Клеточная оболочка образована клетчаткой
Оболочку растительной клетки, одну из структур, отличающих растительные клетки от клеток животного организма, обычно считают отложением, или секрецией, цитоплазмы. Она состоит из трех основных групп соединений: целлюлозы, лигнина и пектина. Целлюлоза и ряд близко-родственных соединений, например гемицеллюлоза, образуют твердый слой клеточной оболочки. В основном эти соединения представляют собой разветвленные длинные цепочки (полисахариды), образованные из простого сахара— глюкозы. Гемицеллюлоза близка к целлюлозе, но, помимо сахаров, содержит и другие соединения. Целлюлоза и ее производные — высокоокисляемые вещества. Лигнин состоит из сложной смеси химически близких соединений, в частности полимеров фенольной кислоты. Отложение лигнина (одревеснение) делает целлюлозные оболочки клетки прочными и неэластичными, устойчивыми к разрушению микроорганизмами. Лигнин вызывает пожелтение бумаги, поэтому при изготовлении высококачественной бумаги он должен быть вымыт из древесной пульпы. Пектины представляют собой водорастворимые полимеры галакту-роновой кислоты, которые при соединении с водой образуют золи и гели. Наиболее знакомый нам пектин это тот, который используется для затвердения джемов и желе.[ …]
Клеточная оболочка древесного волокна имеет несколько слоев: первичный, который называется наружной оболочкой волокна, и вторичный (стенка, состоящая, в свою очередь, из трех слоев: наружного, среднего и внутреннего). Между первичными стенками клеток находится слой межклеточного вещества, при помощи которого волокна соединяются друг с другом. Вторичная стенка относительно толстая и представляет собой главную массу объема клетки.[ …]
Клеточные оболочки у гонатозиговых обычно окружены слизистым слоем пектозы, который при действии метиленовой синьки и индиго обнаруживает явно волокнистую структуру. Волокна располагаются перпендикулярно поверхности клеточных стенок что указывает на присутствие пор в оболочке, через которые и выделяется слизь.[ …]
Клеточная оболочка обладает свойством накапливать, адсорбировать на своей поверхности ионы. Еще работами советских физиологов Д. А. Сабинина и И. И. Колосова (1940) было показано, что клеточные оболочки обладают способностью адсорбировать из раствора катионы и анионы. Адсорбция вначале происходит очень быстро, а затем резко замедляется. Это связано с насыщением адсорбирующей поверхности.[ …]
Клеточная оболочка состоит из клетчатки, пектина, лигнина, фосфолипидов и гемицеллюлозы. Пектин, содержащий уроновые кислоты, обладает благодаря находящимся в них Карбоксильным группам способностью обменивать Н-ионы (отдиссоциируемые этими группами) на другие катионы.[ …]
Участие клеточной оболочки в окислении Ре+2 было изучено с целью найти составные части клеток, окисляющие железо. Выделенные при этом большие количества фосфатидилсерина и фосфатидилэтаноламина являются компонентами наружной мембраны клеточной оболочки [42]. Присутствие больших количеств фосфатидилсерина обеспечивает отрицательный заряд, способствующий связывание Ре+2. Недавно эта гипотеза была подтверждена экспериментально [42а].[ …]
Первичная оболочка. На различных этапах онтогенеза постоянно меняются структура, химический состав и свойства клеточных оболочек, формирующихся от слияния мелких мембранных пузырьков (вакуолей) в экваториальной плоскости клетки. Вновь образовавшаяся оболочка молодой клетки представляет собой тонкую (0,5—1 мкм) эластичную мембрану, способную легко растягиваться. Оболочка зрелых дифференцированных клеток состоит из трех слоев: средний из них — межклеточное вещество, так называемая срединная пластинка, а два других принадлежат каждый соответственно двум соседним клеткам, составляя их собственные первичные оболочки, склеенные прослойкой из межклеточного вещества. Описанное строение характерно для меристематических и интенсивно растущих клеток.[ …]
Первичная клеточная оболочка — сложная структура. Пектиновые вещества, целлюлоза, гемицеллюлоза и другие соединения, входящие в состав клеточных оболочек, содержат карбоксильные группы, которые при диссоциации заряжают ее отрицательно.
составе оболочки имеются также белки, которые могут в зависимости от pH сообщать поверхности клеток отрицательный или положительный заряд. В результате этого клеточпая оболочка может выступать как иоеообмеп-ник, обратимо связывая катионы и анионы. На поверхности клеточной оболочки оказываются адсорбированными ионы Н+ и НСОз , которые в эквивалентных количествах меняются на ионы, находящиеся в свободном пространстве1. Благодаря тому что оболочка имеет значительное количество (межфибриллярных) промежутков, не занятых матриксом, у нее имеется большая внутренняя поверхность, которая и участвует в адсорбции иопов из свободного пространства. Ионы могут, таким образом, накапливаться в клеточной оболочке, и она служит как бы их резервуаром.[ …]
Реакция на лигнин Лигнификация клеточных оболочек древесины свидетельствует о прошедшем этапе вызревания побега и его подготовке к зиме. Существует две большие группы лигнина: компонент «Ф» — фло-роглюциновая реакция, компонент «М» — реакция Меуле с перманганатом. При одревеснении побегов и подготовке их к зиме появляется сначала лигнин «М», а потом лигнин «Ф». Таким образом, положительная реакция только с перманганатом свидетельствует о неполном одревеснении побега.[ …]
По данным Гизена и Холловей [363, 382], клеточная оболочка составляет 20—30% сухого веса бактериальной клетки.
е бактерии, за исключением микоплазм, миксобактерий и спирохет, имеют плотную и достаточно жесткую ригидную клеточную оболочку подобно растительным клеткам. Однако клеточные оболочки бактерий по своему химическому составу и структуре резко отличаются от клеточных стенок растений, которые, как известно, состоят из целлюлозы. У бактерий целлюлозы в оболочках нет, исключением являются уксусные бактерии Aceto-bacter xylinum и A. acetigenum, образующие толстую кожистую пленку из целлюлозных фибрилл.[ …]
Одиночная клетка E. coli окружена трехслойной клеточной оболочкой толщиной порядка 40 нм, представляющей собой «мешок» или «конверт», в котором заключено клеточное содержимое в виде, примерно, 2 х Ю 1Я г белка, 6 х 10 16 г ДНК и 2 х 10 14 г РНК (в основном ри-босомной РНК). В бактериальной клетке синтезируется около 2000 разных белков, большинство которых содержится в цитоплазме. Концентрация одних белков составляет 10“® М, тогда как других — порядка 2 х 10″4 М (от 10 до 200 000 молекул на клетку).[ …]
Содержимое бактериальной клетки без клеточной оболочки получило название протопласта. Протопласт состоит из цитоплазмы, покрытой мембраной. Разработан метод освобождения протопласта грамположительных бактерий посредством обработки клеток ферментом лизоцимом. Оболочки клеток при этом растворяются, а протопласты сохраняются живыми, способными к росту, делению, синтезу протеинов и нуклеиновых кислот [363]. Цитоплазма представляет собой водянистую или слегка вязкую массу — сложную композицию белков, жиров, углеводов и многочисленных других органических соединений, минеральных веществ и воды.
топлазма не гомогенная коллоидная жидкость, она содержит множество субми-кроскопических мембранных структур, выявленных электронной микроскопией. В цитоплазматических белках найдено 20 различных аминокислот, обусловливающих различные евойства белков. Например, аминокислота тирозин имеет спиртовые группы (ОН) в боковой цепи и этим обусловливает гидрофильность цитоплазмы. Липоиды, наоборот, обусловливают гидрофобность цитоплазмы.[ …]
Дрожжевая клетка состоит из 75% воды и 25% сухих веществ. В клеточной оболочке сосредоточено около 20% сухого вещества клетки.[ …]
В смешанные биоценозы входят бактерии, простейшие, грибы. Клеточная оболочка бактерий окружена слизистым слоем — капсулой. Капсула образуется вокруг группы бактериальных клеток, связанных вместе, образуя так называемую зооглею. Вещества .капсулы может быть использовано как пищевой резерв и как средство защиты от неблагоприятных условий. У одних видов бактерий капсулы состоят из полисахаридов, у других — из полипептидов, что оказывает влияние на свойства активного ила (способность к агрегации, связывание воды и т. д.). Активный ил, населенный смешанными биоценозами, отличается лучшей структурой, способностью к оседанию, что оценивается иловым индексом .[ …]
К таким особенностям относятся: наличие упругой и прочной клеточной оболочки; значительное, развитие вакуолярной системы, в большой степени определяющей осмотические свойства клетки; существование пластид, способствующих прохождению первичного синтеза органических веществ из углекислого газа и воды под влиянием солнечной энергии; преобладание в-, клетках процессов синтеза над процессами освобождения энергии.[ …]
На рисунке 56 схематично представлен процесс образования новой клеточной оболочки после митоза. Утолщение срединной пластинки осуществляется благодаря присоединению к ней с обеих сторон новых пузырьков Гольджи, вследствие чего молодая клеточная оболочка приобретает бугристую поверхность и превращается в так называемую первичную оболочку. Это вновь возникшее трехслойное образование состоит из изотропного геля, гемицеллюлоз и пектиновых веществ. Так образуется матрикс клеточной оболочки» представляющий собой аморфную пластичную массу сильно гидратированных углеводов.[ …]
В образующем бактериальные клеточные оболочки мукопептиде содержится значительное количество остатков D-аналина с «неприродной» конфигурацией, что увеличивает устойчивость этих оболочек к действию протеолитических ферментов.[ …]
Характерной особенностью растительной клетки является палпчие твердой клеточной стоики. Неточная оболочка придает клеткам п тканям растении мехапкчоскую прочность, защищает протоплазма-тическую мембрану ст разрушения под влияпием гидростатического давления, развиваемого внутри клетки. Клеточная обо дочка, особенно в молодых клетках, играет определенную физиологическую роль.
