Эволюция членистоногих
Международная группа палеонтологов выпустила свежий обзор данных, касающихся появления в палеонтологической летописи первых представителей самого крупного типа животных — членистоногих. Сразу несколько дисциплин, включая молекулярную филогенетику и тафономию (науку о закономерностях захоронения ископаемых остатков), приводят к выводу, что реальным временем возникновения членистоногих был рубеж эдиакарского и кембрийского периодов, когда произошел так называемый «кембрийский взрыв». Авторы обзора отвергают конкурирующую гипотезу «долгой скрытой докембрийской эволюции», согласно которой главные ветви животных, в том числе и членистоногих, возникли гораздо раньше.
Хозяева Земли
Говорят, что знаменитый английский биолог Джон Холдейн (John Burdon Sanderson Haldane) однажды оказался в компании богословов, и они задали ему вопрос: какие черты Творца обнаруживаются при изучении природы творения? Холдейн, известный своими материалистическими взглядами, проворчал: «Неумеренная увлеченность жуками» (“An inordinate fondness for beetles”; цитируется по статье: G. E. Hutchinson, 1959.
mage to Santa Rosalia or why are there so many kinds of animals?). Этот шуточный ответ был, однако, основан на биологических фактах. Широко известно, что жуки, или жесткокрылые, — это самый многочисленный отряд во всем животном царстве. Если, например, сравнить жуков и млекопитающих, окажется, что число известных на данный момент видов жуков превосходит число видов млекопитающих в 72 раза. И это несмотря на то, что млекопитающие — не отряд, а класс, то есть группа значительно более высокого уровня.
Сказанное о жуках можно распространить и на их эволюционных родственников — от близких до дальних. Жуки относятся к насекомым, которые, в свою очередь, входят в тип членистоногих. Разнообразие этого типа поистине грандиозно. По современным подсчетам, больше 80% всех видов многоклеточных животных — это членистоногие. Другой настолько успешной эволюционной ветви просто нет в природе. Замечательная книга Эдварда Уилсона (Edward Osborne Wilson), значительная часть которой рассказывает о возникновении социальных систем у членистоногих, недаром называется «Хозяева Земли».
Неудивительно, что эволюция членистоногих занимает многих исследователей. Дело облегчается тем, что палеонтологическая летопись членистоногих сравнительно богата. Более того, ее «чтение» — работа достаточно благодарная.
гласно принципу актуализма, ключом к пониманию прошлого служит настоящее. В частности, любые выводы, касающиеся ископаемых животных, должны быть в конечном счете обязательно основаны на данных о животных современных — иначе исследователю будет просто не на что опереться. В случае с членистоногими этот принцип работает прекрасно, потому что в распоряжение ученых попадает огромное количество как современных, так и ископаемых форм, очень разнообразных, но в то же время устроенных более-менее по одному плану. Это — идеальная ситуация для палеонтолога, и естественно, что исследователи ископаемых членистоногих пользуются ей сполна (см., например: Палеоэнтомология в России).
Членистоногие — группа организмов, очень подходящая для проверки всевозможных гипотез, касающихся хода эволюции. Например, действительно ли большинство современных крупных групп животных возникло во время кембрийского взрыва или их внезапное появление в начале кембрийского периода связано с образованием твердых скелетов, а истинные эволюционные «корни» лежат гораздо глубже? Попытки выяснить это, основываясь на материале по членистоногим, уже предпринимались (см. Членистоногие подтверждают реальность кембрийского взрыва, «Элементы», 17.11.2013). Но эволюция членистоногих, особенно ранняя, крайне интересна и сама по себе — как история, героями которой являются многие совершенно необычные с нашей современной точки зрения живые существа. Палеонтология XXI века успела сделать много открытий в этой области. Впрочем, и нерешенных вопросов тут, как всегда, еще хватает.
Усики, жвалы и хелицеры
По современным данным, тип членистоногих состоит из двух крупных эволюционных ветвей.
Одна ветвь — хелицеровые (Chelicerata), куда относятся паукообразные, мечехвосты и морские пауки. У них нет усиков, зато есть хелицеры — околоротовые придатки, оканчивающиеся клешнями, крючками или стилетами. Большинство хелицеровых, во всяком случае крупных, — хищники, потому что для других способов питания их ротовой аппарат подходит мало.
Вторая ветвь членистоногих называется жвалоносными (Mandibulata). В их ротовой аппарат входят челюсти — в том числе мандибулы, которые иначе называются жвалами, — представляющие собой не что иное, как сильно видоизмененные ходильные ноги. Этим жвалы принципиально отличаются от хелицер, которые не имеют с ногами ничего общего. Ротовой аппарат жвалоносных, оснащенный набором челюстей, позволяет осваивать предельно разнообразные способы питания, что мы и видим на примере членов этой группы — многоножек, ракообразных и особенно насекомых. Кроме того, для жвалоносных очень характерны усики, или антенны, — одна или две пары. Как уже упоминалось, у хелицеровых усиков нет.
Строго говоря, тип, состоящий из жвалоносных и хелицеровых, называется Euarthropoda (эуартроподы, «настоящие членистоногие»). Это название, более точное, чем просто «членистоногие», предложил в 1904 году знаменитый английский зоолог Рэй Ланкестер (Sir Edwin Ray Lankester). Среди признаков эуартропод — четкое деление тела на отделы (тагмы) и присутствие членистых конечностей, внутри которых есть суставы.
Животные, у которых членистых конечностей нет и никогда не было, не относятся к эуартроподам, даже если по многим другим признакам они к ним близки. Существует две группы таких животных: онихофоры — наземные существа, которых иногда называют «бархатными червями» (velvet worms), — и тихоходки, мельчайшие водяные жители (см., например: Укорочение тела тихоходок связано с потерей Hox-генов, «Элементы», 04.03.2016). И у онихофор, и у тихоходок тело сегментировано почти так же, как у членистоногих, но конечности нечленистые. Поэтому диагнозу типа Euarthropoda, равно как и буквальному значению слова «членистоногие», они не соответствуют.
Тем не менее онихофоры и тихоходки, безусловно, являются близкими родственниками эуартропод. В конце XX века эуартропод, онихофор и тихоходок формально объединили в надтип Panarthropoda (букв. «все членистоногие»). Панартроподы — огромная эволюционная ветвь, которая, в свою очередь, входит в группу линяющих животных (Ecdysozoa; см., например: Китайские палеонтологи нашли древнейшего головохоботного червя, «Элементы», 04.06.2014). Таково положение членистоногих в системе животного мира.
