. Подвижность и поведение микроорганизмов. Т. 2: Эукариоты
- Греннер Д., Марри Р., Мейес П., Родуэлл В. Биохимия человека
На других языках:
- Bisby FA, Roskov YR, Ruggiero MA, Orrell TM, Paglinawan LE, et al. Species 2000 & ITIS catalogue of life: 2007 annual checklist. Species 2000. Retrieved Jan. 2007. 21, 2008
- Patterson DJ. The diversity of eukaryotes. Am Nat. 1999
- Stechmann A, Cavalier-Smith T. Rooting the eukaryote tree by using a derived gene fusion. Science. 2002
- Richards TA, Cavalier-Smith T. Myosin domain evolution and the primary divergence of eukaryotes. Nature. 2005
- Stechmann A, Cavalier-Smith T. Phylogenetic analysis of eukaryotes using heat-shock protein Hsp90. J Mol Evol. 2003
- Makiuchi T, Nara T, Annoura T, Hashimoto T, Aoki T. Occurrence of multiple, independent gene fusion events for the fifth and sixth enzymes of pyrimidine biosynthesis in different eukaryotic groups. Gene. 2007
- Kim E, Simpson AGB, Graham LE. Evolutionary relationships of apusomonads inferred from taxon-rich analyses of 6 nuclear encoded genes. Mol Biol Evol. 2006
- Nozaki H, Matsuzaki M, Misumi O, Kuroiwa H, Higashiyama T, et al. Phylogenetic implications of the CAD complex from the primitive red alga Cyanidioschyzon merolae (Cyanidiales, Rhodophyta). J Phycol. 2005
- Adl SM, Simpson AGB, Farmer MA, Andersen RA, Anderson OR, et al. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. J Eukaryot Microbiol. 2005
- Keeling PJ, Burger G, Durnford DG, Lang BF, Lee RW, et al. The tree of eukaryotes. Trends Ecol Evol. 2005
- Simpson AGB, Roger AJ. The real ‘kingdoms’ of eukaryotes. Curr Biol. 2004
- Parfrey LW, Barbero E, Lasser E, Dunthorn M, Bhattacharya D, et al. Evaluating support for the current classification of eukaryotic diversity. PLoS Genet. 2006
- Burki F, Shalchian-Tabrizi K, Minge M, Skjaeveland A, Nikolaev SI, et al. Phylogenomics reshuffles the eukaryotic supergroups. PLoS ONE. 2007
- Bodyl A. Do plastid-related characters support the chromalveolate hypothesis? J Phycol. 2005
- Stiller JW, Riley J, Hall BD. Are red algae plants? A critical evaluation of three key molecular data sets. J Mol Evol. 2001
- Grzebyk D, Katz ME, Knoll AH, Quigg A, Raven JA, et al. Response to comment on «The evolution of modern eukaryotic phytoplankton». Science. 2004
- Yoon HS, Grant J, Tekle YI, Wu M, Chaon BC, et al. Broadly sampled multigene trees of eukaryotes. BMC Evol Biol. 2008
- Jarvis P, Soll M. Toc, Tic, and chloroplast protein import. Biochim Biophys Acta. 2001
- Marin B, Nowack ECM, Melkonian M. A plastid in the making: primary endosymbiosis. Protist. 2005
- Nowack ECM, Melkonian M, Glockner G. Chromatophore genome sequence of Paulinella sheds light on acquisition of photosynthesis by eukaryotes. Curr Biol. 2008
- Theissen U, Martin W. The difference between organelles and endosymbionts. Curr Biol. 2006
- Bhattacharya D, Archibald JM. The difference between organelles and endosymbionts — response to Theissen and Martin. Curr Biol. 2006
- Okamoto N, Inouye I. The katablepharids are a distant sister group of the Cryptophyta: a proposal for Katablepharidophyta divisio nova/Kathablepharida phylum novum based on SSU rDNA and beta-tubulin phylogeny. Protist. 2005
- Andersen RA. Biology and systematics of heterokont and haptophyte algae. Am J Bot. 2004
- Cavalier-Smith T. Principles of protein and lipid targeting in secondary symbiogenesis: euglenoid, dinoflagellate, and sporozoan plastid origins and the eukaryote family tree. J Eukaryot Microbiol. 1999
- Graham LE, Wilcox LW. Algae. Upper Saddle River, NJ: Prentice Hall; 2000
- Schnepf E, Elbrachter M. Dinophyte chloroplasts and phylogeny: a review. Grana. 1999
- Kohler S, Delwiche CF, Denny PW, Tilney LG, Webster P, et al. A plastid of probable green algal origin in apicomplexan parasites. Science. 1997
