Где образуются субъединицы рибосом
Рибосомы — субмикроскопические немембранные органеллы, необходимые для синтеза белка. Они объединяют аминокислоты в пептидную цепь, образуя новые белковые молекулы. Биосинтез осуществляется по матричной РНК путем трансляции.
Особенности строения
Рибосомы находятся на гранулярном эндоплазматическом ретикулуме или свободно плавают в цитоплазме. Крепятся они к эндоплазматической сети своей большой субъединицей и синтезируют белок, который выводится за пределы клетки, используется всем организмом. Цитоплазменные рибосомы в основном обеспечивают внутренние потребности клетки.

Форма шаровидная или овальная, в диаметре около 20нм.
На этапе трансляции к мРНК может прикрепляться несколько рибосом, образуя новую структуру – полисому. Сами же они образуются в ядрышке, внутри ядра.
Выделяют 2 вида рибосом:
- Малые – находятся в прокариотических клетках, а также в хлоропластах и митохондриальном матриксе. Они не связаны с мембраной и имеют меньшие размеры (в диаметре до 15нм).
- Большие – находятся в эукариотических клетках, могут достигать в диаметре до 23нм, связываются с эндоплазматической сетью или крепятся к мембране ядра.
Строение обоих видов идентичное. В состав рибосомы входят две субъединицы — большая и малая, которые в сочетании напоминают гриб. Объединяются они при помощи ионов магния, сохраняя между соприкасающимися поверхностями небольшую щель. При дефиците магния субъединицы отдаляются, происходит дезагрегация и рибосомы уже не могут выполнять свои функции.
Химический состав
Рибосомы состоят из высокополимерной рибосомальной РНК и белка в соотношении 1:1. В них сосредоточено примерно 90% всей клеточной РНК. Малая и большая субъединицы содержат около четырех молекул рРНК, которая имеет вид нитей собранных в клубок. Окружены молекулы белками и формируют вместе рибонуклеопротеид.
Полирибосомы – это объединение информационной РНК и рибосом, которые нанизываются на нить иРНК. В период отсутствия синтезирующих процессов, рибосомы разъединяются и обмениваются субъединицами. При поступлении иРНК они снова собираются в полирибосомы.
Количество рибосом может изменяться в зависимости от функциональной нагрузки на клетку. Десятки тысяч находятся в клетках с высокой митотической активностью (меристема растений, стволовые клетки).
Образование в клетке
Субъединицы рибосом формируются в ядрышке. Матрицей для синтеза рибосомальной РНК является ДНК. Для полного созревания они проходят несколько этапов:
- Эосома – первая фаза, при этом в ядрышке на ДНК синтезируется лишь рРНК;
- неосома – структура включающая не только рРНК, но и белки, после ряда модификаций выходит в цитоплазму;
- рибисома – зрелая органелла, состоящая из двух субъединиц.
| Функции элементов рибосом | ||
|---|---|---|
| Большая субъединица | Большая субъединица Треугольная, в диаметре 16нм, состоит из 3 молекул РНК и 33 белковых молекул Трансляция, декодирование генетической информации | Трансляция, декодирование генетической информации |
| Малая субъединица | Вогнутая, овальная, в диметре 14нм, состоит из 1 молекулы РНК и 21 белковых молекул | Объединение аминокислот, создание пептидных связей, синтез новых молекул белка |
Биосинтез белков на рибосомах
Трансляция или синтез белков на рибосомах с матрицы иРНК – конечный этап преобразования генетической информации в клетках. Во время трансляции информация, закодированная в нуклеиновых кислотах, переходит в белковые молекулы со строгой последовательностью аминокислот.
Трансляция – весьма непростой этап (в сравнении с репликацией и транскрипцией). Для проведения трансляции в процесс включаются все виды РНК, аминокислот, множество ферментов, которые могут исправлять погрешности друг друга. Самые важные участники трансляции – это рибосомы.
После транскрипции, новообразованная молекула иРНК, выходит из ядра в цитоплазму. Здесь после нескольких преобразований она соединяется с рибосомой. При этом аминокислоты приводятся в действие после взаимодействия с энергетическим субстратом – молекулой АТФ.