«Стволовые эуартроподы»
До сих пор, говоря о членистоногих и их родственниках, мы упоминали только современные группы. Однако для полного понимания эволюционных событий, сформировавших самый многочисленный тип животного царства, просто необходимо учитывать палеонтологическую летопись, и прежде всего — летопись кембрийского периода, когда сформировались почти все крупные эволюционные ветви животных. В кембрии встречаются интереснейшие живые существа, совершенно не похожие на своих современных родственников (многих из них нельзя отнести ни к какому современному типу — по крайней мере, без явных натяжек). Трудность в том, что «мягкотелые» существа, у которых нет минеральных скелетов или раковин, сохраняются в ископаемом состоянии относительно редко, и к членистоногим это, увы, тоже относится. Для их захоронения нужны лагерштетты — глинистые осадочные породы, образующиеся в малокислородных условиях (там, где кислорода много, в грунт проникают роющие животные, которые сразу нарушают структуру формирующегося осадка; см. А. Ю. Журавлев, 2014. Ранняя история Metazoa — взгляд палеонтолога). Особой известностью в качестве источника кембрийских ископаемых, заключенных в лагерштеттах, пользуются канадские сланцы Бёрджесс (Burgess Shale), где раскопки ведутся уже больше ста лет. Фауна сланцев Бёрджесс настолько знаменита, что местонахождения с подобным типом сохранности обозначаются в научных статьях аббреавиатурой BST — Burgess Shale-type, «тип сланцев Бёрджесс».
леонтология в наши дни развивается быстро, и местонахождений BST известно много, причем в некоторых из них степень сохранности просто поразительна. Это позволило неплохо изучить древнейших родственников членистоногих и, что называется, пролить свет на их происхождение. Правда, открывшуюся картину никак не назовешь простой. Но тем она интереснее.
Бросим взгляд на эволюционное древо (рис. 2). По молекулярным данным, которые в данном случае неплохо согласуются с палеонтологическими, ближайшие «внешние» родственники эуартропод, дожившие до наших дней, — это онихофоры. В палеонтологической летописи онихофоры — или, во всяком случае, близкие к ним формы — тоже присутствуют. Правда, в кембрийском периоде они были еще не наземными, а морскими (см. Aysheaia). Это гусеницеобразные существа, которые передвигались по дну на множестве коротких ножек, оканчивающихся коготками. Вполне вероятно, что общие предки эуартропод и онихофор были на них похожи.
Однако онихофоры вряд ли являются непосредственными предками эуартропод (во всяком случае, эуартропод в строгом и традиционном смысле этого слова). Вот тут-то и начинается самое интересное. На реальном эволюционном древе между ветвью онихофор и ветвью эуартропод находится целая совокупность весьма оригинальных полностью вымерших ветвей, члены которых обладали отдельными «эуартроподными» признаками, но еще не набрали их полного комплекса, какой мы видим у паука, сороконожки или мухи.
и ветви можно было бы назвать переходными, но надо помнить, что для своей среды и своего времени каждое относящееся к ним животное было вполне завершенным организмом, отлично вписанным в определенную экологическую нишу (иначе бы оно не попало в палеонтологическую летопись). Хотя, конечно, с нашей современной точки зрения многие члены этих «переходных» ветвей выглядят крайне странно.
В современной научной литературе упомянутую совокупность эволюционных ветвей принято называть «стволовой группой эуартропод», или просто «стволовыми эуартроподами» (stem-Euarthropoda). Честно говоря, такое обозначение может изрядно запутать: ведь в том-то и дело, что животные, на которых оно распространяется, к эуартроподам в узком смысле этого слова не относятся (см. J. Ortega‐Hernandez, 2014. Making sense of ‘lower’and ‘upper’ stem‐group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848). Однако — тут нам поневоле придется употребить предложение, насыщенное терминами, — кладистическая систематика, господствующая в современной биологии почти безраздельно, категорически запрещает вводить парафилетические таксоны. Если говорить попросту, это означает, что группа организмов, охватывающая не единственную эволюционную ветвь, а целый эволюционный уровень, не может считаться единицей классификации и иметь собственное научное название (в частности, латинское).
скольку в данном случае такая группа налицо и как-то называть ее всё же надо, исследователи используют временное обозначение, которое было сочтено корректным по формальным соображениям. Мы тоже — куда деваться — будем этим обозначением пользоваться, оставляя его в кавычках и помня, что с точки зрения классической зоологии «стволовые эуартроподы» — это не эуартроподы.
Положение «стволовых эуартропод» на эволюционном древе показано на рис. 2. Если на время оставить в стороне строгую терминологию, можно сказать, что «стволовые эуартроподы» — это эволюционный уровень, охватывающий часть древа выше онихофор и ниже эуартропод в узком смысле. Что же на этом эволюционном уровне происходило?
А происходило там много интересного. «Стволовые эуартроподы» могут поразить чье угодно воображение — настолько необычен и многообразен их облик (см. рис. 1 и 3). Их членистое туловище часто украшали ряды жаброподобных, крылоподобных или лезвиеподобных выростов, способных служить своего рода плавниками. Появлялись хитиновые головные щиты, которые в некоторых группах становились двустворчатыми. Ходильные конечности менялись во всем диапазоне от примитивных «сосочков» (или их отсутствия) до сложно устроенных длинных ног, членистых, а иногда и двуветвистых, как у ракообразных.
вероятным разнообразием отличались околоротовые придатки: членистые или нечленистые, зачаточные или очень мощные, короткие или длинные, хватательные или фильтрующие, иногда ветвящиеся, а иногда с клешнями, крючками, шипами или щупальцами. Наконец, у ряда форм возникли огромные сложные фасеточные глаза, примерно такие же, как у современных насекомых, и иногда сидящие на стебельках. И все эти признаки вступали между собой во множество сочетаний, часто неожиданных для современных зоологов.
Эволюционный процесс, охвативший «стволовых эуартропод», хорошо описывается введенным Александром Георгиевичем Пономаренко термином «артроподизация» (А. Г. Пономаренко, 2004. Артроподизация и ее экологические последствия). Это означает, что признаки членистоногих возникали параллельно во многих (хотя и родственных) эволюционных ветвях, подчиняясь, с одной стороны, общей наследственности, а с другой — общим требованиям окружающей среды. При этом одни и те же адаптации зачастую реализовались в разных эволюционных линиях, во-первых, в разной последовательности, а во-вторых — слегка различными способами. Именно это и дает возможность отличить постепенную параллельную эволюцию от гипотетического явления нового типа сразу «во всеоружии», — подобно Афине, которая вышла в полном вооружении из головы Зевса.
Например, некоторые исследователи считают, что как минимум в двух разных ветвях «стволовых эуартропод» членистые хватательные околоротовые придатки, функционально сходные между собой, на самом деле принадлежат к разным сегментам головы — а значит, развились независимо и никак не могут быть гомологами друг друга (D. A. Legg, J. Vannier, 2013.
e affinities of the cosmopolitan arthropod Isoxys and its implications for the origin of arthropods). Это очень правдоподобно: именно так и должен был бы выглядеть процесс артроподизации, если исходить из теоретических представлений, подтвержденных данными по другим группам организмов, которые претерпели аналогичную эволюцию (классический пример такой группы — зверообразные, предки млекопитающих).