- Kohler S. Multi-membrane-bound structures of Apicomplexa: I. the architecture of the Toxoplasma gondii apicoplast. Parasitol Res. 2005
- Hopkins J, Fowler R, Krishna S, Wilson I, Mitchell G, et al. The plastid in Plasmodium falciparum asexual blood stages: a three-dimensional ultrastructural analysis. Protist. 1999
- Tomova C, Geerts WJC, Muller-Reichert T, Entzeroth R, Humbel BM. New comprehension of the apicoplast of Sarcocystis by transmission electron tomography. Biol Cell. 2006
- Moore RB, Obornik M, Janouskovec J, Chrudimsky T, Vancova M, et al. A photosynthetic alveolate closely related to apicomplexan parasites. Nature. 2008
- Stiller JW, Reel DC, Johnson JC. A single origin of plastids revisited: convergent evolution in organellar genome content. J Phycol. 2003
- Larkum AWD, Lockhart PJ, Howe CJ. Shopping for plastids. Trends Plant Sci. 2007
- McFadden GI, van Dooren GG. Evolution: red algal genome affirms a common origin of all plastids. Curr Biol. 2004
- Stiller JW, Hall BD. The origin of red algae: implications for plasmid evolution. Proc Natl Acad Sci U S A. 1997
- Sanchez-Puerta MV, Bachvaroff TR, Delwiche CF. Sorting wheat from chaff in multi-gene analyses of chlorophyll c-containing plastids. Mol Phylogenet Evol. 2007
- Falkowski PG, Katz ME, Knoll AH, Quigg A, Raven JA, et al. The evolution of modern eukaryotic phytoplankton. Science. 2004
- Fast NM, Kissinger JC, Roos DS, Keeling PJ. Nuclear-encoded, plastid-targeted genes suggest a single common origin for apicomplexan and dinoflagellate plastids. Mol Biol Evol. 2001
- Bucknam J, Boucher Y, Bapteste E. Refuting phylogenetic relationships. Biol Direct. 2006
- Gupta RS, Golding GB. Evolution of HSP70 gene and its implications regarding relationships between archaebacteria, eubacteria, and eukaryotes. J Mol Evol. 1993
- Gupta RS, Singh B. Phylogenetic analysis of 70 kD heat shock protein sequences suggests a chimeric origin for the eukaryotic cell nucleus. Curr Biol. 1994
- Gomez-Lorenzo MG, Spahn CMT, Agrawal RK, Grassucci RA, Penczek P, et al. Three-dimensional cryo-electron microscopy localization of EF2 in the Saccharomyces cerevisiae 80S ribosome at 17.5 angstrom resolution. EMBO J. 2000
- Jorgensen R, Merrill AR, Andersen GR. The life and death of translation elongation factor 2. Biochem Soc Trans. 2006
- Moreira D, Le Guyader H, Philippe H. The origin of red algae and the evolution of chloroplasts. Nature. 2000
- Germot a, Philippe H. Critical analysis of eukaryotic phylogeny: a case study based on the HSP70 family. J Eukaryot Microbiol. 1999
- Philippe H, Delsuc F, Brinkmann H, Lartillot N. Phylogenomics. Annu Rev Ecol Evol Syst. 2005
- Wiens JJ. Missing data and the design of phylogenetic analyses. J Biomed Inform. 2006
- Philippe H, Snell EA, Bapteste E, Lopez P, Holland PWH, et al. Phylogenomics of eukaryotes: Impact of missing data on large alignments. Mol Biol Evol. 2004
- Patron NJ, Inagaki Y, Keeling PJ. Multiple gene phylogenies support the monophyly of cryptomonad and haptophyte host lineages. Curr Biol. 2007
- Hackett JD, Yoon HS, Li S, Reyes-Prieto A, Rummele SE, et al. Phylogenomic analysis supports the monophyly of cryptophytes and haptophytes and the association of Rhizaria with Chromalveolates. Mol Biol Evol. 2007
- McFadden GI. Primary and secondary endosymbiosis and the origin of plastids. J Phycol. 2001
- Rodriguez-Ezpeleta N, Brinkmann H, Burey SC, Roure B, Burger G, et al. Monophyly of primary photosynthetic eukaryotes: green plants, red algae, and glaucophytes. Curr Biol. 2005
- Nosenko T, Bhattacharya D. Horizontal gene transfer in chromalveolates. BMC Evol Biol. 2007
- Lane CE, van den Heuvel K, Korera C, Curtis BA, Parsons BJ, et al. Nucleomorph genome of Hemiselmis andersenii reveals complete intron loss and compaction as a driver of protein structure and function. Proc Natl Acad Sci U S A. 2007