Аминокислоты и иРНК имеют разный химический состав и без постороннего участия не могут взаимодействовать между собой. Для преодоления этой несовместимости существует транспортная РНК. Под действием ферментов аминокислоты соединяются с тРНК. В таком виде они переносятся на рибосому и тРНК, с определенной аминокислотой, прикрепляется на иРНК в предназначенном месте. Далее рибосомальные ферменты формируют пептидную связь между присоединенной аминокислотой и строящимся полипептидом. После рибосома перемещается по цепи информационной РНК, оставляя участок для прикрепления следующей аминокислоты.
Рост полипептида идет до того момента, пока рибосома не встретит «стоп-кодон», который сигнализирует об окончании синтеза. Для освобождения новосинтезированного пептида от рибосомы включаются факторы терминации, окончательно завершающие биосинтез. К последней аминокислоте прикрепляется молекула воды, а рибосома распадается на две субъединицы.
Когда рибосома продвигается дальше по иРНК, она освобождает начальный отрезок цепи. К нему снова может присоединиться рибосома, которая начнет новый синтез. Таким образом, используя одну матрицу для биосинтеза, рибосомы создают одномоментно множество копий белка.
Роль рибосом в организме
- Рибосомы синтезируют белок для собственных нужд клетки и за ее пределы. Так в печени образуются плазменные факторы свертывания крови, плазмоциты продуцируют гамма-глобулины.
- Считывание закодированной информации с РНК, соединение аминокислот в запрограммированном порядке с образованием новых белковых молекул.
- Каталитическая функция – формирование пептидных связей, гидролиз ГТФ.
- Свои функции в клетке рибосомы выполняют более активно в виде полирибосом. Эти комплексы способны одновременно синтезировать несколько молекул белка.
Источник: animals-world.ru
(от «рибонуклеиновая кислота» и сома), органоид клетки, осуществляющий биосинтез белка. Представляет собой частицу сложной формы диам. ок. 20 нм. Р. состоит из 2 неравных субчастиц (субъединиц) — большой и малой, на к-рые может диссоциировать. Различают 2 осн. типа Р.— эукариотные (с константами седиментации: целой Р.— 80 S, малой субчастицы — 40 S, большой — 60 S) и прокариотные (соответственно 70 S, 30 S и 50 S). Кроме того, в митохондриях и хлоропластах содержатся мелкие Р. (константа седиментации 55 S— 70 S), осуществляющие автономный синтез белка. В бактериальной клетке содержится 104 — 105 Р.
состав Р. входит рРНК (3 молекулы у прокариот и 4 — у эукариот) и белки. Молекулы рРНК составляют 50—63% массы Р. и образуют её структурный каркас. Каждый из белков Р. представлен в ней одной молекулой, т. е. на одну Р. приходится неск. десятков разных белков (ок. 55 для Р. прокариот и ок. 100 для Р. эукариот). Большинство белков специфически связано с определ. участками рРНК. Нек-рые белки — т. н. факторы инициации (начала), элонгации (продолжения) и терминации (окончания) — входят в состав Р. только во время биосинтеза белка. В отсутствие биосинтеза белка субчастицы Р. находятся в динамич. равновесии с целыми Р. При начале трансляции с малой субчастицей связываются иРНК, формилметионил-тРНК и факторы инициации; затем этот комплекс присоединяется к большой субчастице. Связь оказывается очень прочной и исчезает только после терминации. Ассоциация выделенных субчастиц Р. осуществляется только при наличии двухвалентных катионов, в физиол. условиях в ней участвует Mg2+. P. имеет специфич. места для присоединения аминоацил-тРНК, пептидил-тРНК, места образования пептидной связи и гидролиза гуанозинтрифосфата, что обеспечивает постепенное скольжение Р. вдоль молекулы иРНК при синтезе полипептидной цепи. Одну молекулу иРНК могут одновременно транслировать неск. Р., образуя комплекс — полирибосому (полисому). Кол-во полирибосом в клетке указывает на интенсивность биосинтеза белка. В эукариотных клетках часть Р.