«Кроновые эуартроподы»
Для полноты картины следует добавить, что эуартроподы в классическом «рэй-ланкастеровском» смысле более или менее соответствуют современному понятию «кроновая группа эуартропод», или просто «кроновые эуартроподы» (crown-Euarthropoda). Кроновой группой (crown group) в современной систематике называется часть эволюционного древа, охватывающая ближайшего общего предка всех доживших до наших дней членов данной группы вместе со всеми его потомками (неважно, ныне живущими или вымершими; см. Древнейшие предки кольчатых червей могли быть похожи на брахиопод, «Элементы», 26.02.2016). А стволовой группой (stem group) называется та часть древа, ветви которой до современности не дотягиваются (см. рис. 2).
Есть одна группа членистоногих, которую мы еще ни разу не упомянули. Это трилобитообразные (Trilobitomorpha). Трилобиты — широко известные палеозойские животные, в целом похожие внешне на мокриц, но обладавшие огромным разнообразием как облика, так и образа жизни (см. Р. Форти, 2014. Трилобиты. Свидетели эволюции). У них есть четкое деление тела на тагмы, а также настоящие антенны и двуветвистые членистые конечности. Все это — эуартроподные признаки. Современные биологи склонны относить трилобитов к кроновой группе эуартропод, несмотря на то, что они полностью вымерли. Правда, к кому они ближе внутри кроновой группы — к хелицеровым или к жвалоносным, — пока непонятно. Молекулярные методы, которые могли бы однозначно решить этот вопрос, в данном случае неприменимы: ведь последние трилобиты исчезли в пермском периоде, и никакой ДНК от них не осталось.
Первые членистоногие
Теперь мы узнали вполне достаточно, чтобы оценить по достоинству крайне интересный обзор, опубликованный весной 2018 года коллективом из четырех довольно молодых, но уже известных палеонтологов (один соавтор родом из Канады, трое других — из Британии). Все они — специалисты по ископаемым членистоногим, их родственникам и их происхождению. Главный вопрос, который интересовал авторов: что могут нам сообщить эуартроподы (как «кроновые», так и «стволовые»), если рассматривать их в качестве свидетелей кембрийского взрыва?
Чтобы вникнуть в суть дела, нам понадобится немного цифр. По наиболее свежим датировкам, кембрийский период начался 541 миллион лет назад. Остатки членистоногих появляются в кембрийских отложениях отнюдь не сразу. Первые найденные в ископаемом состоянии эуартроподы «кроновой группы» — это трилобиты, самые ранние остатки которых имеют возраст 521 миллион лет. Причем обнаруживаются они почти одновременно в нескольких точках Земли: едва появившись, трилобиты быстро приобрели глобальное распространение (очевидно, не в последнюю очередь потому, что в жизненном цикле у них были планктонные личинки, которых могут свободно переносить морские течения). Что касается «стволовых эуартропод», то самый древний их представитель — существо, относящееся к группе аномалокаридид, от которого найден характерный околоротовой придаток (A. C. Daley, D. A. Legg, 2015. A morphological and taxonomic appraisal of the oldest anomalocaridid from the Lower Cambrian of Poland). Возраст этой находки пока определен лишь приближенно: 519–520 миллионов лет. Чуть позже, 518 миллионов лет назад, на сцену выходит так называемая Чэнцзянская биота, где «стволовые эуартроподы» представлены во всем великолепии. И наконец, 514 миллионов лет назад в летописи появляются микроскопические остатки первых ракообразных. Еще через несколько миллионов лет ракообразных становится много — фауна приближается к современной, кембрийский взрыв остается позади.
Что дают нам все эти даты? Прежде всего, налицо парадоксальный факт: «стволовые эуартроподы» появляются в палеонтологической летописи позже, чем «кроновые». Грубо говоря, предки появляются в летописи позже потомков (пусть и совсем ненамного позже). Что это значит? Никто, конечно, не сомневается, что «кроновые эуартроподы» произошли от кого-то из «стволовых». Но не менее несомненно и то, что какое-то время эти группы существовали вместе. Скорее всего, дело в том, что «стволовые эуартроподы» в принципе хуже представлены в летописи: их кутикула была существенно менее твердой, чем у трилобитов и ракообразных, а потому их тела (или части тел) с меньшей вероятностью сохранялись в донных осадках. Тут стоит вспомнить, что у онихофор, которых не зря называют «бархатными червями», кутикула до сих пор мягкая, бархатистая, растягивающаяся, — совсем не такая, как у ракообразных или насекомых.
Есть, правда, и другая возможность. Что, если трилобиты — это не «кроновые эуартроподы», а одна из групп «стволовых», вышедшая на «истинно эуартроподный» эволюционный уровень совершенно самостоятельно? Это теоретически не исключено, но нуждается в серьезных доказательствах, которых пока нет.
Как видим, в первые двадцать миллионов лет кембрия ни «стволовые», ни «кроновые эуартроподы» не обнаруживаются (во всяком случае, пока — не стоит забывать, что палеонтология продолжает стремительно развиваться и новые находки могут возникнуть в любой момент). Значит ли это, что они появляются в летописи внезапно? Нет. Как обычно и бывает в таких случаях, находкам захороненных тел (или отпечатков тел) предшествуют находки ископаемых следов, предположительно оставленных теми же организмами. Это тоже достойный предмет изучения. Существует целая наука об ископаемых следах — палеоихнология. Итак, в отложениях начала кембрия присутствуют довольно многочисленные ископаемые следовые дорожки, часть из которых, видимо, принадлежит «стволовым эуартроподам», а часть «кроновым». Возраст древнейших из них — примерно 537 миллионов лет.
Ну, а что было еще раньше? Весь отрезок истории Земли, предшествующий началу кембрийского периода, принято называть докембрием. Непосредственно с кембрием снизу — со стороны более древних времен — граничит эдиакарский период, относящийся к протерозойской эре. Живая природа эдиакария уникальна. Она резко отличается от живой природы кембрия. Следы членистоногих — «стволовых», «кроновых» и вообще каких бы то ни было — в эдиакарии, по словам некоторых авторов, «поразительным образом отсутствуют» (M. G. Mangano, L. A. Buatois, 2014. Decoupling of body-plan diversification and ecological structuring during the Ediacaran–Cambrian transition: evolutionary and geobiological feedbacks). Впрочем, это не так уж и поразительно, если исходить из вполне вероятной гипотезы, что членистоногих тогда просто не было.