- Douglas S, Zauner S, Fraunholz M, Beaton M, Penny S, et al. The highly reduced genome of an enslaved algal nucleus. Nature. 2001
- Vørs N. Ultrastructure and autecology of the marine, heterotrophic flagellate Leucocryptos marina (Braaud) Butcher 1967 (Kathablepharidaceae/Kathablepharidae), with a discussion of the genera Leucocryptos and Katablepharis/Kathablepharis. Eur J Protistol. 1992
- McFadden GI, Gilson PR, Hill DRA. Goniomonas: ribosomal RNA sequences indicate that this phagotrophic flagellate is a close relative of the host component of cryptomonads. Eur J Phycol. 1994
- Maddison WP. Gene trees in species trees. Syst Biol. 1997
- Stiller JW. Plastid endosymbiosis, genome evolution and the origin of green plants. Trends Plant Sci. 2007
- Steiner JM, Yusa F, Pompe JA, Loffelhardt W. Homologous protein import machineries in chloroplasts and cyanelles. Plant J. 2005
- Stoebe B, Kowallik KV. Gene-cluster analysis in chloroplast genomics. Trends Genet. 1999
- Durnford DG, Deane JA, Tan S, McFadden GI, Gantt E, et al. A phylogenetic assessment of the eukaryotic light-harvesting antenna proteins, with implications for plastid evolution. J Mol Evol. 1999
- Rissler HM, Durnford DG. Isolation of a novel carotenoid-rich protein in Cyanophora paradoxa that is immunologically related to the light-harvesting complexes of photosynthetic eukaryotes. Plant Cell Physiol. 2005
- Stoebe B, Martin W, Kowallik KV. Distribution and nomenclature of protein-coding genes in 12 sequenced chloroplast genomes. Plant Mol Biol Rep. 1998
- Loffelhardt W, Bohnert HJ, Bryant DA. The complete sequence of the Cyanophora paradoxa cyanelle genome (Glaucocystophyceae). Plant Syst Evol. 1997
- O’Kelly C. Relationships of eukaryotic algal groups to other protists. In: Berner T, editor. Ultrastructure of microalgae. Boca Raton, FL: CRC Press; 1993
- Stiller JW, Harrell L. The largest subunit of RNA polymerase II from the Glaucocystophyta: functional constraint and short-branch exclusion in deep eukaryotic phylogeny. BMC Evol Biol. 2005
- Baldauf SL, Roger AJ, Wenk-Siefert I, Doolittle WF. A kingdom-level phylogeny of eukaryotes based on combined protein data. Science. 2000
- Burger G, Saint-Louis D, Gray MW, Lang BF. Complete sequence of the mitochondrial DNA of the red alga Porphyra purpurea: cyanobacterial introns and shared ancestry of red and green algae. Plant Cell. 1999
- Secq MPO, Goer SL, Stam WT, Olsen JL. Complete mitochondrial genomes of the three brown algae (Heterokonta: Phaeophyceae) Dictyota dichotoma, Fucus vesiculosus and Desmarestia viridis. Curr Genet. 2006
- Kim E, Lane CE, Curtis BA, Kozera C, Bowman S, et al. Complete sequence and analysis of the mitochondrial genome of Hemiselmis andersenii CCMP644 (Cryptophyceae). BMC Genomics. 2008
- Gibbs SP. The Chloroplasts of some algal groups may have evolved from endosymbiotic eukaryotic algae. Ann N Y Acad Sci. 1981
- Rumpho ME, Summer EJ, Manhart JR. Solar-powered sea slugs. Mollusc/algal chloroplast symbiosis. Plant Physiol. 2000
- Leander BS, Keeling PJ. Morphostasis in alveolate evolution. Trends Ecol Evol. 2003
- Moriya M, Nakayama T, Inouye I. A new class of the stramenopiles, Placididea classis nova: description of Placidia cafeteriopsis gen. et sp nov. Protist. 2002
- Kim E, Archibald JM. Diversity and evolution of plastids and their genomes. In: Sandelius AS, Aronsson H, editors. The Chloroplast: Interactions with the environment. Heidelberg: Springer; 2008
- Harper JT, Keeling PJ. Nucleus-encoded, plastid-targeted glyceraldehyde-3-phosphate dehydrogenase (GAPDH) indicates a single origin for chromalveolate plastids. Mol Biol Evol. 2003
- Takishita K, Ishida KI, Maruyama T. Phylogeny of nuclear-encoded plastid-targeted GAPDH gene supports separate origins for the peridinin- and the fucoxanthin derivative-containing plastids of dinoflagellates. Protist. 2004