а способпа к росту, является противоипфекциолпым барьером, принимает участие п поглощении минеральных веществ, являясь своеобразным попообмеипнком. Клеточная оболоч!» облагает такими свойствами, которые поаволяют противостоять давлециго воды внутри клетки, в в то же время обладает растяжимостью п способностью к росту. Она образуется из веществ, вырабатываемых компонентами протопласта. Первичная клеточная оболочка характерна для молодых клеток (рис. 5). По мере их старения образуется вторичная структура.[ …]
Цитоплазма окружена плазматической мембраной, прилегающей к стенке клетки. Клеточная оболочка частично проницаема для всех растворимых веществ и растворителей; плазматическая мембрана полупроницаема. Она состоит из липопротеинов, которые из-за их эластичности и высокой проницаемости относят к жировым веществам. Плазматическая мембрана, несмотря на ее микроскопическую толщину, была видима в плазмолизированных клетках. С помощью электронного микроскопа было открыто, что она служит не просто границей раздела клеток, но имеет определенную структуру. Клетка кажется пронизанной внутренними мембранами, которые вместе с наружной мембраной составляют систему, называемую эндоплаз-матияеской сетью.[ …]
Кроме му-реина оболочка содержит и другие компоненты, не связанные ковалентно с муреи-ном. Полисахариды непостоянного состава иногда содержат остатки сахаров — глюкозы, галактозы, рамнозы и ман-нозы.
тигенные свойства грамположительных бактерий обусловливаются тейхоевыми кислотами и полисахаридами клеточной оболочки. Химический состав оболочек бактерий резко отличен от состава оболочек растений и животных. Этим и обусловлена чувствительность бактерий к таким лекарственным веществам, как антибиотики, не действующим на животные клетки. В состав оболочки входят простети-ческие группы ферментов — коэнзимы, обусловливающие первый этап обмена веществ при контакте бактерий с питательными веществами. В этом слое происходит дробление макромолекул.[ …]
Растительная клетка представляет собой осмотическую систему. Пектоцеллюлозпая оболочка хорошо проницаема как для воды, так и для растворевпых веществ. В этом можно убедиться, рассмотрев явлепия плазмолиза в тургора. Если поместить клетку в раствор более высокой концентрации, чем в клетке, то под микроскопом видно, что протоплазма отстает от клеточной оболочки. Это особенно хорошо проявляется на клетках с окрашенным клеточным соком. Клеточный сок остается внутри вакуоли, а между протоплазмой и оболочкой образуется пространство, заполненное внешним раствором. Явление отставания протоплазмы от клеточной оболочки получило название плазмолиза. Плазмодия происходит в результате того, что под влиянием концентрированного внешнего раствора вода выходит из клетки, тогда как растворенные вещества остаются в клетке. При помещении клеток в чистую воду или в слабо концентрированный раствор, вода поступает в клетку. Количество йоды в клетке увеличивается, объем вакуоли возрастает, клеточный сок давит па цитоплазму и прижимает ее к кле-точпой оболочке. Под влиянием внутреннего давления клеточная оболочка растягивается, в результате клетка переходит в напряженное состояние (тургор).[ …]
Одним из важнейших жирсодержащих биологических объектов являются липидные мембраны — основа клеточных оболочек любого живого организма — от водорослей и бактерий до человека. Сами мембраны представляют собой очень сложный комплекс липидов, белков» Ь углеводов, поэтому имеет смысл рассматривать взаимодействие с водой компонентов этого комплекса по отдельности.[ …]
Как только на поверхности клетки оорбируется избыточное количество оеребра, последнее проникает внутрь клетки и задерживается цитоплазматической мембраной. В цитоплазматической мембране расположены основные ферментные системы клетки. Серебро блокирует бактериальные ферменты, в результате чего клетка гибнет (бактерицидный эффект).[ …]
В дифференцирующихся клетках камбия пузырьки Гольджи, приближаясь к поверхности клетки, укрупняются и, образуя выпячивания, захватывают гиалоплазму для растяжения клеточной оболочки и одновременного увеличения поверхности плазма-леммы.[ …]
Сосущая сила клетки. Величина осмотического давления имеет большое значение для определения силы, которая вызывает поступление воды в клетку. Одпако надо учесть, что клеточная оболочка, свободно пропуская воду и питательные вещества, обладает ограниченной растнжимостью.
и поступлении в клетку воды в ней развивается гидростатическое давление, которое заставляет плазмалемму прижиматься к клеточной оболочке. Его называют тургорным давлением (Т). Клеточная оболочка растягивается и, в свою очередь, оказывает нротиводавлеиие (гг?); оно тем больше, чем больше поступает воды в клетку. Благодаря ограниченной растяжимости клеточной оболочки наступает такой момент, когда давление оболочки целиком уравновесит силу осмотического поступления воды. При этом p =T = w. Насыщеппая водой клетка находится в максимально растянутом состоянии. Одпако у наземных растений такое состояние насыщенности крайне редко. В обычных условиях осмотическое давление не уравновешено полностью противодавлением (р>ы>). Это показывает, что клеточная оболочка еще пе полпостыо растянута и вода может поступать в плетку. Рязнпца между осмотическим давлением клеточного сока и противодавлением клеточной оболочки определяет сосущую силу (5) — это сила, с которой клетка притягивает воду в каждый данный момент. Имепио сосущая сила определяет поступление воды в клетку. Лишь в состоянии плазмолиза клетка всасывает воду с силой, равпой всей величине осмотического давления S = p. Это происходит потому, что в состоянии плазмолиза или завядапия вода не давит на клеточную оболочку. Тургорпое давление (Т) =0 и соответствующее противодавление клеточной оболочки также равно 0. По мере поступления воды в клетку появляется тургорпое давление, а следовательно, развивается и противодавление клеточной оболочки. В этом случае сосущая сила клетки будет равпа разности между осмотическим давлением и противодавлением оболочки S—р—w. Чем больше поступает воды в клетку, тем больше растет тургорное давление и противодавление оболочки. Наконец, наступает такой момент, при котором клеточная оболочка растягивается до предела. В этом случае осмотическое давление целиком уравновешивается противодавлением оболочки p = T = w, а сосущая сила становится равной нулю.[ …]
Для питания микроорганизмов необходимы соединения.углерода, лучшим источником которого являются углеводы. Они используются для синтеза белков и жиров, для образования клеточных оболочек и как энергетический материал в дыхательных и других процессах, происходящих в микробных клетках. Из углеводов для питания, например, дрожжей используются главным об1разом сахара. В качестве углеродистого питания применяются органические кислоты и их соли (молочная, уксусная, яблочная, янтарная), а также некоторые спирты (этиловый, мап-нит). В последнее время при помощи «меченых» атомов установлено, что дрожжи и бактерии для синтеза жиров могут использовать уксусную кислоту, которая превращается в жирные кислоты.[ …]
Слоевище улотрикса состоит из неразветвлен-ных нитей неопределенной длины, которые в начале роста прикрепляются к субстрату базальной клеткой. Клетки нитей цилиндрические или слегка бочонковидные, часто короткие. Клеточные оболочки обычно тонкие, но нередко они утолщаются и могут становиться слоистыми. Клетки улотрикса, так же как клетки всех водорослей этого порядка, содержат единственный пристенный хлоропласт с одним или несколькими пиреноидами и одно ядро, расположенное по продольной оси клетки. Хлоропласт имеет форму пояска, который опоясывает весь протопласт или только часть его (рис. 216, 3).[ …]
Таким образом, энергетическое состояпие воды, способность совершать работу и ее подвижность определяются водным потенциалом. Водный потенциал чистой воды приравнивается к пулю. Всякое присутствие растворенных веществ в клеточном соке уменьшает способность воды производить работу. Водный потенциал клеточного сока всегда меньше потенциала чистой воды. При этом вода передвигается в сторопу меньшего (более отрицательного) водного потенциала. Тургориое давление увеличивает водпый потенциал внутри клетки н при максимальном растяжепии клеточной оболочки делает его равным нулю (как 8 чистой воде), вода перестает поступать в клетку.[ …]
Электронно-плотный материал формирует срединную пластинку (5), разделяющую две первичные оболочки (4)> Мембраны пузырьков Гольджн образуют новую нлазмалемму [ …]
Как известно, мицедиадьные грибы содержат значительно меньше нуклеиновых кислот, пуринов» чем дрожжи, и обладают широким набором ферментов, гидролизующих отдельные трудно перевариваемые животными полимеры (клетчатку, гемицеллюлозы и др.) клеточной оболочки растений до мономеров — глюкозы, ксилозы и др.[ …]
Подобная картина внутриклеточных изменений и такая же последовательность нарастающих во времени нарушений наблюдались и при воздействии ультразвуковых волн на дрожжевые клетки [183, 182]. Общими для них являются набухание протоплазмы и оводне-ние клеточных структур, растворение ядер в цитоплазме или клеточном соке. К ним же относят повышение проницаемости клеточных оболочек и изменение процессов прижизненной окраски. Гибель клетки сопровождается коагуляцией протоплазмы и цитоплазматических белков. Дезинтеграция и диспергирование коагулированного вещества наступают уже после гибели клетки.[ …]
Кальций усиливает обмен веществ в растениях, играет важную роль в передвижении углеводов, оказывает влияние на превращение азотистых веществ, ускоряет распад запасных белков семени при прорастании. Кроме того, он имеет существенное значение для построения нормальных клеточных оболочек и для установления благоприятного кислотно-щелочного равновесия в растениях.[ …]
Пектиновые вещества красных водорослей представляют собой соли кальция и магния особых пектиновых кислот. Они обладают способностью растворяться в кипящей воде с образованием слизистых растворов. К группе пектиновых веществ относятся также особые коллоидные вещества, которые содержатся в клеточных оболочках и межклетниках многих багрянок. Они представляют собой сложную смесь содержащих серу полисахаридов и носят общее название фикоколлоидов. Фико-коллоиды не растворяются в холодной воде, но хорошо растворяются в кипящей с образованием коллоидных систем. В настоящее время фикоколлоиды красных водорослей, так же как и бурых, являются самыми важными продуктами, получаемыми из морских водорослей. Поскольку эти вещества широко применяются в хозяйственной жизни человека, их химический состав достаточно хорошо изучен. Фикоколлоиды получены из многих видов, в результате установлено несколько их разновидностей. Эти вещества отличаются друг от друга по составу и свойствам, но обладают общей желирующей способностью.[ …]
Можно предполагать, что интенсивный рост мицелия при использовании нерастворимых форм целлюлозы, высокая активность ее трансформации в белок и низкая активность внеклеточных целлюлаз обусловлены, с одной стороны, более прочной сорбцией отдельных компонентов целлюлазного комплекса на поверхности клеточной оболочки« с другой — высокой интенсивностью утилизации низ к смоле ку лярных углеводов, образуемых в результате конверсии целлюлозных суботратов.[ …]
Для технической реализации наиболее простым способом является щелочная обработка суспензии микроорганизмов, например дрожжей или активного ила. При выдерживании суспензии микроорганизмов при pH выше 8,5—9,0, температуре около 60—90 °С в течение 1—4 ч в жидкую среду выделяются растворимые белки, которые вместе с клеточными оболочками могут служить достаточно эффективными флокулянтами [45]. При этом имеет важное значение состояние исходной культуры микроорганизмов. Для получения биофлокулянтов следует применять по возможности физиологически активную культуру.[ …]