Загадочная сприггина
Однако голым отрицанием тут не отделаешься. Существует такая вещь, как тафономия — наука о закономерностях захоронения ископаемых остатков, которой дал название великий Иван Антонович Ефремов. Каждый палеонтолог прекрасно знает, что отсутствие той или иной группы организмов в палеонтологической летописи может объясняться эволюционными причинами (эта группа либо еще не возникла, либо уже вымерла), а может и тафономическими (эта группа существовала, но жила в таких условиях, где захоронение остатков ее представителей было крайне маловероятно). Выбор между этими вариантами часто приводит к серьезным дискуссиям. Здесь перед нами именно такой случай. Что, если отсутствие в летописи докембрийских членистоногих — это на самом-то деле тафономический артефакт? Такое возможно, если условия захоронения в эдиакарии были принципиально иными, чем в кембрии.
Понимая эту возможность, авторы обзора дотошно проанализировали данные о том, насколько эдиакарские местонахождения древних организмов (а их известно не так уж мало) отличаются от кембрийских. Был сделан вывод, что наиболее важные с точки зрения нашей проблемы типы сохранности являются для эдиакария и кембрия фактически общими. Речь идет не только о классических лагерштеттах «типа Бёрджесс», но и о захоронениях фосфатизированных микрофоссилий «типа Эрстен» (Orsten-type), которые тоже дают очень интересные находки. Эти типы местонахождений отнюдь не ограничиваются кембрием: в эдиакарии они тоже распространены, и в них отлично сохраняются тогдашние организмы, причем весьма разнообразные. Иное дело, что сплошь и рядом такие находки бывает сложно интерпретировать: увы, эдиакарские живые существа слишком необычны по современным меркам, и поминавшийся нами принцип актуализма начинает тут давать сбои (см., например: Ископаемые эмбрионы из Доушаньтуо, вероятно, не эмбрионы, «Элементы», 09.01.2012). Но уж если бы там были членистоногие — то они сохранились бы наверняка, по крайней мере, хоть в каком-нибудь количестве.
Более того, можно утверждать, что по совокупности геохимических факторов, влиявших на захоронение ископаемых, кембрий был ближе к эдиакарию, чем к более поздним эпохам. Именно к концу кембрия условия в морях сильно изменились — в первую очередь из-за деятельности животных, которые связывали растворенные соли в материале своих раковин, меняя химический состав морской воды, а заодно активно фильтровали эту воду, делая ее более прозрачной и тем самым улучшая условия для водорослей, насыщающих океан кислородом. Кроме того, животные вовсю рыли норы в морском дне, перемешивая грунт и в результате обогащая кислородом его тоже. Все это, вместе взятое, сокращало возможности для образования стабильных бескислородных минерализованных осадков — а как раз из таких осадков и получаются лагерштетты. Неудивительно, что после кембрия их стало гораздо меньше.
Итак, объяснить отсутствие докембрийских членистоногих чисто тафономическими причинами невозможно. Правда, известно несколько эдиакарских ископаемых, которых разные авторы объявляли членистоногими (или членами вымерших типов, близких к ним). Но эта гипотеза ни в одном случае не подтверждена сколько-нибудь надежно.
Типичным примером такого ископаемого можно назвать сприггину (Spriggina) — эдиакарское существо с вытянутым сегментированным телом и неким подобием головного щита. Ее не раз описывали то как кольчатого червя, то как членистоногое (см., например: M. McMenamin, 2003. Spriggina is a trilobitoid ecdysozoan). Однако, во-первых, у сприггины не найдено никаких усиков, ножек, челюстей и вообще конечностей. А во-вторых, обычную для животных двустороннюю симметрию у нее заменяет симметрия скользящего отражения, когда правая и левая стороны тела сдвинуты относительно друг друга примерно на половину сегмента. Уже одно это означает, что если сприггина и была многоклеточным животным, то от плана строения членистоногих ее план строения отличался радикально. Принадлежать к этому типу она никак не могла. Примерно так же обстоит дело и со всеми остальными эдиакарскими находками.
История со сприггиной хорошо показывает, насколько усложняется работа палеонтологов при погружении в древнейшие времена, где уже не действуют сравнения с современной флорой и фауной. Ее предполагаемая более поздняя родственница — метасприггина (Metaspriggina) — в итоге вообще оказалась хордовым животным (P. Janvier, 2015. Facts and fancies about early fossil chordates and vertebrates). Ну, а сама сприггина пока что так и остается загадкой.
Источник: elementy.ru
Нет сомнения, что Arthropoda произошли от червеподобного предка, близкого по общему строению к Annelida, или кольчатым червям. Тело этого предка (рис. 2.1) состояло из серии одинаковых сегментов, каждый из которых представлял собой замкнутое кольцо. Голова имела простое строение и, по-видимому, несла чувствительные щетинки. Ротовое отверстие было расположено на вентральной стороне тела между головой и первым сегментом тела. Из-за своего положения впереди ротового, или стомодеального, отверстия головной отдел этих животных называют простомиумом.
Как отмечал Мэнтон (Manton, 1973), этот червеподобный предок членистоногих отличался от современных Annelida по ряду важных черт в строении тела. У Annelida каждый сегмент тела имеет свою полость, или целом, причём целомы разных сегментов разделены перегородками — септами. В каждом сегменте имеются выделительные органы (парные нефридии), открывающиеся наружу выводными протоками. Ноги, если они имеются, также расположены посегментно и характеризуются примитивным строением. У гипотетических членов ряда, ведущего к Arthropoda, отсутствовали межсегментные перегородки и полость тела простиралась по всей его длине. Выделительная система представляла собой один орган, обслуживавший весь организм животного. Ноги, если они имелись, были с более специализированной, чем у Annelida, мускулатурой. На рис. 2.1 показаны ступени эволюционного развития гипотетического предка от простой преартроподной организации через генерализованный артроподный тип строения к насекомым.
Первая важная ступень состояла в образовании на вентральной стороне каждого сегмента пары конечностей, или ног, обеспечивавших передвижение животного (рис. 2.1). Очевидно, последний сегмент, перипрокт, на котором располагалось анальное отверстие, никогда не имел конечностей.
Следующим вероятным изменением было слияние конечностей первого сегмента и образование лабрума — вентрального придатка, функция которого состояла в направлении пищи к ротовому отверстию. Конечности второго сегмента переместились на дорсальную сторону и превратились в антенны. Конечности третьего сегмента, имеющиеся на ранних эмбриональных стадиях насекомых, исчезают как самостоятельные структуры на более поздних стадиях развития. Один за другим эти первые три сегмента, носящие соответственно названия лабральный, антеннальный и интеркалярный, сливаются с анцестральным простомиумом, образуя сложную голову, составленную из исходных шести сегментов. Ганглии этих сегментов сливаются с первичным мозгом, или архицеребрумом, и образуется сложный головной мозг членистоногих.