язана спец. белками большой субчастицы с мембранами эндоплазматич. сети. Эти Р. синтезируют в осн. белки, к-рые поступают в комплекс Гольджи и секретируются клеткой. Р; расположенные в гиалоплазме, синтезируют белки для собств. нужд клетки. У эукариот Р. образуются в ядрышке. На ядрышковой ДНК синтезируются предшественники рРНК, к-рые покрываются поступающими из цитоплазмы рибосомальными белками, расщепляются до нужных размеров и формируют рибосомные субчастицы, к-рые выходят в цитоплазму. Полностью сформированных Р. в ядре нет. Осн. массу клеточной РНК составляет рРНК. Она обусловливает базофильную окраску ядрышка и участков эргастоплазмы (напр., Ниссля вещества в нейронах).
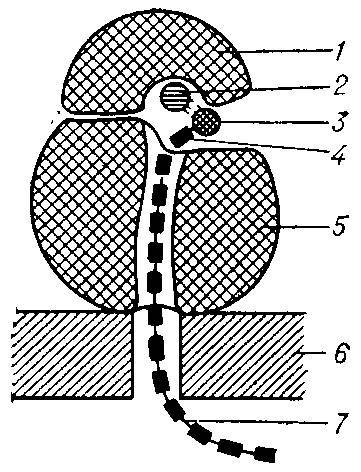
↑ Схема строения рибосомы, сидящей на мембране эндоплазматнческой сети: 1 — малая субъединица; 2 — иРНК; 3 — аминоацил — тРНК; 4 — аминокислота; 5 — большая субъединица; 6 — мембрана эндоплазматической сети; 7 — синтезируемая полипептидная цепь.
Источник: Биологический энциклопедический словарь на Gufo.me
Источник: gufo.me
Генетический код разных организмов обладает некоторыми общими свойствами:
1) Триплетность. Для записи любой, в том числе и наследственной информации используется определенный шифр, элементом которого является буква, или символ. Совокупность таких символов составляет алфавит.
дельные сообщения записываются комбинацией символов, которые называются кодовыми группами, или кодонами. Известен алфавит, состоящий всего из двух символов, — это азбука Морзе. В ДНК 4 буквы – первые буквы названий азотистых оснований (А, Г, Т, Ц), значит, генетический алфавит состоит всего из 4 символов. Что же является кодовой группой, или, словом генетического кода? Известно 20 основных аминокислот, содержание которых должно быть записано генетическим кодом, т. е. 4 буквы должны дать 20 кодовых слов. Допустим, слово состоит из одного символа, тогда мы получим только 4 кодовые группы. Если же слово состоит из двух символов, то таких групп будет только 16, а этого явно мало, чтобы закодировать 20 аминокислот. Следовательно, в кодовом слове должно быть минимум 3 нуклеотида, что даст 64 (43) сочетания. Такого количества триплетных сочетаний вполне достаточно для кодирования всех аминокислот. Таким образом, кодон генетического кода – это триплет нуклеотидов.
2) Вырожденность (избыточность) – свойство генетического кода состоящее с одной стороны, в том, что он содержит избыточные триплеты, т. е. синонимы, а с другой – «бессмысленные» триплеты. Поскольку код включает 64 сочетания, а кодируются только 20 аминокислот, то некоторые аминокислоты кодируются несколькими триплетами (аргинин, серин, лейцин – шестью; валин, пролин, аланин, глицин, треонин – четырьмя; изолейцин – тремя; фенилаланин, тирозин, гистидин, лизин, аспарагин, глутамин, цистеин, аспарагиновая и глутаминовая кислоты – двумя; метионин и триптофан – одним триплетом).
которые кодовые группы (УАА, УАГ, УГА) вообще не несут смысловой нагрузки, т. е. являются «бессмысленными» триплетами. «Бессмысленные», или nonsense, кодоны выполняют функцию терминаторов цепей – знаков препинания в генетическом тексте – служат сигналом окончания синтеза белковой цепи. Такая избыточность кода имеет большое значение для повышения надежности передачи генетической информации.
3) Неперекрываемость. Кодовые триплеты никогда не перекрываются, т. е. всегда транслируются вместе. При считывании информации с молекулы ДНК невозможно использование азотистого основания одного триплета в комбинации с основаниями другого триплета.
4) Однозначность. Нет случаев, когда один и тот же триплет соответствовал бы более чем одной кислоте.
5) Отсутствие разделительных знаков внутри гена. Генетический код считывается с определенного места без запятых.