Вещества, тормозящие рост микробов. Химических веществ, которые способны угнетать рост микробов очень много. Максимальное бактерицидное действие каждого химического вещества проявляется лишь при определенной его концентрации. Это объясняется тем, что растворы дезинфицирующих веществ могут проникать через клеточную оболочку лишь при определенном их разведении. Так, наиболее сильное бактерицидное действие на микробы проявляется при концентрации карболовой кислоты 5%, этилового спирта 70% и т. д. Основными дезинфицирующими агентами являются: карболовая кислота, алкоголь, хлор, хлорная известь, азохлорамид, дихлорамин, толуол озон и новый дезинфектор — азид натрия (КаЫз).[ …]
Вода содержится в живых клетках, в мертвых элементах ксилемы и в межклетниках. В межклетниках вода находится главным образом в парообразном состоянии. Основными испаряющими органами растения являются листья. В связи с этим естественно, что наибольшее количество паров воды заполняет межклетники листьев. В жидком состоянии вода находится в различных частях клетки: клеточной оболочке, вакуоли, протоплазме. Вакуоли — паиболее богатая водой часть клетки, где содержание ее достигает 96%. Наименьшее содержание воды характерно для клеточных оболочек. Вода в клеточных оболочках заполняет промежутки между фибриллами целлюлозы и удерживается силами поверхностного натяжения в менисках. Часть молекул воды находится в адсорбированном состоянии на поверхности фибрилл, составляющих клеточную оболочку. Количественное определение содержания воды в клеточных оболочках затруднено; по-видимому, оно колеблется от 30 до 50%.[ …]
Вследствие достаточно сложного состава таких многокомпонентных смесей, каковыми являются биомассы и среды для культивирования микроорганизмов, трудно найти какой-либо универсальный метод выделения микробных метаболитов. Решение задачи затруднено тем, что образующиеся соединения мало устойчивы. Тем не менее, указанные выше способы выделения микробных метаболитов и клеточных оболочек позволяют в ряде случаев успешно решить поставленные практические задачи.[ …]
Многие химики возражали против термина гемицеллюлоза и пытались дать новую терминологию. Хуземан [7] в соответствии с терминологией, предложенной Штаудингером и Рейнеке [8], использует термин поли-озы, весьма популярный среди европейских химиков. Каррер [9] предпочитает классифицировать полисахариды клеточной оболочки по содержащимся в них сахарам (ксилан, арабан, маннан и т. д.); Гесс [10] говорит о них как о «сопутствующих углеводах». Первые относятся к углеводам, тесно связанным с целлюлозой (и, как полагает Норман, не содержащим групп уроновой кислоты), вторые содержат типичные группы уроновых кислот и (как предполагают) тесно ассоциируются с лигнином. Гемицеллюлозы можно было бы также назвать нецеллюлозными полисахаридами клеточной стенки, но этот термин громоздок, и, поскольку деление на истинную целлюлозу и стойкие гемицеллюлозы является произвольным, то тоже не подходит. Таким образом, терминология Шульце, хотя может быть и не совершенная, вероятно будет сохранена, и мы будем пользоваться ею в нашей монографии.[ …]
Первый этап поступления солей в клетку (поступление в свободное пространство) — зто пассивный процесс. Свободное пространство клетки это пространство, в которое ионы могут пропикать пассивно, путем диффузии по градиенту концентрации. О локализации свободного пространства в литературе единого мнепня нет. По-видимому, прежде всего это заполненные водой межфибриллярные пространства в клеточной оболочке, затем это промежутки между клеточной оболочкой и мембраной.[ …]
Одно из наиболее важных преимуществ сернистой кислоты перед фугами консервантами заключается в ее высокой асептической актив-юсти. Сущность консервирующего действия сернистой кислоты (НгБОз), исходя из физико-химических свойств и химической природы »того консерванта заключается в следующем. Сернистая кислота растворяется в липидно-протеиновой мембране, выстилающей с внутренней стороны клеточную оболочку микроорганизма, и проникает путем /льтрафильтрации через мельчайшие поры этой полупроницаемой мемб-заны в плазму. При этом нарушается структура цитоплазматической зболочки и морфологически изменяется клетка. Сернистая кислота 1егко окисляется в растворах; восстановительный характер ее является причиной изменения оптимальных для микроорганизмов значе—шй окислительно-восстановительного потенциала. Нарушение жизненных функций микроорганизмов вызывается тем, что сернистая кислота вступает в соединения с промежуточными продуктами их жизнедеятельности и ферментами. Все это приводит к глубокому нарушению обмена веществ и гибели микробной клетки.[ …]
На рис. 9 представлена схема передачи кислорода от газового пузырька 1 с пограничной пленкой газа 2 через поверхность фазового контакта 3 (газ — жидкость), дополнительную пленку 4, образуемую например ПАВ, границу раздела 5 и пограничный слои жидкости 6 в основную массу перемешиваемой жидкости 2. Далее передача кислорода идет по пути преодоления пограничной пленки 8 жидкости вокруг микробиальной клетки, после чего через клеточную оболочку 9 кислород поступает в клетку 10. Профиль концентрации кислорода изображает ломаная линия а — д, крутизна которой тем выше, чем выше сопротивление массопередаче на отдельных участках. Так, на участке а — d, где заметную роль играют процессы молекулярной диффузии, скорость рстворения кислорода зависит главным образом от площади поверхности контакта фаз, определяемой размером газовых пузырьков, а также наличием загрязнений в жидкости. На участке d — е, имеющем пологий характер и отражающем влияние главным образом конвективной диффузии, концентрация кислорода не меняется или меняется незначительно, так как гидродинамика реальных сооружений характеризуется режимом развитой турбулентности. И, на конец, на участке е — g уклон профиля концентрации кислорода при постоянстве факторов определяется- главным образом скоростью его потребления микробиальной клеткой, зависящей в свою очередь от скорости поступления питательных веществ, иначе говоря от технологических нагрузок по органическим загрязнениям.[ …]
Они входят в состав полимеров, составляющих срединную пластинку (ламеллу! и наружные слои клеточных оболочек растений, в комплексе с другими полимерами — целлюлозой, гемицеллюлозой. Характерным первичным действием пектиназ на паренхимные ткани растений является их мацерация (размягчение) и распад на отдельные клетки. Наличие внеклеточных пектиназ определяли визуально по интенсивности роста биомассы испытуемых грибов и по наличдо продуктов ферментативного расщепления пектина в культуральной жидкости (табл 33).[ …]
Из теоретических исследований в направлении повышения продуктивности лесов все большее значение приобретает изучение фотосинтеза и деятельности камбия древесных пород в различных лесорастительных условиях, а на основе этого изучения — применение методов воздействия на камбий в желательном для лесного хозяйства направлении. Здесь речь идет не только о количественной, но и о качественной стороне прироста, так как с деятельностью камбия связаны, например, соотношение ранней и поздней древесины в годичных слоях, утолщение клеточных оболочек и т.д.[ …]
Микроорганизмы «е имеют специальных органов питания. Питательные вещества попадают в клетку, а продукты жизнедеятельности выделяются из нее через всю поверхность клетки благодаря процессам диффузии, осмоса и адсорбции. Поскольку клетка является живым организмом, то она постоянно усваивает питательные вещества и, следовательно, их концентрация внутри клетки всегда меньше, чем снаружи. Это обусловливает постоянный процесс диффузии. Если при этом процессе вещества проходят через полупроницаемую перегородку, то такой процесс называется осмосом. Роль полупроницаемой перегородки выполняет клеточная оболочка и цитоплазматическая мембрана. Внутрь клетки через перегородку ¡проходят вода, ионы (противоположного клетке заряда) и мелкие молекулы (сахароза, глюкоза). Высокомолекулярные вещества (крахмал, клетчатка, белки) не могут проникнуть в клетку. Вода движется от меньшего осмотического давления к большему, а растворенное вещество — в обратном направлении. Согласно закону парциального осмотического давления общее осмотическое давление раствора равно сумме парциальных осмотических давлений отдельных растворенных веществ. Этот закон объясняет причину попадания в клетку растворов очень малой концентрации.[ …]
В антиподах отмечено четкое разграничение в вакуолизации микропилярного и халазального полюсов. В функции этих клеток входят транспорт питательных веществ из халазы в зародышевый мешок, а также участие в преобразовании поступающих метаболитов. Являясь аппаратом питания зародышевого мешка, антиподы на определенном этапе его развития становятся центром абсорбции, переработки и транспорта питательных веществ из халазы семяпочки в зародышевый мешок, т. е. являются как бы пищеварительными клетками. У аконита антиподы сохраняются вплоть до созревания семени; они окружены четко выраженной, но неоднородной клеточной оболочкой: в базальной части она втрое толще, чем в апикальной; внутренняя поверхность ее имеет многочисленные разветвленные выросты. Наиболее сильно эти выросты развиты в местах непосредственного контакта антипод друг с другом. Плазмалемма антипод имеет многочисленные инвагинации самой разнообразной формы.[ …]
После возрастания концентрации кислорода в атмосфере и достижения уровня 10% от современного (2-я точка Пастера) озоновый слой стал настолько эффективно защищать живое от жесткого ультрафиолетового излучения, что жизнь постепенно вышла из водной среды на сушу. Дальнейшее формирование наземных экосистем пошло относительно автономно от процессов эволюции водных экосистем. Развитие наземной зеленой растительности обеспечило большие количества кислорода и пищи, которые были необходимы для последующей эволюции таких крупных животных, как динозавры и млекопитающие, а также человека. Одновременно океанический планктон дополнительно к формам с клеточными оболочками из органических веществ пополнился формами с известковыми, а позже и с кремниевыми оболочками.[ …]
К величинам, которые могут служить основой для выражения результатов, относятся сырая и сухая масса ткани, содержание белкового азота, содержание ДНК, площадь листа; результаты выражаются в расчете на растение, на лист или на клетку. Данные о сырой массе получить легче всего, и соответствующие измерения издавна проводились наиболее широко. Этот метод является удовлетворительным, когда нужно обнаружить сильно выраженные различия на самых ранних стадиях после заражения. Однако в результате вирусной инфекции содержание воды в тканях вскоре после заражения может нарушаться. В болое поздние сроки возможно прекращение роста, что обусловливает серьезные трудности с использованием этого параметра. В зараженных вирусом листьях концентрация многих компонентов на единицу сирой массы может оказаться выше из-за того, что листья у таких растений имеют меньшие размеры, чем здоровые листья того же возраста. В то же время если сравнивать листья одинакового размера, то концентрация этих компонентов на единицу сырой массы в зараженных листьях может оказаться ниже, чем в здоровых. Аналогичные осложнения возникают также при измерениях сухой массы. Сам вирус может составлять около 10% сухой массы листа. В результате остановки роста доля инертных компонентов клеточной оболочки возрастает, а за счет накопления крахмала, вызванного вирусной инфекцией, может увеличиться сухая масса. Содержание белкового азота также может значительно изменяться в связи с присутствием вируса и с общей остановкой роста.[ …]
Источник: ru-ecology.info
Строение клеточной оболочки
Основу клеточной оболочки составляет плазматическая мембрана (наружная клеточная мембрана, плазмолемма) — биологическая мембрана, ограничивающая внутренние содержимое клетки от внешней среды.