Эти изменения в строении головы и головного мозга сопровождались другим эволюционным преобразованием, важным для питания животных. Ротовое отверстие, расположенное вначале позади лабрального сегмента, затем переместилось в положение позади антеннального сегмента и, наконец, оказалось позади интеркалярного сегмента. Несомненно, что на этом доартроподном этапе эволюции конечности нескольких следующих сегментов, пока лишь слегка модифицированные, служили для направления пищи к ротовому отверстию. Ротовое отверстие, смещённое кзади, вместе с лабрумом, также переместившимся назад, образовало нечто вроде кармана, где пища могла накапливаться до того, как животное проглатывало её. Поскольку термин «простомиум» не соответствовал более тому строению, которое приобрела первичная голова после перемещения ротового отверстия назад, она получила название акрон.
Перемещение ротового отверстия из положения между акроном и сегментом 1 в положение между сегментами 3 и 4 требует некоторого разъяснения. На ранних стадиях эмбрионального развития структуры вентральной части тела формировались до того, как сегменты приобретали свою окончательную форму-форму колец. Предшественниками будущих нервных центров, или ганглиев, были расположенные в каждом сегменте парные, но не связанные между собой латеральные массы нервной ткани, называемые нейромерами (нервными телами). На ранних стадиях эмбриогенеза эти отдельные нейромеры, по-видимому, продвигались вперёд по обеим сторонам от ротового отверстия и затем сливались с первичным мозгом.
Примерно на этой стадии эволюции членистоногих произошло превращение конечностей сегмента 4 в хватательные и режущие придатки, называемые мандибулами. Однако нейромеры этого сегмента не переместились вперёд и ротовое отверстие осталось между сегментами 3 и 4.
Эта стадия эволюции представлена тремя группами Onychophora, Pentastomida и Tardigrada, особенно чётко характеризующие её черты строения можно наблюдать у современных представителей Onychophora (Peripatus и близкие к нему формы, рис. 2.2.). Онихофоры имеют мягкое тело с посегментно расположенными парными конечностями, хотя внешние признаки сегментации чётко не выражены. Ноги онихофор, называемые лобоподиями, не сегментированы, но имеют специальные мышцы, прикреплённые к неподвижным основаниям (Manton, 1973). У них также имеется пара придатков, сходных по своему виду и положению с мандибулами членистоногих, однако гомология их пока не доказана. Даже если это и так, онихофор всё-таки следует рассматривать как отдельную чёткую ветвь этого направления эволюции.
Представители двух других групп, Tardigrada (рис. 2.3.) и Pentastomida (рис. 2.4.), претерпели столь сильную редукцию организации, что их родственные связи требуют дальнейшего тщательного изучения. Tardigrada (тихоходки, или водяные медведи; рис. 2.3.) — это крошечные животные, живущие во влажном мху, а также в пресной и солёной воде. Их тело, никогда не превышающее в длину 11 мм, имеет 4 пары бугорковидных ног, заканчивающихся коготками. На голове нет ни чётко различимых ротовых придатков, ни каких-либо других структур. Pentastomida (язычковые, или пятиустки; рис. 2.4.) — небольшая группа животных с червеподобными взрослыми формами и крошечными четырёхногими неполовозрелыми стадиями, по общему виду похожими на клещей. Пятиустки являются внутренними паразитами ряда позвоночных.
Следующая стадия эволюции той ветви, которая ведёт к членистоногим, характеризуется некоторыми изменениями в сторону специализации.
- Ноги приобрели членистое строение. что способствовало развитию локомоторной функции.
- Некоторые участки покровов тела затвердели, и в этих местах образовались внутренние выросты (так называемые подемы и апофизы), служащие для прикрепления мышц, участвующих в движениях частей тела и конечностей (см. также рис. 5.1, В, Г).
- Конечности нескольких посторальных сегментов приобрели функцию — хотя поначалу и не выраженную структурно) перемещения пищи к ротовому отверстию.
В результате всех этих преобразований возник характерный представитель типа Arthropoda.
Самыми ранними из известных членистоногих животных были трилобиты — обитатели морей, составляющие подтип Trilobita (рис. 2.5, 2.6). Они появились в кембрийский период примерно 600 млн. лет назад и исчезли примерно 200 млн. лет назад. У этих животных была небольшая голова, ориентированная вперёд, с антеннами и лабрумом, а позади головы несколько коротких ног, возможно участвовавших в захвате пищи. Ноги и их основания не были, однако, слиты с головой.
Трилобиты дали начало развитию нескольких морфологически различающихся групп. Возможно, что две из них послужили исходными для образования двух крупных групп членистоногих — подтипов Chelicerata и Mandibulata.
У Chelicerata антенны отсутствуют и тело состоит из двух частей — головогруди (цефалоторакса), выполняющей функции питания и локомоции, и брюшка (абдомена), выполняющего функцию размножения. К Chelicerata относятся скорпионы, пауки, клещи и близкие к ним группы.
У Mandibulata (ствол, ведущий к насекомым) первые три посторальных сегмента и их конечности слились с головой, которая состоит, таким образом, из первичного акрона и 6 сегментов тела. Конечности трёх сегментов, присоединившихся к голове, превратились в разнообразные структуры, предназначенные для захватывания пищи и помещения её в рот. Три сегмента, несущие эти ротовые придатки, называют гнатальными сегментами.
Такое слияние, объединившее в одну функциональную единицу все начальные структуры, связанные в конечном счёте с питанием, представляет собой характерную черту всех Mandibulata за очень редкими исключениями.
Древние Mandibulata дали начало двум линиям, каждая из которых представлена сегодня большой многообразной группой; это — Crustacea, или ракообразные (крабы, креветки и близкие к ним формы), и группа Myriapoda-Insecta (многоножки и насекомые). Ракообразные — преимущественно обитатели водной среды, как, возможно, и древние Mandibulata . Исходными же формами ветви Myriapoda-Insecta, вполне вероятно были наземные животные. Они могли жить в листовой подстилке по берегам мелких затенённых водоёмов и речек подобно некоторым современным Isopoda-Crustacea). Некоторые зоологи считают, что ракообразные стоят особняком среди современных Mandibulata, а сходвство в строении их ротового аппарата является результатом конвергентной эволюции.
В ветви Myriapoda-Insecta произошло слияние третьей пары ротовых конечностей, или придатков (которые у ракообразных называют вторыми максиллами), образовался длинный поперечный вырост — лабиум. Этим завершилось формирование новой и функционально важной стркутуры — предротовой, или преоральной полости, передняя и задняя стенки которой образованы лабрумом и лабиумом, а боковые — мандибулами и максиллами. Наличие ротовых придатков и предротовой полости обеспечивало животным большие возможности для захватывания пищи. Штёрмер (Stormer) оценивает это морфологическое приспособление как важнейшую адаптацию к наземной жизни.