6) Универсальность. У различных видов живых организмов (вирусов, бактерий, растений, грибов и животных) одинаковые триплеты кодируют одни и те же аминокислоты.
7) Видовая специфичность. Количество и последовательность азотистых оснований в цепи ДНК у разных организмов различные.
Лекция 5. Генетический код
Определение понятия
Генетический код — это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК.
Поскольку ДНК непосредственного участия в синтезе белка не принимает, то код записывается на языке РНК. В РНК вместо тимина входит урацил.
Свойства генетического кода
1. Триплетность
Каждая аминокислота кодируется последовательностью из 3-х нуклеотидов.
Определение: триплет или кодон — последовательность из трех нуклеотидов, кодирующая одну аминокислоту.
Код не может быть моноплетным, поскольку 4 (число разных нуклеотидов в ДНК) меньше 20. Код не может быть дуплетным, т.к. 16 (число сочетаний и перестановок из 4-х нуклеотидов по 2) меньше 20. Код может быть триплетным, т.к. 64 (число сочетаний и перестановок из 4-х по 3) больше 20.
2. Вырожденность.
Все аминокислоты, за исключением метионина и триптофана, кодируются более чем одним триплетом:
2 АК по 1 триплету = 2.
9 АК по 2 триплета = 18.
1 АК 3 триплета = 3.
5 АК по 4 триплета = 20.
3 АК по 6 триплетов = 18.
Всего 61 триплет кодирует 20 аминокислот.
3. Наличие межгенных знаков препинания.
Определение:
Ген — это участок ДНК, кодирующий одну полипептидную цепь или одну молекулу tPHK , r РНК или sPHK .
Гены tPHK , rPHK , sPHK белки не кодируют.
В конце каждого гена, кодирующего полипептид, находится, по меньшей мере, один из 3-х триплетов, кодирующих терминирующие кодоны РНК, или стоп-сигналы. В мРНК они имеют следующий вид: UAA , UAG , UGA . Они терминируют (оканчивают) трансляцию.
Условно к знакам препинания относится и кодон AUG — первый после лидерной последовательности. (См. лекцию 8) Он выполняет функцию заглавной буквы. В этой позиции он кодирует формилметионин (у прокариот).
4. Однозначность.
Каждый триплет кодирует лишь одну аминокислоту или является терминатором трансляции.
Исключение составляет кодон AUG . У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой — метионин.
5. Компактность, или отсутствие внутригенных знаков препинания.
Внутри гена каждый нуклеотид входит в состав значащего кодона.
В 1961 г. Сеймур Бензер и Френсис Крик экспериментально доказали триплетность кода и его компактность.
Суть эксперимента: «+» мутация — вставка одного нуклеотида. «-» мутация — выпадение одного нуклеотида. Одиночная «+» или «-» мутация в начале гена портит весь ген. Двойная «+» или «-» мутация тоже портит весь ген.
Тройная «+» или «-» мутация в начале гена портит лишь его часть. Четверная «+» или «-» мутация опять портит весь ген.
Эксперимент доказывает, что код тршплетен и внутри гена нет знаков препинания. Эксперимент был проведен на двух рядом расположенных фаговых генах и показал, кроме того, наличие знаков препинания между генами.
6. Универсальность.
Генетический код един для всех живущих на Земле существ.
В 1979 г. Беррел открыл идеальный код митохондрий человека.
Определение:
«Идеальным» называется генетический код, в котором выполняется правило вырожденности квазидублетного кода: Если в двух триплетах совпадают первые два нуклеотида, а третьи нуклеотиды относятся к одному классу (оба — пурины или оба — пиримидины), то эти триплеты кодируют одну и ту же аминокислоту.
Из этого правила в универсальном коде есть два исключения. Оба отклонения от идеального кода в универсальном касаются принципиальных моментов: начала и конца синтеза белка:
|
Кодон |
Универсальный код |
Митохондриальные коды |
|||
|
Позвоночные |
Беспозвоночные |
Дрожжи |
Растения |
||
|
STOP |
STOP |
||||
|
С UA |
|||||
|
А G А |
STOP |
||||
|
STOP |
230 замен не меняют класс кодируемой аминокислоты. к рываемость.