Все биологические мембраны представляют собой двойной слой липидов, гидрофобные концы которых обращены внутрь, а гидрофильные головки — наружу.
Кроме липидов в состав мембраны входят белки: периферические, погруженные (полуинтегральные) и пронизывающие (интегральные). Периферические белки прилегают к билипидному слою с внутренней или внешней стороны, полуинтегральные — частично встроены в мембрану, интегральные — проходят через всю толщу мембраны. Белки способны перемещаться в плоскости мембраны.
Мембранные белки выполняют различные функции: транспорт различных молекул; получение и преобразование сигналов из окружающей среды; поддержание структуры мембран. Наиболее важное свойство мембран — избирательная проницаемость.
Плазматические мембраны животных клеток имеют снаружи слой гликокаликса, состоящий из гликопротеинов и гликолипидов и выполняющий сигнальную и рецепторную функции. Он играет важную роль в объединении клеток в ткани.
Плазматические мембраны растительных клеток покрыты клеточной стенкой из целлюлозы. Поры в стенке позволяют пропускать воду и небольшие молекулы, а жесткость обеспечивает клетке механическую опору и защиту.
Функции клеточной оболочки
Клеточная оболочка выполняет следующие функции:
- определяет и поддерживает форму клетки;
- защищает клетку от механических воздействий и проникновения повреждающих биологических агентов;
- отграничивает внутреннее содержимое клетки;
- регулирует обмен веществ между клеткой и окружающей средой, обеспечивая постоянство внутриклеточного состава;
- осуществляет узнавание многих молекулярных сигналов (например, гормонов);
- участвует в формировании межклеточных контактов и различного рода специфических выпячиваний цитоплазмы (ресничек, жгутиков).
Механизмы проникновения веществ в клетку
Между клеткой и окружающей средой постоянно происходит обмен веществ. Ионы и небольшие молекулы транспортируются через мембрану путем пассивного или активного транспорта, макромолекулы и крупные частицы — путем эндо- и экзоцитоза.
| Способ переноса | Направление переноса | Переносимые вещества | Затраты энергии | Описание способа |
|---|---|---|---|---|
| Диффузия: через липидный слой (пассивный транспорт) | По градиенту концентрации | O2, CO2, мочевина, этанол | Без затрат энергии (пассивный процесс) | Мелкие нейтральные молекулы просачиваются между молекулами липидов. Гидрофобные вещества, как правило, диффундируют быстрее гидрофильных. Ионы и крупные молекулы не могут пересечь липидный бислой |
| Диффузия: через белковые поры (пассивный транспорт) | Ионы (в том числе Ca2+, K+, Na+), вода | Трансмембранные (интегральные) белки могут иметь водные каналы, по которым ионы или полярные молекулы пересекают мембрану, минуя гидрофобные хвосты липидов | ||
| Облегченная диффузия (пассивный транспорт) | Глюкоза, лактоза, аминокислоты, нуклеотиды, глицерин | Белок-переносчик, находящийся в клеточной мембране, на одной стороне мембраны присоединяет молекулу или ион. Это изменяет форму молекулы переносчика, и его положение в мембране изменяется так, что молекула или ион выделяются уже с другой стороны мембраны | ||
| Активный транспорт | Против градиента концентрации | Na+ и K+, H+, аминокислоты в кишечнике, Ca2+ в мышцах, Na+ и глюкоза в почках | С затратами энергии (активный процесс) | Как и облегченная диффузия, осуществляется белками-переносчиками. Но в данном случае изменение формы молекулы переносчика (ее конформация) вызывается присоединением не молекулы переносимого вещества, а фосфатной группы, отделившейся от молекулы АТФ в ходе гидролиза. |
| Фагоцитоз | Крупные макромолекулы и твердые частицы | В месте контакта с частицами мембрана впячивается, затем формируется пузырек, который отшнуровывается от плазматической мембраны и поступает в цитоплазму. Характерен для амебоидных простейших, кишечнополостных, клеток крови — лейкоцитов, клеток капилляров костного мозга, селезенки, печени, надпочечников | ||
| Пиноцитоз | Капли жидкости | Поглощение капель жидкости по механизму, аналогичному фагоцитозу. Характерен для амебоидных простейших и клеток крови — лейкоцитов, клеток печени, некоторых клеток почек |
Пассивный транспорт — перемещение веществ по градиенту концентрации; осуществляется без затрат энергии путем простой диффузии, осмоса или облегченной диффузии с помощью белков-переносчиков.
Диффузия — транспорт ионов и молекул через мембрану из области с высокой в область с низкой их концентрацией, т.е. по градиенту концентрации. Диффузия может быть простой и облегченной. Если вещества хорошо растворимы в жирах, то они проникают в клетку путем простой диффузии. Например, кислород, потребляемый клетками при дыхании, и углекислый газ в растворе быстро диффундируют через мембраны. Вода способна проходить также через мембранные поры, образованные белками, и переносить молекулы и ионы растворенных в ней веществ.
Осмос — диффузия воды через полупроницаемую мембрану из области с меньшей концентрацией солей в область с более высокой их концентрацией. Возникающее давление на полупроницаемую мембрану называют осмотическим. Клетки содержат растворы солей и других веществ, что создает определенное осмотическое давление. Живые клетки способны регулировать его, изменяя концентрацию веществ. Например, амебы имеют сократительные вакуоли для регуляции осмоса. В организме человека осмотическое давление регулируется системой органов выделения.
Облегченная диффузия — транспорт веществ в клетку через ионные каналы, образованные в мембране белками, с помощью белков-переносчиков, также находящихся в мембране. Таким образом попадают в клетку нерастворимые в жирах и не проходящие через поры вещества. Например, путем облегченной диффузии глюкоза поступает в эритроциты.
Активный транспорт — перенос веществ белками-переносчиками против градиента концентрации с затратами энергии. Например, транспорт аминокислот, глюкозы, ионов натрия, калия, кальция и др.
Эндоцитоз — поглощение веществ (путем окружения) выростами плазматической мембраны с образованием окруженных мембраной пузырьков. Экзоцитоз — выделение веществ из клетки (путем окружения) выростами плазматической мембраны с образованием окруженных мембраной пузырьков. Поглощение и выделение твердых и крупных частиц получило названия фагоцитоз и обратный фагоцитоз, жидких или растворенных частичек — пиноцитоз и обратный пиноцитоз соответственно.
Источник: jbio.ru

Рис. Схема строения клеточной стенки
1 — срединная пластинка; 2 — пора; 3 — вторичная стенка; 4 — первичная стенка
Клетки растений в отличие от клеток животных имеют твердые клеточные стенки (оболочки), которые 1) придают клетке определенную форму, 2) защищают протопласт, 3) противостоят внутриклеточному тургорному давлению и препятствуют разрыву клетки. Они, являясь внутренним скелетом растения, обеспечивают его механическую прочность. Клеточные стенки, как правило, бесцветны и легко пропускают солнечный свет. По клеточным стенкам могут передвигаться вода и растворенные в ней низкомолекулярные вещества. Стенки соседних клеток скреплены межклеточным веществом — срединной пластинкой. Срединная пластинка — единый слой, общий для двух соседних клеток. Срединная пластинка менее обводнена, в ней могут присутствовать молекулы лигнина (аморфное вещество ароматической природы, обеспечивающее одревеснение у двудольных растений). Углы клеточных стенок в результате тургорного давления округляются, и между соседними клетками образуются межклетники.
Система клеточных стенки и межклетников носит название свободного пространства — апопласта. Апопласт участвует в транспорте воды и ионов в растении. При разрушении срединной пластинки стенки соседних клеток разъединяются. Растворение межклеточного вещества, приводящее к разъединению клеток, называется мацерацией. Естественная мацерация происходит в перезрелых плодах груши, дыни, айвы и др. Искусственно ее проводят, например, примочке льна для освобождения прядильного сырья — групп клеток лубяных волокон.
Первоначально кнаружи от плазмалеммы возникает первичная клеточная стенка. Она состоит из полисахаридов — пектина и целлюлозы[1]. У двудольных растений в оболочках пектиновых веществ[2] и целлюлозы поровну, у однодольных преобладают гемицеллюлозы[3]. Целлюлоза обусловливает прочность клеточной стенки. Волоконца клетчатки — микрофибриллы — эластичны и по прочности на разрыв сходны со сталью. Полисахариды определяют такие свойства стенки, как высокая проницаемость для воды, растворенных мелких молекул и ионов, сильная набухаемость. Некоторые гемицеллюлозы могут откладываться в стенках клеток семян в качестве запасных веществ.
Клеточная стенка образуется в результате деятельности протопласта. Строительные материалы — молекулы целлюлозы, пектина, лигнина и других веществ — накапливаются и частично синтезируются в цистернах аппарата Гольджи.