Наземный предок многоножек и насекомых дал начало трём ветвям, сохранившимся и в наше время: одна представлена классом Chilopoda (губоногие), другая-тремя классами Symphyla, Pauropoda и Diplopoda (симфилы, пауроподы и двупарноногие), и третья — классом Insecta (насекомые). В первых четырёх классах (многоножки) ходильные ноги имеются почти на всех туловищных сегментах.
Насекомые подверглись дальнейшей специализации. Первые три пары локомоторных конечностей увеличились в размерах, остальные редуцировались, а затем исчезли совсем или превратились в специальные образования (рис. 2.1, Е), не связанные с локомоцией. Таким образом, локомоторная функция сосредоточилась в трёх первых сегментах, расположенных позади головы, и сформировался хорошо выраженный отдел тела, носящий название грудь (торакс). Задняя часть тела, где расположена большая часть органов, называется брюшком (абдоменом). Задние абдоминальные конечности превратились в наружные половые придатки, предназначенные для спаривания и откладки яиц. У некоторых ракообразных грудь и брюшко также чётко разделены, но при этом грудь обычно состоит примерно из 8 сегментов.
Анализ преобразований, имевших место в эволюции членистоногих от их примитивного безногого предка, позволяет предположить что а) неспециализированные конечности появились сначала на всех посторальных сегментах и б) затем подверглись постепенным изменениям, приведшим к их морфологической и функциональной дифференцировке. У насекомых это привело к чёткому разделению тела на три отдела: голову, несущую сенсорные придатки и ротовые части, грудь, несущую три пары ног, и брюшко, внутри которого находятся жизненно важные органы, а снаружи — терминальные придатки, выполняющие функцию наружных половых органов.
Представление о родственных связях внутри типа членистоногих, принятое в настоящем руководстве, схематически изображено на рис. 2.5. Согласно этому представлению, все гексаподы произошли от одной ветви древних членистоногих и поэтому представляют собой моно-филетическую группу. Как считает Кристенсен (Kristensen, 1975), предлагаемая схема происхождения насекомых нуждается в наименьшем числе эволюционных параллелей для своего объяснения. Другие авторы придерживаются иной точки зрения и считают, что не все насекомые произошли от одной эволюционной ветви, то есть их происхождение носит полифилетический характер (Manton, 1949, 1964, 1977). Обе гипотезы о происхождении насекомых рассматриваются в книге под редакцией Гупты (Gupta, 1979).

Рисунок 2.1. Схематическое изображение гипотетических стадий (А-Е) в развитии различных отделов тела и его придатков — от червеподобного предка до насекомого
Рисунок 2.2. Древние и современные Onychophora
A. Peripatoides novae-zealandiae. Б. Реконструкция ископаемого Aysheaia pedunculata из среднего кембрия. В. Предполагаемый представитель онихофор Xenusion auerswaldae из докембрийского или раннекембрийского кварцита. Г, Д. Передний конец тела Peripatoides novae-zealandiae, вид с вентральной (Г) и латеральной (Д) сторон. Е. Его правая челюсть с мышцами: а-d — челюстные мышцы; 1 — антенна; 2 — оральная папилла; 3 — лабрум; 4 — челюсть; 5 — нога; 6 — складка вокруг ротового отверстия; 7 — глаз; 8 — аподема.
Рисунок 2.3. Тихоходка
Рисунок 2.4. Пятиустка
Слева — Porocephalus annulatus. А. Голова, вид с вентральной стороны. Б. Целое животное, вид с вентральной стороны. Справа — личинка Porocephalus proboscideus, вид с вентральной стороны: 1 — вбуравливающийся передний конец; 2 — первая пара склеротизированных отростков, видимая между вилками второй пары; 3 — вентральный нервный ганглий; 4 — пищеварительный канал; 5 — рот; 6 и 7 — железистые клетки.
Рисунок 2.5. Гипотетическое родословное древо членистоногих
Рисунок 2.6. Общий вид трилобита Triarthrus becki
А. Дорсальная (верхняя) поверхность карапакса (щита), состоящего из трёх долей: плевры (5), осевой доли (рахиса, 4) глабеллы (1) и свободных щёк (2), несущих глаза (3). Б. Вентральная (нижняя) поверхность; видны двуветвистые конечности (7 и 8), прикреплённые к осевой доле, и верхняя губа, или гипостом (6), покрывающая ротовое отверстие. Двуветвиртые ноги имеют двойную функцию; верхняя опушённая ветвь несёт жабры для дыхания и служит, кроме того, гребными лопастями, нижняя голая ветвь служит для ползания. Короткие передние придатки, возможно, участвуют в процессе питания.
Источник: www.entomologa.ru
Тип членистоногие наиболее богатый видами среди всех животных. Он насчитывает около 1,5 млн. видов, которые широко расселились по всему земному шару и на данный эволюционный период — господствуют, достигнув невиданного расцвета и отлично приспособившись к жизни вблизи людей.
Членистоногие — первичноротые трехслойные многоклеточные животные, произошедшие от многощетинковых кольчецов и включающие в себя многочисленные классы — ракообразные, паукообразные и насекомые. Давайте обозначим ароморфозы, которые помогли занять членистоногим их доминирующее положение, и общие черты строения.
По-другому членистоногих называют артроподы (от др.-греч. ἄρθρον — сустав и πούς, род. п. ποδός — нога), что еще раз подчеркивает их важнейшие ароморфозы, о которых вы скоро узнаете.
Ароморфозы членистоногих
- Членистые конечности
- Поперечно-полосатая мускулатура
- Слияние сегментов тела
- Наружный скелет — хитиновый покров
- Способность к полету у насекомых
Органы движения членистоногих имеют более сложное строение, через суставы соединяясь с телом животного. Конечности подобны многоколенным рычагам, к которым крепятся мышцы, приводящие их в движение. Выполняют самые разные функции: копание, передвижение, собирание, плавание, служат органом защиты и нападения на жертву.
Поперечно-полосатая мышечная ткань впервые возникла у членистоногих. Мышечная ткань дифференцирована на отдельные пучки. Она построены из длинных волокон, которые начинаются от одних члеников и крепятся к другим членикам тела, превращая их в подвижные рычаги.
Для членистоногих характерна гетерономная сегментация тела (от греч. heteros — другой и nomos — закон) — сегменты тела (членики) имеют разное строение и выполняют разные функции. Сегменты тела сливаются и образуют отделы — голову, груди и брюшко. Отделы тела также могут сливаться между собой.