В 1956 г. Георгий Гамов предложил вариант перекрываемого кода. Согласно Гамовскому коду, каждый нуклеотид, начиная с третьего в гене, входит в состав 3-х кодонов. Когда генетический код был расшифрован, оказалось, что он неперекрываем, т.е. каждый нуклеотид входит в состав лишь одного кодона. Достоинства перекрываемого генетического кода: компактность, меньшая зависимость структуры белка от вставки или делеции нуклеотида. Недостаток: большая зависимость структуры белка от замены нуклеотида и ограничение на соседей. В 1976 г. была секвенирована ДНК фага φХ174. У него одноцепочечная кольцевая ДНК, состоящая из 5375 нуклеотидов. Было известно, что фаг кодирует 9 белков. Для 6 из них были определены гены, располагающиеся друг за другом. Выяснилось, что есть перекрывание. Ген Е полностью находится внутри гена D . Его инициирующий кодон появляется в результате сдвига считывания на один нуклеотид. Ген J начинается там, где кончается ген D . Инициирующий кодон гена J перекрывается с терминирующим кодоном гена D в результате сдвига на два нуклеотида. Конструкция называется «сдвиг рамки считывания» на число нуклеотидов, некратное трем. На сегодняшний день перекрывание показано только для нескольких фагов. Информационная емкость ДНК На Земле живет 6 миллиардов человек. Наследственная информация о них 4×10 13 книжных страниц. Эти страницы заняли бы объем 6-и зданий НГУ. 6×10 9 сперматозоидов занимают половину наперстка. Их ДНК занимает менее четверти наперстка. |
||||
Генетический код и его свойства
Запись генетической информации в виде последовательности нуклеотидов в молекулах ДНК и и-РНК, называется генетическим кодом .
Триплет нуклеотидов, кодирующий определенную аминокислоту, называется кодоном . Кодон – это элементарная функциональная единица гена.
Свойства генетического кода:
— триплетность : одной аминокислоте в молекуле полипептида соответствует один кодон;
— универсальность : у всех живых организмов один и тот же кодон определяет одинаковые аминокислоты;
— неперекрываемость : один нуклеотид входит в состав только одного триплета;
— вырожденность, или избыточность: одну аминокислоту может кодировать несколько триплетов (аминокислот – 20, возможных триплетов – 64);
— непрерывность (нет разделительных знаков между нуклеотидами);
— однонаправленность (образование и-РНК происходит в направлении от 3″ конца к 5″ концу).
Наличие среди триплетов инициирующих кодонов (с них начинается биосинтез белка), кодонов-терминаторов (обозначают конец биосинтеза белка).
Соответствие порядка нуклеотидов в молекуле ДНК порядку аминокислот в молекуле полипептида называется колинеарностью.
Гены характеризуются: специфичностью, целостностью, дискретностью, стабильностью, лабильностью, плейотропностью, экспрессивностью и пенетрантностью
1. Специфичность – уникальная последовательность нуклеотидов для каждого структурного гена.
2. Целостность – как функциональная единица (программирование синтеза белка) ген неделим.
3. Дискретность – в составе гена имеются субъединицы: мутон – субъединица, которая отвечает за мутацию; рекон – отвечает за рекомбинацию. Минимальная их величина – пара нуклеотидов.
4. Стабильность – гены относительно устойчивы (стабильны). Частота самопроизвольной мутации одного гена составляет примерно 1: 10-5 на поколение.
5. Лабильность – устойчивость генов не абсолютная, они могут изменяться, мутировать.
6. Плейотропия – множественное действие гена (один ген отвечает за несколько признаков).
7. Экспрессивность – степень фенотипического проявления гена. Она определяется факторами среды и влиянием других генов.
8. Пенетрантность – частота проявления гена: отношение (в процентах) числа особей, имеющих данный признак, к числу особей, имеющих данный ген.
Классификация генов
По функциям гены классифицируют на структурные и функциональные. Структурные гены содержат информацию о белках-ферментах, гистонах, о последовательности нуклеотидов в разных видах РНК.
Функциональные гены оказывают влияние на работу структурных генов. Функциональными являются гены-модуляторы и гены-регуляторы. Гены-модуляторы – это ингибиторы, интенсификаторы, модификаторы. Они усиливают, ослабляют или изменяют работу структурных генов. Регулируют работу структурных генов гены-регуляторы и гены-операторы .