При образовании первичной клеточной стенки в ней выделяются более тонкие участки, где фибриллы целлюлозы лежат более рыхло. Канальцы эндоплазматической сети проходят здесь через клеточные стенки, соединяя соседние клетки. Эти участки называются первичными поровыми полями.
К моменту, когда рост клетки заканчивается, рост клеточной стенки может продолжаться в толщину. Этот процесс носит название вторичного утолщения. Вторичная клеточная стенка откладывается на внутренней поверхности первичной клеточной стенки. Ее рост происходит в результате наложения новых мицелл целлюлоюзы (упорядоченно расположенных микрофибрилл). Таким образом, наиболее молодые слои клеточной стенки ближе всего к плазмалемме. Для некоторых типов клеток (многие волокна, трахеиды, членики сосудов) образование вторичной клеточной стенки — основная функция протопласта, после завершения вторичного утолщения он отмирает. Вторичная стенка выполняет главным образом механические, опорные функции.
Поры.Утолщается клеточная стенка неравномерно. Обычно неутолщенными остаются лишь небольшие участки первичной клеточной стенки в местах расположения первичных поровых полей — поровые каналы. Поровые каналы двух соседних клеток располагаются обычно друг против друга и разделяются замыкающей пленкой поры — двумя первичными клеточными стен-ками с межклеточным веществом между ними (срединной пластинкой). В пленке сохраняются микроскопические отверстия, через которые проходят плазмодесмы. Таким образом, пора — это два поровых канала и замыкающая пленка между ними.
Поры бывают простые и окаймленные или полукокаймленные. В простых порах диаметр порового канала по всей длине одинаковый, полость канала цилиндрическая и в поперечном сечении поры округлые. Они характерны для паренхимных клеток. В прозенхимных (очень удлиненных) клетках простые поры имеют щелевидные полости.
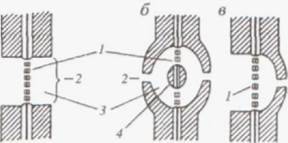
Рис. Различные пары пор (по Гуляеву, 1965):
а — простые; б — окаймленные;
в — полуокаймленные;
1 — замыкающая пленка; 2 — входное отверстие;
3 — поровый канал; 4 – торус
Окаймленные поры встречаются в стенках клеток, проводящих воду и минеральные вещества, — трахеидах и сосудах. Их поровый канал имеет форму воронки, которая своей широкой стороной прилегает к замыкающей пленке. В клетках хвойных растений замыкающая пленка окаймленных пор несет в центре дискообразное утолщение — торус, который одревесневает и становится непроницаемым для воды.
Плазмодесмы – тончайшие цитоплазматические нити или каналы, пересекающие оболочки смежных клеток, пронизывают замыкающие пленки пор. В каждой клетке имеется от нескольких сотен до десятков тысяч плазмодесм. Плазмодесмы встречаются только в растительных клетках, где имеются клеточные стенки. Плазмодесмы образуются из канальцев ЭР, которые остаются в клеточной пластинке между двумя дочерними клетками после деления.
По плазмодесмам, плазмодесменным каналам осуществляется межклеточный транспорт веществ. Объединенные плазмодесмами протопласты клеток в растении образуют единое целое — симпласт. Транспорт веществ через плазмодесмы получил название симпластического в отличие от апопластического транспорта по клеточным стенкам и межклетникам.
Видоизменения клеточной стенки.В процессе жизнедеятельности клетки целлюлозная клеточная стенка может претерпевать изменения.
Одревеснение клеточной стенки, или лигнификация, — отложение в межмицеллярные промежутки лигнина[4]. При этом возрастают твердость и прочность стенки, но уменьшается ее пластичность. Одревесневшие клеточные стенки не теряют способности пропускать воду и воздух. Протопласт их может оставаться живым, хотя обычно отмирает. Древесина хвойных и лиственных растений содержит до 20…30 % лигнина. Одревесневают клеточные стенки и многих трав, особенно к концу вегетации.
Опробковение, или суберинизация — отложение в клеточную стенку очень стойкого жироподобного аморфного вещества — суберина. Опробковсвшие клеточные стенки становятся непроницаемыми для воды и газов. К моменту завершения опробковения протопласт отмирает. Клетки с опробковевшими клеточными стенками защищают растение от испарения.
Кутинизация — отложение кутина (вещества, близкого к суберину) в поверхностных слоях наружных клеточных стенок и на их поверхности; образующаяся при этом пленка (кутикула) препятствует испарению.
Минерализация — отложение в клеточных стенках солей кальция и кремнезема (Si02). Эти вещества заполняют микрокапилляры стенки и придают ей твердость и хрупкость. Отложение кремнезема наиболее характерно для клеток эпидермы хвощей, осок и злаков. Окремнение защищает растение от улиток и слизней.
Ослизнение — превращение целлюлозы и пектина в слизи и близкие к ним камеди, представляющие собой полимерные углеводы, которые отличаются способностью к сильному набуханию при соприкосновении с водой. Ослизнение наблюдается в клеточных стенках кожуры семян, например у льна. Образование слизей имеет большое приспособительное значение. При прорастании семян слизь закрепляет их на определенном месте, легко поглощает и удерживает влагу, защищая семена от высыхания, улучшает водный режим всходов. Ослизнение клеточных стенок корневых волосков обеспечивает прочное склеивание их с частицами почвы. Иногда слизи и камеди образуются в значительных количествах при растворении клеточных стенок вследствие болезненного их состояния. (У вишни, например, часто наблюдается выделение камеди из пораненных ветвей и стволов. Камедь выделяется в виде застывающих наплывов — вишневого клея. Ослизнение такого рода называется гуммозом. Это патологическое явление).
ВАКУОЛИ –полости в протопласте, которые образуются из расширений эндоплазматической сети и пузырьков комплекса Гольджи. Вакуоли ограничены мембраной – тонопластом и заполнены клеточным соком.
В молодых делящихся растительных клетках вакуоли представляют систему канальцев и пузырьков, по мере роста клеток они увеличиваются, а затем сливаются в одну большую центральную вакуоль. Она занимает от 70 до 90 % объема клетки, в то время как протопласт располагается в виде тонкого постенного слоя.
Клеточный сок – это слабокислый (рН 3-5) водный раствор различных органических и неорганических веществ. По химическому составу и консистенции клеточный сок существенно отличается от протопласта. Эти различия связаны с избирательной проницаемостью тонопласта, выполняющего барьерную функцию.
Функции вакуолей многообразны: 1) они формируют внутреннюю водную среду клетки, с их помощью осуществляется регуляция водно-солевого обмена; 2) поддерживают тургорное давление внутриклеточной жидкости в клетке; 3) накапливают запасные и некоторые другие вещества (простые белки, углеводы, гликозиды, пигменты, алкалоиды) и 4) изолируют эргастические вещества (отбросы, конечные продукты обмена).
Тургорное давление в растительных клетках способствует 1) поддержанию формы неодревесневших частей растений, 2) также служит одним из факторов роста, обеспечивая рост клеток растяжением. Потеря тургора вызывает увядание растений. Тургорное давление связано с явлением осмоса. Осмос – это односторонняя диффузия воды через полупроницаемую перегородку в сторону водного раствора солей большей концентрации. Поступающая в клеточный сок вода оказывает давление на цитоплазму, а через нее – на стенку клетки, вызывая упругое ее состояние, обеспечивая тургор.
В состав клеточноко сока входят органические кислоты (лимонная, яблочная, щавелевая, янтарная), дубильные вещества, гликозиды, включая пигменты флавоноиды (флавоны, антоцианы), алкалоиды.
Гликозиды — эфироподобные вещества, соединения моносахаридов со спиртами, альдегидами и другими веществами. Ряд гликозидов используют в медицине: сердечные гликозиды наперстянки, ландыша, адониса, строфанта и др.; антраценовые гликозиды корней ревеня, листьев сенны, коры крушины, оказывающие слабительное действие; фенольные гликозиды листьев толокнянки, брусники при лечении воспалений почек и мочеполовых органов. К гликозидам принадлежат пигменты клеточного сока — флавоноиды. Флавоноиды окрашивают клеточный сок в лепестках цветков и плодов и тем самым способствуют привлечению насекомых-опылителей и распространению плодов. Флавоны — желтые пигменты. Антоцианы — пигменты, меняющие свою окраску в зависимости от реакции клеточного сока от красной до синей. В нейтральной среде антоцианы имеют лиловатую окраску, в щелочной — синюю, в кислой — красную. Красный цвет у антоцианов в цветках пионов, гераней, маков, роз; синий — в цветках дельфиниумов, васильков; малиново-лиловый — и плодах слив, винограда. Антоцианы окрашивают осенние листья в ярко-красный цвет. Они образуются в холодную солнечную погоду и проявляются по мере разрушения хлорофилла. Наиболее ярко окрашены листья холодной ясной осенью.
Дубильные вещества — эфиры фруктозы и ароматических кислот, предохраняющие растения от загнивания. Соединяясь с белками, они дают нерастворимые соединения, поэтому широко применяются для дубления кож. После дубления кожа делается мягкой, прочной и не пропускает воду. Больше всего дубильных веществ содержит дуб (в его коре 10…20 %), чай (в листьях 15…20 %), лиственница, бадан. Благодаря вяжущему и противовоспалительному действию дубильные вещества используют при лечении желудочно-кишечных расстройств, ожогов, кожных и других болезней человека.
Алкалоиды — органические основания, содержащие азот. В растениях находятся в виде солей органических кислот, они жгучие и ядовитые, что делает их косвенно полезными для растений. Как правило, проявляют высокую физиологическую активность и оказывают сильное влияние на организм человека и животных. Широко используют в качестве лекарств разнообразного действия: наркотики, транквилизаторы, болеутоляющие средства и др. (хинин, атропин, кофеин, эфедрин, пилокарпин, стрихнин, морфин) и инсектицидных (против насекомых) средств (никотин, анабазин).