Тело членистоногих покрыто твердым наружным покровом — хитинизированной кутикулой. Хитин — природный полимер из группы азотсодержащих полисахаридов. Такая оболочка надежно защищает все внутренние органы от механических повреждений, предотвращает высыхание и служит опорой для внутренних органов, мышц.
Перед вами одно из величайших событий эволюции на Земле — приобретение насекомыми крыльев. Это важное новшество дало возможность осваивать обширные ранее недоступные территории, заселить множество разных экологических ниш.
Перечисленные ароморфозы членистоногих и их маленькие размеры дали этим животным невероятные преимущества, и, повторюсь, мы с вами на данный момент живем в эру господства членистоногих. Они прекрасно уживаются с человеком — вы не раз встречали насекомых в своем жилище, и порой это были не самые лучшие и ожидаемые встречи, но ведь они были — это еще одно очевидное доказательство их адаптации к жизни рядом с нами.
Приступим к перечислению общих особенностей, характерных для членистоногих в целом.
Общие особенности членистоногих
- Покровы тела
- Гетерономная сегментация
- Членистые конечности
- Полость тела
- Кровеносная система
- Дыхательная система
- Выделительная система
- Нервная система
- Половая система
Снаружи тело членистоногих покрыто хитиновой кутикулой — твердым наружным скелетом (его также называют экзоскелет). Он выполняет защитную функцию, изнутри к нему крепятся многочисленные мышечные пучки.
Важно отметить, что хитиновый покров нерастяжим и сдерживает рост животного, поэтому запомните, что членистоногие растут только в период линьки, когда спадает хитиновая кутикула.
Тело членистоногих подразделяется на неоднородные сегменты, которые отличаются по строению и функции — такая сегментация называется гетерономная. Группы сегментов сливаются в отделы тела: голову — несущую органы чувств и конечности для захвата пищи, грудь — с расположенными на ней ходильными конечностями, и брюшко — задний отдел.
У паукообразных и ракообразных голова и грудь сливаются в единый отдел — головогрудь. У клещей тело вообще не дифференцировано на отделы, представлено единой монолитной структурой.
Членистые конечности — важная прогрессивная черта членистоногих, они представляют собой систему множество подвижно соединенных друг с другом рычагов. Между отдельными члениками в конечности образуются суставы, что обеспечивает большой объем двигательных возможностей.
Конечности могут видоизменяться, и в зависимости от выполняемой функции конечность получает разные названия: бегательная, прыгательная, копательная, плавательная, хватательная, собирательная и другие.
Как уже было сказано, хитиновая кутикула выполняет функцию экзоскелета, поэтому у членистоногих полость тела утрачивает опорную функцию. Вследствие гетерономной сегментации тела и смешения первичной полости тела со вторичной, у членистоногих полость тела называется — миксоцель (от греч. μῖξις — «смешение» и κοῖλος — «полый»).
Остатки вторичной полости тела (целома) можно обнаружить в протоках половых железах (гонад) и выделительных органах. За счет крови, заполняющей большую часть тела, полость тела членистоногих (и моллюсков) также называют гемоцель.
У членистоногих кровеносная система незамкнутого (лакунарного) типа. Из сосудов кровь попадает в щелевые промежутки и омывает внутренние органы, доставляя клеткам питательные вещества. Сердце располагается на спинной стороне тела.
По сосудам и полостям движется единственная тканевая жидкость насекомых — гемолимфа. Она не переносит кислород, в ней редко можно обнаружить гемоцианин. Чаще всего она бесцветна, либо желтовато-зеленоватого цвета. Тем не менее, гемоцианин (за счет содержания меди придает характерный голубой цвет), обнаруживается среди мечехвостов, некоторых паукообразных, ракообразных и насекомых.
У водных форм представлена жабрами, у наземных — листовидными легкими и трахеями.
Органы выделения членистоногих представлены видоизмененными метанефридиями. У насекомых имеются мальпигиевы сосуды, позволяющие максимально экономить воду, всасывая ее прямо из мочи. У ракообразных выделительная система представлена зелеными железами, располагающимися на головном конце тела.
Обитание на суше в целом требует от членистоногих сильной экономии жидкости, что вызывает изменения продуктов выделения. Так у паукообразных легко растворимые мочевина и аммиак заменяются малорастворимым гуанином, у насекомых мочевина и аммиак заменяется мочевой кислотой.
Нервная система узлового типа. Состоит из головного мозга (надглоточного ганглия), подглоточного ганглия и брюшной нервной цепочки. От ганглиев (нервных узлов) отходят нервные волокна, иннервирующие близлежащие внутренние органы и ткани.
Все членистоногие размножаются только половым путем. Большинство из них раздельнополые животные, развитие чаще непрямое (происходит с метаморофозом). Метаморфоз (греч. metamorphosis — превращение) — глубокое преобразование в строении организма, в результате которого личинка через стадию куколки превращается во взрослую особь.
Подтип Трилобитовые
Этих животных мы с вами могли увидеть, если бы переместились на 540 млн. лет назад, в самое начало палеозоя (кембрийский период). В кембрии они впервые появились, и тогда же начался их расцвет. Вымерли эти удивительные животные к концу пермского периода.
Для этих животных характерна гомономная сегментация, отсутствие дифференциации конечностей.
Значение членистоногих
Значение членистоногих для человека сложно переоценить. Некоторых членистоногих употребляют в пищу — раки, крабы, омары (лобстеры). Медоносные пчелы создают мед — ценнейшее природное «лекарство», из меда изготавливается множество других фармацевтических лекарств. Хитиновая кутикула наружного скелета используется в фармацевтике. Из яда паукообразных изготавливают снотворные средства.
Членистоногие доставляют и множество хлопот человеку. Часть из них являются переносчиками тяжелых заболеваний (комар рода Anopheles переносит возбудителя малярии — малярийного плазмодия, возбудителей различных инфекционных заболеваний могут переносить мухи, москиты, блохи, тараканы).
Ряд членистоногих напрямую вызывает заболевания у человека, к примеру — чесоточный зудень приводит к развитию чесотки. Многие насекомые наносят вред культурным растениям, выращиваемым человеком, это — саранча, клопы, жук-короед.
Источник: studarium.ru
Общая характеристика Членистоногих
Если говорить о распространённости и разнообразии видов, то данному типу просто нет равных в царстве животных.
Тип Членистоногих включает в себя такие классы:
- Ракообразные;
- Паукообразные;
- Насекомые;
- Многоножки.
Рис. 1. Классы Членистоногих
Классы имеют общие признаки, которые являются основополагающими для определения к данному типу. А также имеют собственные особенности, которых нет у сородичей.
К общим признакам относят:
- Членистое строение конечностей, которое и стало основой для названия типа.
- Хитиновый покров тела, который затрудняет рост животного, поэтому время от времени происходит линька.