- Takishita K, Kawachi M, Noel MH, Matsumoto T, Kakizoe N, et al. Origins of plastids and glyceraldehyde-3-phosphate dehydrogenase genes in the green-colored dinoflagellate Lepidodinium chlorophorum. Gene. 2008
- Martin W, Rujan T, Richly E, Hansen A, Cornelsen S, et al. Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proc Natl Acad Sci U S A. 2002
- Ohta N, Matsuzaki M, Misumi O, Miyagishima S, Nozaki H, et al. Complete sequence and analysis of the plastid genome of the unicellular red alga Cyanidioschyzon merolae. DNA Res. 2003
- Bachvaroff TR, Puerta MVS, Delwiche CF. Chlorophyll c-containing plastid relationships based on analyses of a multigene data set with all four chromalveolate lineages. Mol Biol Evol. 2005
- Bodyl A, Moszczynski K. Did the peridinin plastid evolve through tertiary endosymbiosis? A hypothesis. Eur J Phycol. 2006
- Lee RE, Kugrens P. Katablepharis ovalis, a colorless flagellate with interesting cytological characteristics. J Phycol. 1991
- Lee RE, Kugrens P, Mylnikov AP. The structure of the flagellar apparatus of two strains of Katablepharis (Cryptophyceae). Br Phycol J. 1992
- Clay B, Kugrens P. Systematics of the enigmatic kathablepharids, including EM characterization of the type species, Kathablepharis phoenikoston, and new observations on K. remigera com. nov. Protist. 1999
- Domozych DS, Wells B, Shaw PJ. Scale biogenesis in the green alga, Mesostigma viride. Protoplasma. 1992
- Domozych DS, Stewart KD, Mattox KR. Development of the cell wall in Tetraselmis: role of the Golgi apparatus and extracellular wall assembly. J Cell Sci. 1981
- Gupta RS. Protein phylogenies and signature sequences: a reappraisal of evolutionary relationships among archaebacteria, eubacteria, and eukaryotes. Microbiol Mol Biol Rev. 1998
- Boorstein WR, Ziegelhoffer T, Craig EA. Molecular evolution of the HSP70 multigene family. J Mol Evol. 1994
- Maddison DR, Maddison WP. MacClade 4: analysis of phylogeny and character evolution. Sunderland, MA: Sinauer Associates Inc; 2001
- Inagaki Y, Simpson AGB, Dacks JB, Roger AJ. Phylogenetic artifacts can be caused by leucine, serine, and arginine codon usage heterogeneity: dinoflagellate plastid origins as a case study. Syst Biol. 2004
- Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006
- Lartillot N, Brinkmann H, Philippe H. Suppression of long-branch attraction artefacts in the animal phylogeny using a site-heterogeneous model. BMC Evol Biol. 2007
- Abascal F, Zardoya R, Posada D. ProtTest: selection of best-fit models of protein evolution. Bioinformatics. 2005
- Schmidt HA, Strimmer K, Vingron M, von Haeseler A. TREE-PUZZLE: maximum likelihood phylogenetic analysis using quartets and parallel computing. Bioinformatics. 2002
- Desper R, Gascuel O. Fast and accurate phylogeny reconstruction algorithms based on the minimum-evolution principle. J Comput Biol. 2002
- Felsenstein J. Seattle: Department of Genome Sciences, University of Washington; 2005
Источник: wiki2.red
Прокариоты
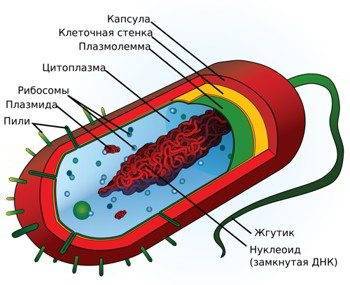
Прокариоты — организмы, состоящие из клеток, которые не имеют клеточного ядра или любых мембранных органелл. Это означает, что генетический материал ДНК у прокариот не связан в ядре. Кроме того, ДНК прокариот менее структурировано, чем у эукариот. В прокариот, ДНК одноконтурный. ДНК эукариот организовано в хромосомы. Большинство прокариот состоят только из одной клетки (одноклеточные), но есть несколько и многоклеточных. Ученые разделяют прокариот на две группы: бактерии и археи.
Типичная клетка прокариота содержит следующие части:
- клеточные стенки;
- плазматическая (клеточная) мембрана;
- цитоплазма;
- рибосомы;
- жгутики и пили;
- нуклеоид;
- плазмиды;
Источник: natworld.info