ВКЛЮЧЕНИЯ— это непостоянные компоненты цитоплазмы, содержание которых меняется в зависимости от функционального состояния клетки. Различают трофические и экскреторные включения. Трофические включения представляют собой запасы питательных веществ. В растительных клетках это крахмальные и белковые зерна, липидные (жировые) капли. Экскреторные включения представляют собой продукты обмена веществ в растительных и животных клетках (например, кристаллы щавелевой кислоты, щавелевокислого кальция в виде одиночных многогранников, стилоидов, рафид, друз; кристаллы карбоната кальция или кремнезема в виде цистолитов). Форма кристаллов нередко специфична для разных таксонов и служит для микродиагностики.
Источник: helpiks.org
Строение
Клеточная оболочка состоит из 3 слоев (Рис.1):
1) наружный (надмембранный) слой — гликокаликс (Glicocalyx);
2) собственно мембрана (биологическая мембрана);
3) подмембранная пластинка (кортикальный слой плазмолеммы).
Гликокаликс — образован ассоциированным с плазмолеммой гликопротеидным и гликолипидным комплексами, в состав которых входят различные углеводы. Углеводы представлены длинными, ветвящимися цепочками полисахаридов, которые связаны с белками и липидами, находящимися в составе плазмолеммы. Толщина гликокаликса 3-4 нм, он присущ практически всем клеткам животного происхождения, но с разной степенью выраженности. Полисахаридные цепочки гликокаликса являются своеобразным аппаратом, при помощи которого происходит взаимоузнавание клеток и их взаимодействие с микроокружением.
Собственно мембрана (биологическая мембрана). Структурная организация биологической мембраны наиболее полно отражена в жидкостно — мозаичной модели Сингер-Никольского, согласно которой молекулы фосфолипидов контактируют своими гидрофобными концами (хвостами), и отталкиваясь гидрофильными концами (головками), образуют сплошной двойной слой.
В билипидный слой погружены полностью интегральные белки (это преимущественно гликопротеиды), полуинтегральные белки погружены частично. Эти две группы белков в билипидном слое мембраны располагаются таким образом, что их неполярные части входят в этот слой мембраны в местах локализации гидрофобных участков липидов (хвосты). Полярная часть молекулы белка взаимодействует с головками липидов обращена в сторону водной фазы.
Кроме этого часть белков расположена на поверхности билипидного слоя, это так называемые примембранные или периферические или адсорбированные белки.
Положение белковых молекул не является жестко лимитированным и в зависимости от функционального состояния клетки может происходить их взаимное перемещение в плоскости билипидного слоя.
Такая изменчивость положения белков, и сходная с мозаикой, топография микромолекулярных комплексов поверхности клетки, дала название жидкостно-мозаичной модели биологической мембраны.
Лабильность (подвижность) структур плазмолеммы зависит от содержания в ее составе молекул холестерина. Чем больше содержится холестерина в составе мембраны, тем легче происходит перемещение макромолекулярных белков в билипидном слое. Толщина биологической мембраны 5-7 нм.
Подмембранная пластинка (кортикальный слой) образована наиболее плотной частью цитоплазмы, богатой микрофилламентами и микротрубочками, которая образует высокоорганизованную сетку, при участии которой происходит перемещение интегральных белков плазмолеммы, обеспечивается цитоскелетная и локомоторная функции клетки, реализуются процессы экзоцитоза. Толщина этого слоя составляет около 1нм.
Функции
К основным функциям, выполняемым клеточной оболочкой относятся следующие:
1) разграничение;
2) транспорта веществ;
3) рецепция;
4) обеспечение межклеточных контактов.
Разграничение и транспорт метаболитов
Благодаря разграничению с окружающей средой клетка сохраняет своя свою индивидуальность, благодаря транспорту клетка может жить и функционировать. Обе эти функции являются взаимоисключающими и взаимодополняющими друг друга и оба процесса направлены на поддержку постоянства характеристик внутренней среды — гомеостаза клетки.
Транспорт из внешней среды внутрь клетки может быть активным и пассивным.
·Путем активного транспорта осуществляется перенос многих органических соединений против градиента плотности с затратой энергии за счет расщепления АТФ, с участием ферментных транспортных систем.
·Пассивный транспорт осуществляется путем диффузии и обеспечивает перенос воды, ионов, некоторых низкомолекулярных соединений.
Транспорт веществ из внешней среды внутрь клетки называется эндоцитозом, процесс выведения веществ из клетки носит название экзоцитоза.
Эндоцитоз делят на фагоцитоз и пиноцитоз.
Фагоцитоз — это захват и поглощение клеткой крупных частиц (бактерий, фрагментов других клеток).
Пиноцитоз — это захват микромолекулярных соединений, которые находятся в растворенном состоянии (жидкостей).
·Эндоцитоз протекает в несколько следующих друг за другом этапов:
-
Сорбция — поверхностью мембраны поглощаемых веществ, связывание которых с плазмолеммой определяется наличием на ее поверхность рецепторных молекул.
-
Образование впячиваний плазмолеммы внутрь клетки. Вначале впячивания имеют вид незамкнутых округлых пузырьков или глубоких инвагинаций.
-
Отшнуровывание впячиваний от плазмолеммы. Отделившиеся пузырьки свободно размещаются в цитоплазме под плазмолеммой. Пузырьки могут сливаться друг с другом.
-
Расщепление поглощенных частиц при помощи гидролитических ферментов, поступающих из лизосом.
Иногда встречается и такой вариант, когда частичка поглощается одной поверхностью клетки и в окружение биомембраны проходит через цитоплазму и выводится из клетки без изменений на противоположной поверхности клетки. Такое явление называется цитопемписом.
Экзоцитоз — это выведение продуктов жизнедеятельности клетки за пределы цитоплазмы.
Существует несколько разновидностей экзоцитоза:
1) секреция;
2) экскреция;
3) рекреция;
4) клазматоз.
Секреция — выделение клеткой продуктов ее синтетической деятельности, необходимых для обеспечения физиологических функций органов и систем организма.
Экскреция — выделение токсических продуктов метаболизма, которые подлежат выведению за пределы организма.
Рекреция — удаление из клетки соединений, которые не меняют своей химической структуры в процессе внутриклеточного метаболизма (вода, минеральные соли).
Клазматоз — удаление за пределы клетки отдельных ее структурных компонентов.
·Экзоцитоз состоит из ряда последовательных стадий:
-
накопление продуктов синтетической деятельности клетки в виде окруженных биомембранной скоплений в составе мешочков и пузырьков комплекса Гольджи;
-
перемещение этих скоплений из центральных участков цитоплазмы к периферии;
3) включение биомембраны мешочка в плазмолемму;
4) эвакуация содержимого мешочка в межклеточное пространство.
Источник: StudFiles.net
Наличие клеточной оболочки более чем все другие признаки отличает растения от животных. Например, простейшие жгутиковые, покрытые оболочкой, причисляют к примитивным растениям (Protophyta), а голые жгутиковые к примитивным животным (Protozoa). У низших растений голыми являются лишь репродуктивные клетки, а клетки, составляющие вегетативное тело, имеют клеточные стенки. У высших растений клеточной стенкой обладают даже гаметы: как яйцеклетка, так и пыльцевая трубка.
Клеточная оболочка окружает собственно клетку со всех сторон и служит связующим звеном между ней и соседними клетками, обеспечивая, таким образом, единство и целостность всего растительного организма. В жестких оболочках растительных клеток образуются каналы, в которых располагаются тончайшие тяжи цитоплазмы — плазмодесмы. Благодаря этому, осуществляются межклеточные взаимодействия. Иными словами, у растений клеточные оболочки призваны обеспечивать те функции, которые у животных выполняют скелет, кожа и система кровообращения (т.е. опорную, защитную и транспортную.) Не удивительно поэтому, что в ходе эволюции у растений возникли весьма разнообразные по структуре и химическому составу типы клеточных стенок. Собственно говоря, растительные клетки во многом различают и классифицируют именно по форме и природе клеточных стенок.
Оболочка, как правило, бесцветна и прозрачна. Она легко пропускает солнечный свет. Оболочки соседних клеток как бы сцементированы межклеточными веществами, образующими так называемую срединную пластинку. Вследствие этого соседние клетки оказываются отделёнными друг от друга стенкой, образованной двумя оболочками и срединной пластинкой. Это и даёт основание называть оболочку также клеточной стенкой.
Клеточная стенка растительных клеток состоит, главным образом, из полисахаридов. Оболочку эмбриональных тканей и клеток, растущих растяжением, называют первичной. В этот период оболочка достаточно эластична. После прекращения роста клетки изнутри на первичную клеточную стенку начинают откладываться новые слои и образуется вторичная клеточная стенка, придающая клетке жёсткость и прочность.
Химический состав
Все компоненты, входящие в состав клеточной стенки, можно разделить на 4 группы:
- Структурные компоненты, представленные целлюлозой у большинства автотрофных растений, хитином (грибы), глюканом (дрожжи), манналом и ксиланом (водоросли).
- Компоненты матрикса, т.е. основного вещества, наполнителя оболочки — гемицеллюлозы, белки, липиды.
- Компоненты, инкрустирующие клеточную стенку, (т.е. откладывающиеся и выстилающие её изнутри) — лигнин и суберин.
- Компоненты, адкрустирующие стенку, т.е. откладывающиеся на её поверхности, — кутин, воск.
Основной структурный компонент оболочки — целлюлоза представлена длинными неразветвленными полимерными молекулами, состоящими из 1000-11000 остатков b -D глюкозы, соединённых между собой гликозидными связями. Наличие гликозидных связей создаёт возможность образования поперечных стивок.
Благодаря этому, длинные и тонкие молекулы целлюлозы объединяются в элементарные фибриллы или мицеллы. Каждая мицелла состоит из 60-100 параллельно расположенных цепей целлюлозы.
Мицеллы сотнями группируются в мицеллярные ряды и составляют микрофибриллы диаметром 10-25 нм. Целлюлоза обладает кристаллическими свойствами благодаря упорядоченному расположению мицелл в микрофибриллах.
Микрофибриллы, в свою очередь, перевиваются между собой как пряди в канате и объединяются в макрофибриллы. Макрофибриллы имеют толщину около 0,5 мкм и могут достигать в длину 4 мкм. Они так же прочны, как равная им по толщине стальная проволока.
Целлюлоза относится к индиферентным веществам: она не обладает ни кислыми, ни щелочными свойствами. По отношению к тепловым воздействиям она достаточно стойка и может быть нагрета без разложения до температуры 200 градусов.