- Полость тела смешанная, называется миксоцель, содержит внутренние органы и гемолимфу.
- Первичноротые.
- Тело симметричное, состоящее из трёх отделов: голова, грудь, брюшко.
- Кровеносная система незамкнутая.
- Нервная система представлена брюшной нервной цепочкой и первыми зачатками головного мозга – головной мозоль.
- Тело сегментировано.
- Отсутствует кожно-мускульный мешок. Вместо него имеются отдельные поперечно-полосатые мышцы, которые крепятся к стенкам сегментов тела, либо же к конечностям.
- Дыхательная система зависит от места обитания и может состоять из жабр, трахей или легочных мешков.
- Хорошо развиты органы чувств, у некоторых видов – сложные фасеточные глаза.
- Пищеварительная система представлена в виде сквозного строения и состоит из ротового отверстия, глотки, пищевода, желудка, печени, средней и задней кишки, анального отверстия.
- Выделительная система зависит от того, к какому классу относится животное. У Ракообразных животных это зелёные железы, у наземных видов – мальпигиевы сосуды.
- Характерно размножение раздельнополое, хотя встречаются гермофродиты. Оплодотворение внутреннее.
Рис. 2. Внутреннее строение Членистоногих
Индивидуальные особенности Членистоногих
Ниже представлена сравнительная таблица, которая кратко расскажет об индивидуальной характеристике классов Членистоногих.
Таблица «Характеристика классов Членистоногих»
Эволюция
Учёным до конца не известно происхождение Членистоногих, поэтому существует несколько гипотез. Первоначально предполагали, что данный тип произошёл от кольчатых червей, которые были покрыты жёсткой щетиной. Однако эволюционное доказательство родства с кольчатыми червями было опровергнуто в начале ХХ века.
Самыми древними предками Членистоногих являются трилобиты, которые жили в период Палеозойской эры. Изучая дальнейшую эволюцию типа, учёные пришли к выводу, что насекомые произошли от Ракообразных животных, а не от многоножек, как считалось ранее. Также доказано родство между Паукообразными, морскими пауками и меростомовыми.
Рис.3. Окаменелости первых Членистоногих
Значительную роль в развитии такого богатого разнообразия сыграло сегментное строение предков членистоногих, которое сохранилось и у современных видов. Многие части тела видоизменялись под воздействием среды обитания. Сегменты, как конструктор, сложились в различные виды, которые объединились в классы.
Источник: obrazovaka.ru
Членистоногие (рис. 104) произошли от каких-то древних многощетинковых кольчатых червей.
У этих червей предротовая лопасть слилась с первыми двумя сегментами тела, образовав акрон. На акроне расположены усики, первые — соответствующие конечностям второго сегмента и верхняя губа, расположенная перед ротовым отверстием, которая образовалась путем укорочения и слияния первой пары ног кольчатых червей.
Главным эволюционным новшеством членистоногих стал наружный скелет, образовавшийся благодаря уплотнению покровов. В панцирь в виде щитков, разделенных перепонками, оделись сегменты тела, ноги стали суставчатыми, состоящими из уплотненных колец, соединенных мягкими подвижными участками покровов. Как и у кольчатых червей, ноги вблизи основания несут жабры, но у членистоногих конечности стали еще выполнять третью роль — роль челюстей благодаря появлению жевательного выступа на внутренних поверхностях их сближенных оснований.
Первоначальное строение у членистоногих было простое: большое число одинаковых сегментов с ногами, спереди располагался акрон, сзади тельсон — хвостовая лопасть. Из яйца вылуплялась личинка (сходная с трохофорой) сразу с 4 первичными сегментами. Дальнейшее развитие шло за счет добавления сегментов между хвостовой лопастью и задним личиночным сегментом.
|
| Рис. 104. На рисунке показано каким образом из сегментированного кольчатого червя могло развиться насекомое. Первые пять сегментов постепенно слились и специализировались, образовав голову, следующие три объединились в грудь, а остальные — в брюшко |
Жесткий наружный скелет препятствует росту, поэтому потребовались линьки, во время которых старый покров отстает, лопается и сбрасывается, отделяясь от лежащего под ним пока еще неживого нового. Новый сначала растягивается, а затем затвердевает в более крупных формах. Так у членистоногих ступенчато, от линьки к линьке происходит рост.
Членистоногие возникли в воде как придонные организмы и их первичная эволюция шла в водной среде.
Наружный скелет открыл большие возможности усложнения и усовершенствования конструкции тела членистоногих, повысил их двигательную активность и создал возможность усложнения движений тела и его конечностей. Совершенствование конструкции тела членистоногих первоначально шло в сторону разделения функций отдельных участков тела и оформления их в своеобразно устроенные отделы, называемые тагмами. Передние сегменты вместе с акроном объединились в головной отдел, в котором сосредотачивались функции ориентации в пространстве и захвата и первичной обработки пищи, средний отдел брал на себя задачи передвижения с помощью ног, задний отдел сохранял и усиливал функцию пищеварения и размножения. По другим важнейшим функциям эти отделы различаются не столь явственно. Первоначально отделы обосабливались нерезко, это позволяло им частично менять свой сегментный состав, некоторые сегменты груди переходили к голове, некоторые сегменты брюшка присоединялись к груди, могло происходить также объединение и разделение отделов.
Наиболее древними ископаемыми членистоногими являются трилобиты и ракообразные. Материал с сайта http://doklad-referat.ru
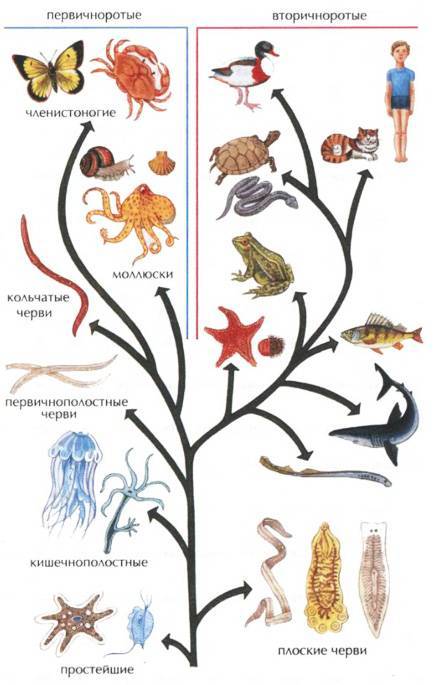
В каких родственных отношениях находятся членистоногие с другими животными показано на рис. 106.
|
| Рис. 105. Родственные связи между классами членистоногих по палеонтологическим данным |
|
| Рис. 106. Родственные отношения между типами животных, установленные методами сравнительной анатомии, сравнительной эмбриологии и палеонтологии |
Источник: doklad-referat.ru