Генотип всех соматических клеток организма одного вида одинаков. Но клетки разных тканей отличаются друг от друга. Вероятно, это связано с тем, что в них работают разные блоки генов. Область проявления действия данного гена называется полем его действия (например, распределение волосяного покрова на теле человека). Как правило, гены, детерминирующие определенные признаки, «работают» непостоянно (например, гены, определяющие синтез половых гормонов); их функция значительно снижается с возрастом. Период функционирования гена называется временем его действия .
По месту действия гены подразделяют на три группы
1) функционирующие во всех клетках (например, гены, кодирующие ферменты энергетического обмена);
2) функционирующие в клетках одной ткани (детерминирующие синтез белка миозина в мышечной ткани);
3) специфичные для одного типа клеток (гены гемоглобина в незрелых эритроцитах).
Гены выполняют в клетке две основные функции.
Гетеросинтетическая функция – это программирование биосинтеза белка в клетке.
Аутосинтетическая функция – репликация спирали ДНК (самоудвоение ДНК).
После открытия структуры ДНК долгое время полагали, что бактериальная хромосома представляет собой чистую ДНК в виде двойной спирали. Однако позднее выяснилось, что хромосома прокариот содержит в своей структуре примерно 20% белков. Их роль — обеспечить определенную компактизацию и прикрепление ДНК к оболочке бактерии. В настоящее время белки прокариотической хромосомы известны. Показано, что мутации в соответствующих генах не приводят к заметным фенотипическим проявлениям. По-видимому, роль этих белков вспомогательная, и они могут заменять друг друга в создании определенной структуры. Таким образом, прокариоты, в отличие от эукариот, не имеют высокоспециализированной системы организации хромосомы.
Хромосома эукариот состоит в основном из белков (50-60%) и ДНК, с незначительным количеством молекул РНК (до 10% от количества ДНК). Белки можно подразделить на гистоновые (половина или большая доля белков хромосомы) и негистоновые. В свою очередь гистоновые белки, доля которых в структуре хромосомы составляет до 80%, делятся на 5 основных классов: НЗ, Н4, Н2А и Н2В и Н1. Негистоновые белки (по большей части кислые, в отличие от гистонов) представлены большим количеством различных видов. Показано, что все они участвуют в образовании структур надмолекулярного уровня. Хромосомная ДНК эукариотической клетки упакована исключительно компактно. Например, самая маленькая хромосома человека — 22, содержит примерно 4.6*107 п.н., что соответствует длине 1,4 см. Во время митоза эта хромосома укорачивается до 2 мкм, т.е. становится в 7000 раз компактнее. Очевидно, чтобы достичь такой плотности упаковки и сохранить эффективность основных генетических процессов (как правило, связанных с локальной распаковкой), структура хромосомы должна иметь несколько уровней организации. Вещество хромосомы — хроматин. В этом термине подчеркивается способность вещества хромосомы к окрашиванию, видимое уже на стадии интерфазы. Химическая структура хроматина различается подлине хромосомы, а сам хроматин претерпевает различные уровни своей упаковки от интерфазы до метафазы клеточных делений.
Существуют две наиболее известные модели, объясняющие механизм упаковки хроматина. Согласно одной из них, наиболее известной в зарубежной литературе, нить ДНК претерпевает пять уровней компактизацни от 2 нм (ее собственный диаметр) до 1400 нм (высококонденсированная метафазная хромосома).
Низшим уровнем иерархической организации хромосом считается нуклеосомный. Нуклеосома состоит из кора (сердцевины, стержня) и намотанной на негоДНК(146 п.н„ 1,8 витка). Кор представляет собой гистоновый октамер Н2А, Н2В, НЗ, Н4 (по две молекулы каждого). Хроматин на этой стадии имеет вид «бусин» (глобул диаметром 11 нм), нанизанных на «нить» (молекулярную ДНК). Такая структура обеспечивает компактизацию примерно в 6-7 раз.
Вторая ступень компактизации — формирование хроматиновой фибриллы диаметром 30 нм. В этом процессе участвует гистон HI, который связывается с ДНК между нуклеосомными корами и сворачивает нуклеосомную фибриллу в спираль, наполобие соленоида, с шагом в 6-8 нуклеосом. Уровень компактизации на этом этапе достигает примерно 40.