Многие из важных свойств целлюлозы обусловлены её высокой стойкостью отношению к ферментам и химическим реагентам. Она не растворима в воде, в спирте, в эфире и в других нейтральных растворителях; не растворяется и в кислотах и щелочах.
Если целлюлоза и растворяется при известных условиях под действием и щелочей, то это растворение сопровождается сильным разрушением её организации. Без разрушения мицелл целлюлоза может быть растворена в реактиве Швейцера (аммиачном растворе окиси меди), при этом целлюлоза превращается в гель с беспорядочным расположением молекул. Такой гель более гигроскопичен и легче окрашивается, чем природная целлюлоза.
При кратковременной и осторожной обработке целлюлозы серной кислотой образуется амилоид, представляющий по некоторым свойствам нечто промежуточное между клетчаткой и крахмалом: по консистенции амилоид подобен клейстеру. Приготовление «пергаментной бумаги», не пропускающей масла, основано именно на этом принципе. Листы бумаги погружаются на очень короткое время в серную кислоту, затем отмываются и высушиваются, при этом тонкий верхний слой бумаги превращается в непромокаемый для жиров амилоид.
Действием азотной кислоты могут быть получены нитроцеллюлозы. Нитроцеллюлозы находят обширное применение в технике: они используются для изготовления шёлка, пороха, лаков, красок пластмасс.
Целлюлоза, пожалуй, самый распространенный вид органических макромолекул на Земле. Она составляет основу питания многих гетеротрофных организмов. Целлюлоза разрушается действием определенных ферментов, содержащихся в теле некоторых низших растений, грибов, животных. Например, грибы — домовой гриб и трутовики, используя клетчатку живых деревьев или построек, разрушают целлюлозу. Термиты способны переваривать целлюлозу, благодаря наличию в их пищеварительном тракте простейших, энзимы которых перерабатывают клетчатку в вещества, усваиваемые термитами. В пищеварительном аппарате крупных травоядных животных содержится целая микрофлора организмов, подготавливающих клетчатку к усвоению.
Микрофибриллы оболочки погружены в амморфный пластичный гель — матрикс. Матрикс является наполнителем оболочки. В состав матрикса входят полисахариды, называемые гемицеллюлозами и пектиновыми веществами. На долю гемицеллюлоз приходится около 30-40 % сухого веса клетчатых стенок.
По строению гемицеллюлозы напоминают целлюлозу и крахмал (это тоже полимеры); их цепи состоят из остатков гексоз либо пентоз, связанных кислородными мостиками. Но если молекулы целлюлозы насчитывают в своём составе от 1000 до 10000 мономеров, то цепочки гемицеллюлоз состоят из 150-300 молекул мономеров. Они значительно короче. Именно поэтому гемицеллюлозы нередко называют полуклетчатками. Основными гексозами гемицеллюлоз являются D-глюкоза и D-галактоза; пентозами — L-ксилоза и L-арабиноза.
По отношению и химическим агентам гемицеллюлозы гораздо менее стойки, чем целлюлоза: они растворяются в слабых щелочах без подогревания; гидролизуются с образованием сахаров в слабых растворах кислот; растворяются полуклетчатки и в глицерине при температуре 300 градусов.
Гемицеллюлозы в теле растений играют механическую роль, участвуя наряду с целлюлозой и другими веществами в построении клетчатых стенок.
роль запасных веществ, отлагающихся, а затем расходующихся. При этом функцию запасного материала несут преимущественно гексозы; а гемицеллюлозы с механической функцией обычно состоят из пектоз. В качестве запасных питательных веществ гемицеллюлозы отлагаются также в семенах многих растений, особенно у однодольных, например, в клеточных стенках эндосперма многих пальм (в частности финиковой пальмы).
Пектиновые вещества имеют довольно сложный химический состав и строение. Химики определяют их, как кальциевые-магниевые соли полимерной a -D галактуроновой кислоты, соединённые a -1,4-связями, т.е. мономером является галактуроновая кислота. Характерная особенность: пектиновые вещества сильно набухают в воде, а некоторые даже в ней растворяются. Легко они разрушаются и под действием щелочей и кислот.
Все клеточные стенки на ранней стадии развития почти нацело состоят из пектиновых веществ.
Межклеточное вещество срединной пластинки, как бы цементирующее оболочки смежных стенок, состоит обычно из пектиновых веществ, главным образом из пектата кальция. Пектиновые вещества, хотя и в небольших количествах, имеются в основной толщине и взрослых клеток.
В состав матрикса клеточных стенок помимо углеводных компонентов входит также структурный белок, называемый экстенсином. Он является гликопротеином, углеводная часть которого представлена остатками сахара арабинозы.
Одревеснение клеточных оболочек
Сильному метаморфозу состава и структуры подвергаются оболочки при одревеснении, опробковении, ослизнении и минерализации. Одревеснение состоит в том, что часть целлюлозной толщи стенки пропитывается лигнином.
Ароматическое вещество лигнин является основным инкрустирующим веществом клеточной стенки. Это полимер с неразветлённой молекулой, состоящей из ароматических компонентов. Мономерами лигнина могут быть конифериловый, синаповый и другие спирты.
Интесивная лигнификация клеточных стенок начинается после прекращения роста клетки.
Отношение между целлюлозой и лигнином в одревесневших слоях оболочки было признано аналогичным конструкции железобетонных сооружений. Лигнин, подобно бетонной массе, заполняет промежутки ячеек сетки; при этом арматура и заполнение образуют монолитное целое.
Одревеснение широко распространено в растительном царстве, за исключением низших растений и мохообразных.
У папоротникообразных одревеснение — явление довольно обычное; а у голосеменных и покрытосеменных распространено повсеместно; здесь одревеснение встречается во всех органах и в разнообразных тканях.
Понятно, что особенно сильному одревеснению подвержены клетки деревьев и кустарников. Так, древесина хвойных содержит 27-30 % лигнина, а древесина лиственных — 18-27 %.
Одревеснение понижает пластичность клеточных стенок, закрепляет их форму. Однако клетки с одревесневшими стенками могут оставаться живыми десятки лет.
Лигнин обладает и консервирующими свойствами и потому действует как антисептик, придавая тканям повышенную стойкость по отношению к разрушительному действию грибов и бактерий.
При разложении в почве растительных остатков относительное содержание лигнина в них повышается. Несомненно, высокая стойкость лигнина играет большую роль при торфо- и углеобразовании. Кроме того, разрушение и конденсация лигнина в почве служит одним из факторов образования гумуса.
Опробковение и кутинизация клеточных оболочек
Весьма распространено в растительном мире наличие в толще клеточных оболочек, либо на их поверхности веществ, называемых кутинами, суберинами и спорополленинами. Общим для них являются следующие черты. Все они — высокополимерные вещества, обязательным компонентом которых являются насыщенные и ненасыщенные, жирные кислоты и жиры. От жиров, встречающихся в полости клетки, в протопласте, они отличаются нерастворимостью в ряде реактивов. Эти вещества стойки даже по отношению к концентрированной серной кислоте. Суберины, кутипы и спорополленины почти непроницаемы для воды, воздуха и других газов.
Эти свойства объясняют нам значение суберинов, кутипов и спорополленинов для растений и их локализацию в растениях. Они находятся в оболочках перефирических (покровных) тканей и защищают органы растения от излишней потери воды.
Суберины. Клеточные оболочки, содержащие суберины, называют опробковевшими. Суберин отлагается внутри клеточной оболочки и поэтому относится к инкрустирующим веществам. Обычно суберин составляет пластинку, находящуюся в так называемом вторичном слое клеточной стенки, так называемые субериновые пластинки.
Кутины — это адкрустирующие гидрофобные вещества, покрывающие поверхность эпидермальных клеток растенеия в виде пленки — кутикулы.
Обычно кутикулярная пленка многослойна.
У папоротников кутикула отсутствует или слабо развита; не выражена кутикула и в корнях растений.
Спорополленины имеются в наружных оболочках спор, в том числе пыльцевых зерён голосеменных и покрытосеменных растений.
Спорополленины настолько стойки к разрушающим факторам, что пыльцевые зёрна со всеми тонкостями структуры их оболочек прекрасно и надолго сохраняются в почве и в торфяных отложениях и могут быть использованы для особого споро-пыльцевого анализа, позволяющего восстановить картину растительности былых эпох.
Ослизнение клеточных оболочек
При ослизнении клеточных оболочек образуются слизи и камеди. Те и другие представляют собой высокомолекулярные углеводы, состоящие большей частью из пентоз и их производных. Они нерастворимы в спирте, эфире, а в воде сильно набухают.
Резкой границы между ними не установлено. Обычно их различают по консистенции в набухшем состоянии: камеди клейки и могут вытягиваться в нити, слизи же сильно расплываются и в нити не тянутся.
В сухом состоянии камеди и слизи очень тверды и хрупки, и лишь при смачивании водой они переходят в тягучее желеобразное состояние.
Ослизение бывает или нормальным явлением в жизни растения, или же происходит как патологический процесс.
В некоторых случаях слизи и камеди появляются в клеточных оболочках путём отложения на ранее образовавшиеся слои клеток, иногда они образуются из имеющегося уже материала стенок в результате химического метаморфоза — «слизистого перерождения».
При патологическом ослизнении — камедетечении (или гуммозе) — в камедь превращаются и стенки, и содержание клеток; процесс этот захватывает большие участки тканей. Гуммоз часто наблюдается у вишен, слив, персиков, у многих акаций и астрагалов.
Значение ослизнения клеточных стенок во многих случаях очевидно. Например, ослизненные наружные слои клеток кожицы семян, набухая весной, входят в соприкосновение с почвой. Слизь, благодаря клейкости, закрепляет семена на влажном месте и, поглощая воду из почвы, улучшает водный режим проростка, передавая ему воду и защищая от высыхания.
Слизь может использоваться как запасное, питательное вещество.
Минерализация клеточных оболочек
В более поздней стадии развития оболочки содержат минеральные вещества, причём в некоторых случаях в весьма значительных количествах. Эти вещества могут отлагаться и в толще оболочки и на её внутренней и наружной поверхности, или же в особых выростах клеточных стенок. По структуре эти отложения могут быть аморфными и кристаллическими.
Наиболее распространены отложения кремнезёма и солей извести.
Богаты кремнезёмом клетки кожицы стеблей и листьев хвощей, злаков, осок. Окремнение стенок свойственно и многим растениям из двудольных, особенно из семейства мареновых. Окремнению подвергаются жгучие волоски у крапивы двудольной.