Третий этап — петельно-доменный — наиболее сложный. Соленоидная фибрилла складывается, образуя петли различной длины. Общий уровень компак-тизации возрастает до 1000, но, очевидно, может различаться в различных районах хромосомы. Диаметр такой структуры в среднем составляет 300 нм., по-видимому, она наиболее типична для интерфазной хромосомы.
На четвертом этапе компактизации 300 нм-фибриллы дополнительно сворачиваются, образуя хроматиды диаметром примерно 600-700 нм.
Последняя, пятая, ступень компактизации (в 7000 раз) характерна для метафазной хромосомы; ее диаметр равен 1400 нм. Известна и другая схема компактизации хроматина, предложенная Ю.С. Ченцовым. Она основана на данных световой и электронной микроскопии. Согласно этой модели первым уровнем также является нуклеосомный. На втором этапе 8-Ю нуклеосом образуют глобулу, называемую нуклеомером. Ряд сближенных нуклсомеров формируют 20-30-нанометровую фибриллу. Третий уровень — хромомерный. Петли фибрилл ДНП, скрепленные негистоновыми белками, образуют розетковидные структуры. На четвертом — хромонемном уровне происходит их сближение с образованием структур, состоящих из петлевых доменов. Предполагается, что на следующем, пятом, уровне компактизации, характерном для хроматид, происходит спиральная укладка хромонемных нитей.
Код триплетен. В состав РНК входят 4
нуклеотида: А, Г, Ц, У. Если бы мы
пытались обозначить одну аминокислоту
одним нуклеотидом, то 16 из 20
аминокислот остались бы не
зашифрованы. Двухбуквенный код
позволил бы зашифровать 16 аминокислот
(из четырех нуклеотидов можно составить
16 различных комбинаций, в каждой из
которых имеется два нуклеотида) .
Природа создала трехбуквенный, или
триплетный, код. Это означает, что каждая
из 20 аминокислот зашифрована
последовательностью трех нуклеотидов,
называемых триплетом или кодоном. Из 4
нуклеотидов можно создать 64 различные
комбинации по 3 нуклеотида в каждой
(4*4*4=64). Этого с избытком хватает для
кодирования 20 аминокислот и, казалось
бы, 44 кодона являются лишними. Однако
это не так.
2. Код вырожден. Это означает, что каждая
аминокислота шифруется более чем одним
кодоном (от двух до шести) . Исключение
составляют аминокислоты метионин и
триптофан, каждая из которых кодируется
только одним триплетом. (Это видно из
таблицы генетического кода.) Тот факт,
что метионин кодируется одним триплетом
АУТ, имеет особый смысл, который вам
станет понятен позже (16).
3. Код однозначен. Каждый кодон шифрует
только одну аминокислоту. У всех
здоровых людей в гене, несущем
информацию о бета-цепи гемоглобина,
триплет ГАА или ГАГ, стоящий на шестом
месте, кодирует глутаминовую кислоту. У
больных серповидноклеточной анемией
второй нуклеотид в этом триплете заменен
на У. Как видно из таблицы, триплеты ГУА
или ГУГ, которые в этом случае
образуются, кодируют аминокислоту валин.
К чему приводит такая замена, вы уже
знаете из раздела о ДНК.
4. Между генами имеются «знаки
препинания». В печатном тексте в конце
каждой фразы стоит точка. Несколько
связанных по смыслу фраз составляют
абзац. На языке генетической информации
таким абзацем являются оперон и
комплементарная ему и-РНК. Каждый ген в
опероне кодирует одну полипептидную
цепочку — фразу. Так как в ряде случаев
по матрице и-РНК последовательно
создается несколько разных
полипептидных цепей, они должны быть
отделены друг от друга. Для этого в
генетическом коде существуют три
специальные триплета — УАА, УАГ, УГА,
каждый из которых обозначает
прекрдщение синтеза одной полипептидной
цепи. Таким образом, эти триплеты
выполняют функцию знаков препинания.
Они находятся в конце каждого гена.
Внутри гена нет «знаков препинания».
Поскольку генетический код подобен
языку, разберем это свойство на примере
такой составленной из триплетов фразы:
жил был кот тих был сер мил мне тот кот.
Смысл написанного понятен, несмотря на
отсутствие «знаков препинания. Если же мы
уберем в первом слове одну букву (один
нуклеотид в гене) , но читать будем также
тройками букв, то получится бессмыслица:
илб ылк отт ихб ылс ерм илм нет отк от
Нарушение смысла возникает и при
выпадении одного или двух нуклеотидов из
гена. Белок, который будет считываться с
такого испорченного гена, не будет иметь
ничего общего с тем белком, который
кодировался нормальным геном.
6. Код универсален. Генетический код един
для всех живущих на Земле существ. У
бактерий и грибов, пшеницы и хлопка, рыб
и червей, лягушки и человека одни и те же
триплеты кодируют одни и те же
аминокислоты.
Код триплетен. В состав РНК входят 4
нуклеотида: А, Г, Ц, У. Если бы мы
пытались обозначить одну аминокислоту
одним нуклеотидом, то 16 из 20
аминокислот остались бы не
зашифрованы. Двухбуквенный код
позволил бы зашифровать 16 аминокислот
(из четырех нуклеотидов можно составить
16 различных комбинаций, в каждой из
которых имеется два нуклеотида) .
Природа создала трехбуквенный, или
триплетный, код. Это означает, что каждая
из 20 аминокислот зашифрована
последовательностью трех нуклеотидов,
называемых триплетом или кодоном. Из 4
нуклеотидов можно создать 64 различные
комбинации по 3 нуклеотида в каждой
(4*4*4=64). Этого с избытком хватает для
кодирования 20 аминокислот и, казалось
бы, 44 кодона являются лишними. Однако
это не так.
2. Код вырожден. Это означает, что каждая
аминокислота шифруется более чем одним
кодоном (от двух до шести) . Исключение
составляют аминокислоты метионин и
триптофан, каждая из которых кодируется
только одним триплетом. (Это видно из
таблицы генетического кода.) Тот факт,
что метионин кодируется одним триплетом
АУТ, имеет особый смысл, который вам
станет понятен позже (16).
3. Код однозначен. Каждый кодон шифрует
только одну аминокислоту. У всех
здоровых людей в гене, несущем
информацию о бета-цепи гемоглобина,
триплет ГАА или ГАГ, стоящий на шестом
месте, кодирует глутаминовую кислоту. У
больных серповидноклеточной анемией
второй нуклеотид в этом триплете заменен
на У. Как видно из таблицы, триплеты ГУА
или ГУГ, которые в этом случае
образуются, кодируют аминокислоту валин.
К чему приводит такая замена, вы уже
знаете из раздела о ДНК.
4. Между генами имеются «знаки
препинания». В печатном тексте в конце
каждой фразы стоит точка. Несколько
связанных по смыслу фраз составляют
абзац. На языке генетической информации
таким абзацем являются оперон и
комплементарная ему и-РНК. Каждый ген в
опероне кодирует одну полипептидную
цепочку — фразу. Так как в ряде случаев
по матрице и-РНК последовательно
создается несколько разных
полипептидных цепей, они должны быть
отделены друг от друга. Для этого в
генетическом коде существуют три
специальные триплета — УАА, УАГ, УГА,
каждый из которых обозначает
прекрдщение синтеза одной полипептидной
цепи. Таким образом, эти триплеты
выполняют функцию знаков препинания.
Они находятся в конце каждого гена.
Внутри гена нет «знаков препинания».
Поскольку генетический код подобен
языку, разберем это свойство на примере
такой составленной из триплетов фразы:
жил был кот тих был сер мил мне тот кот.
Смысл написанного понятен, несмотря на
отсутствие «знаков препинания. Если же мы
уберем в первом слове одну букву (один
нуклеотид в гене) , но читать будем также
тройками букв, то получится бессмыслица:
илб ылк отт ихб ылс ерм илм нет отк от
Нарушение смысла возникает и при
выпадении одного или двух нуклеотидов из
гена. Белок, который будет считываться с
такого испорченного гена, не будет иметь
ничего общего с тем белком, который
кодировался нормальным геном.
6. Код универсален. Генетический код един
для всех живущих на Земле существ. У
бактерий и грибов, пшеницы и хлопка, рыб
и червей, лягушки и человека одни и те же
триплеты кодируют одни и те же
аминокислоты.
Источник: www.dosaaf-khab.ru