Кальций встречается в клеточных оболочках в виде углекислой, щавелевокислой и пектиновокислой извести.
Широко распространено наличие кальция в срединной пластинке клеточных стенок.
Структурная организация клеточной оболочки
Клеточная стенка, как мы видели, построена из немногих основных компонентов. Применение химических методов анализа позволило выявить, что:
соседние цепи целлюлозы в микро — и макрофибриллах связаны водородными связями;
молекулы гемицеллюлозы прикреплены к поверхности целлюлозных микрофибрилл также водородными связями;
некоторые молекулы гемицеллюлозы связаны с молекулами кислого пектина через молекулы нейтрального пектина;
сами пектиновые полимеры сшиваются между собой ионами кальция (Ca);
гликопротеины вероятно присоединены к молекулам пектина; и, наконец,
существуют ковалентные связи между лигнином и целлюлозой.
Таким образом, согласно этой модели, клеточную стенку можно рассматривать как единую гигантскую макромолекулу.
Формирование и рост клеточных оболочек
Даже молодые меристематические клетки снабжены первичной оболочкой. Новая оболочка формируется в процессе деления клетки в заключительной стадии митоза — телофазе. Заключительная в телофазе стадия — процесс деления цитоплазмы на дочерние клетки называется цитокинезом.
В ранней телофазе между двумя дочерними ядрами формируется бочкообразная система волокон, называемая фрагмопластом. Волокна фрагмопласта, так же как и волокна митотического веретена состоят из микротрубочек. В световом микроскопе видно, как в экваториальной плоскости фрагмопласта появляются мелкие капли, которые затем сливаются, образуя клеточную пластинку. Клеточная пластинка растёт центробежно до тех пор, пока не достигнет оболочки делящейся клетки. С помощью электронного микроскопа было установлено, что сливающиеся капельки — это пузырьки, отрывающиеся от аппарата Гольджи. Они в основном содержат пектиновые вещества, из которых формируется срединная пластинка, а мембраны пузырьков участвуют в построении плазматической мембраны по обеим сторонам пластинки. В это время из фрагментов трубчатого эндоплазматического ретикулума образуются плазмодесмы.
После образования срединной пластинки протопласт дочерних клеток откладывает на нее первичную оболочку.
Слой целлюлозы, который откладывается во время роста клетки, называется первичной клеточной оболочкой. Помимо целлюлозы, гемицеллюлоз и пектина первичные оболочки содержат и структурный белок — гликопротеин. Первичные оболочки могут и лигнифицироваться, хотя, как правило, лигнин им не свойственен.
Однако наиболее характерную часть первичной оболочки составляет пектиновый компонент. Он придаёт оболочке пластичность, позволяет ей растягиваться, по мере удлинения органов: корня, стебля, листа.
Пектиновые вещества способны сильно набухать, поэтому первичные оболочки содержат много воды (60-90%).
В целом, на долю гемицеллюлоз и пектиновых веществ, приходится 50-60 % сухого веса первичной оболочки, содержание целлюлозы не превышает 30 %, структурный белок занимает до 10%, лигнин, как правило, отсутствует.
Вторичное утолщение оболочек обычно начинается после прекращения роста клеток. При этом толщина оболочки увеличивается, а объём, запасаемый полостью клетки, сокращается.
Во многих случаях по завершении вторичного утолщения протопласты клеток отмирают, но клетки продолжают функционировать, выполняя главным образом механическую и опорную функцию.
В соответствие с механической функцией строение и химический состав вторичной оболочки сильно отличается от первичной. В ней значительно меньше воды и преобладают плотно сомкнутые микрофибриллы целлюлозы (40-50 %), в первичной оболочке — они расположены рыхло. Много во вторичных оболочках и лигнина — 25-30%, он придаёт оболочкам дополнительную жёсткость и прочность; гемицеллюлозы составлют 20-30% и практически нет пектиновых веществ.
Итак: лицо первичной оболочки составляют пектиновые вещества, в то время как вторичной — лигнин.
Вторичная оболочка далеко не всегда откладывается равномерно. У некоторых специализированных водопроводящих клеток она имеет вид колец или спиральных лент. Такие клетки сохраняют способность к продольному растяжению и после отмирания.
Плазмодесмы
Протопласты соседних клеток связаны между собой тонкими нитями цитоплазмы — плазмодесмами. Эти структуры присущи только растительным клеткам.
В нормальном состоянии плазмодесмы невидимы в световой микроскоп, однако, если стимулировать набухание оболочки плазмодесмы, становятся заметными, поэтому выявлены и описаны они были уже достаточно давно. Хотя детали строения плазмодесм изучены сравнительно недавно с помощью электронного микроскопа.
Под электронным микроскопом плазмодесмы выглядят как узкие каналы (диаметром от 30 до 60 нм), выстланные плазматической мембраной. По оси канала из одной клетки в другую тянется цилиндрическая трубочка меньшего размера — десмотрубочка, которая сообщается с эндоплазматическим ретикулом обеих смежных клеток.
Десмотрубка напоминает цитоплазматические микротрубочки или жгутики простейших. Она состоит из 11 спирально расположенных белковых субъедениц.
Вокруг десмотрубки локализируется цитоплазма, которая во многих типах плазмодесм непосредственно не соединяется с цитоплазмой клеток.
В плазмодесмах обнаружена АТФ — азная активность.
Наличие плазмодесм обеспечивает непрерывность цитоплазмы клеток, составляющих органы и ткани. Такая непрерывная система называется симпласт.
Кроме того, за счёт плазмодесм обеспечивается единство эндоплазматической сети, переходящей из клетки в клетку. Единая эндоплазматическая сеть получила название эндопласт.
Таким образом, выделяется три непрерывных компартмента в растительных тканях — это:
- единая цитоплазма — симпласт,
- непрерывный ретикулум — эндопласт и соприкасающиеся между собой клеточные стенки, вместе с межклетниками составляют непрерывную систему — апопласт.
Размещаются плазмодесмы в стенке либо группами, либо равномерно разбросаны по всей стенке.
Поры
Внутренне утолщение клеточной стенки не бывает вполне равномерным. Сформировавшаяся оболочка имеет более толстые и менее утолщенные участки.
Даже в тех случаях, когда стенки, в общем, имеют равномерную толщину, в них, при детальном рассмотрении обнаруживаются небольшие углубления. Эти места, в которых оболочка очень тонка, и называются порами.
Таким образом, поры у растений — это не сквозные многочисленные отверстия, как это понимается в общеупотребительном смысле. У растений порой называют любое неутолщенное место оболочки.
Для обозначения сквозных отверстий у растений используется другое название — перфорации.
Поры в 2-х сосединх клетках располагаются одна против другой, образуя так называемую пару пор.
У клеток с мощной вторичной оболочкой поры в разрезе имеют вид радиальных каналов. На поперечном срезе эти каналы могут иметь разную форму: чаще округлую, реже щелевидную (эллиптическую или крестообразную). Округлые поры обычно формируются в паренхимных клетках, щелевидные — в прозенхимных.
По форме порового канала обычно различают поры 2-х типов: простые и окаймлённые.
Простые поры имеют достаточно ровный канал, с одинаковым диаметром на всём протяжении.
У окаймлённых пор голосеменных растений на первичной оболочке образуется линзовидное утолщение — торус, а выросты вторичных оболочек как бы нависают над торусом.
Окаймлённые поры характерны для водопроводящих элементов древесины. Эти элементы имеют вид длинных труб разного диаметра. По этим трубкам, как по капиллярам, поднимается вода. Понятно, что давление воды в смежных клетках неодинаково. В этом случае торус смещается и прижимается к выступам вторичной оболочки клеток с меньшим давлением.
Обычно к порам приурочены и плазмодесменные канальцы. Нередко через одну пору проходят десятки плазмодесм.
В любом случае, поры, как и плазмодесмы, облегчают диффузию веществ, растворённых в воде, из одной клетки в другую.
Сквозные отверстия клеточных стенок — перфорации особенно характерны для водопроводящих поперечных перегородок водопроводящих члеников сосудов. Как правило, в этих перегородках образуются одна, две или несколько крупных перфораций.
Многочисленные мелкие перфорации имеются в так называемых ситовидных трубках, по этим трубкам также передвигается вода с органическими веществами, но сверху вниз, от листьев к корням.
Функции и эволюция клеточной стенки
Являясь продуктом метаболической деятельности протопласта клеточная стенка выполняет ряд функций:
- Она защищает клеточное содержимое от повреждений и инфекций (защитная функция);
- Клеточная стенка поддерживает форму и определяет размер клетки; стенка играет скелетную (опорную) роль, которая особенно возрастает у наземных растений;
- Она имеет большое значение в росте и дифференцировании клетки;
- Стенка участвует в ионном обмене и поглощении клеткой веществ;
- Единый апопласт способствует перемещению веществ из клетки в клетку внеклеточным путём (проводящая функция);
- Структура клеточных стенок предохраняет клетки от избыточной потери воды (покровная функция).
Примитивные клетки были окружены слизистым чехлом, состоявшим из пектиновых веществ, как и фрагмопласт, возникающий при митотическом делении в клетках современных растений.
Совершенствование защитной функции клеточной оболочки привело к появлению в её составе гемицеллюлозных компонентов. Форма клетки могла поддерживаться кремниевым и карбонатным наружным чехлом, сохранившимся, как вы знаете из предыдущего курса, у некоторых водорослей. По предположению Фрей-Висслинга первичный слизистый углеводный чехол мог быть предшественником матрикса клеточной стенки.
С возникновением автотрофного способа питания в оболочках клеток в качестве структурного компонента появилась целлюлоза. Выход растений на сушу поставил клеточную стенку перед необходимостью выполнять функцию опоры тела в воздухе. Именно целлюлоза оказалась наиболее оптимальным материалом (в меру прочным и в то же время эластичным) в динамичной и переменчивой среде, где подземным органам растений пришлось испытывать более сильные нагрузки.
Выход на сушу и увеличение размеров растительных организмов привели также к необходимости снабжения клеток водой. Именно с развитием у наземных растений сосудов, проводящих воду, связывают появление в клеточных стенках лигнина. Лигнин не обнаружен у ископаемых океанских и современных водных растений.
Источник: www.AgroJour.ru