Клеточный цикл определение
| |
Предмет генетики.
Генетика-раздел биологии, изучающий материальные основы наследственности и изменчивости, а также закономерности наследования и изменения признаков в ряду поколений.
Наследственность— это свойство живых организмов передавать дочернему поколению способность к определённому типу индивидуального развития, в ходе которого у потомства возникают признаки, присущие особям данного вида, а также воспроизводятся некоторые индивидуальные особенности родителей.
Наследственность (по Дарвину)— это свойство живых организмов передавать свои признаки и особенности развития следующим поколениям.
Благодаря наследственности организмы в пределах вида схожи между собой ( из семян пшеницы вырастает пшеница, у человека рождается человек и т.д.).
Изменчивость-— это свойство живых организмов приобретать новые признаки в процессе индивидуального развития.
Благодаря изменчивости особи в пределах вида отличаются друг от друга. Дети одних родителей при значительном сходстве могут существенно отличаться друг от друга по признакам (полу, росту, группам крови, цвету глаз и тд.).
Следовательно, наследственность и изменчивость -2 противоположных, но взаимосвязанных свойства организма. Благодаря единству этих свойств ,виды с одной стороны , остаются неизменными на протяжении долгого времени, а с другой- способны изменяться, давая новые формы, виды.
2. Медицинская генетика.
Наследственность человека изучает антропогенетика. Её составной частью является медицинская генетика.Она изучает:
· Закономерности наследственности и изменчивости человека под углом зрения патологии, а именно причины возникновения наследственных болезней человека, характер наследования в семьях, распространение. Особым разделом является клиническая генетика, исследующая вопросы патогенеза, клиники. Диагностики, профилактики и лечения наследственных заболеваний.
· Виды изменчивости, в частности хромосомные, геномные, генные мутации, условия их возникновения, профилактику мутагенных факторов.
· Изучение новых медико-биологических проблем: вопросы генной инженерии, которая разрабатывает методы лечения наследственных болезней, клонирование, несовместимость матери и плода, трансплантацию органов.
Генетика расширяет и углубляет биологическое мышление специалиста. Генетические знания необходимы для понимания новых методов диагностики, лечения и профилактики наследственных болезней. Наследственные заболевания занимают существенное место в работе врача и медицинской сестры, т.к. они часты и тяжелы. По разным источникам насчитывается от 5000-8000 наследственных болезней, которые поражают все органы, системы и функции организма. Около 5% детей рождается с наследственными и врождёнными болезнями. Не менее 25% больничных коек заняты пациентами, страдающими заболеваниями с наследственной предрасположенностью ( гипертония, ишемия сердца, псориаз, бронхиальная астма, глаукома, рассеянный склероз).
Наследственная патология может «преследовать» человека от зарождения до старости. Следовательно, мед. помощь и психологическая поддержка должны оказываться не только больному, но и его семье. В этом велика роль медсестры. Генетика ставит задачу избавить человечество от наследственных дефектов. Сохранение наследственной информации в ряду поколений внутри каждого вида осуществляется благодаря способности клеток (из которых построено тело) к делению. Поэтому необходимо познакомиться с цитологией – наукой о клетке.
Клетка.
Клетка — элементарная единица жизни. Тело всех многоклеточных организмов на Земле: растений, животных, человека состоит из клеток.
Одноклеточные представляют собой единственную клетку.
Открыл клетку (да и сам термин «клетка» – ячейка) английский физик Роберт Гук, в 1665 году -в срезе пробки, 1680 Левенчук — животную клетку. Шлейден — ботаник и Шванн — зоолог в 1838-1839 году сформулировали клеточную теорию.
1) Клетка – элементарная единица живого. Она обладает всеми свойствами живого — осуществляет обмен веществ и энергии, размножается, растет, для неё характерна раздражимость.
2) Клетка разных животных живых органов имеет общий план строения.
3) Образование новых клеток возможно только путём деления уже существующей клетки.
4) Клетка – это функциональная единица в многоклеточном организме.
Организмы делятся на:доклеточные (вирусы) и клеточные.
По строению клеток организмы делятся на:
прокариоты- доядерные (бактерии, сине-зелёные водоросли) – нет сформированного ядра.
эукариоты— собственноядерные организмы, имеют ядро (растения, грибы, животные)
Мы будем изучать эукариотические клетки — животные клетки. Клетки имеют разнообразную форму: звездчатая (нерв.), призматическая (эпит), овальная (эритроц), веретеновидная, шаровидная (яйцеклетка) и др.
Размеры клеток так же варьируют от нескольких микрон до 100 мкр. Наиболее крупными являются яйцеклетки- птиц (желток) до 1см и более. В основном от 0,5-40 мкр.
В организме многоклеточных различают соматические клетки- клетки тела (сома- тело) и генеративные клетки (qenero- рожаю)- половые клетки (гаметы).
Строение клетки.
 КЛЕТКА
КЛЕТКА
ПЛАЗМОЛЕММА ЦИТОПЛАЗМА ЯДРО
гиалоплазма органеллы включ ения
1.троф. 1Яд. оболочка
мембранные немембранные 2.белков 2.Кариопл.
1)митохондрии 1) рибосомы 3.ядроген
2)Э.П.С. 2) микротр. 4.хроматин
3)Ком Г 3) центриоли
4)лизосом 4) миофибриллы
5)пластиды 5) ресничек
Содержимое клетки отделено от внешней среды или от соседних клеток плазматической мембраной (плазмолеммой). Клетка состоит из 2х основных компонентов: цитоплазмы и ядра. Органоиды так же делятся на: общего назначения (во всех клетках), специального назначения (специализируемых клетках), реснички, жгутики.
Ядро(неделящееся) содержит: хроматин, ядрышко, кариоплазму (нуклеоплазма) и ядерную оболочку. Ядро является обязательным компонентом эукариотических клеток. Выполняет особые сложные функции:
— хранение основной генетической информации, её реализация (гл. роль в передаче наследственности);
— обеспечение синтеза белка;
— является регулятором всех жизненных отправлений клетки.
Без ядра клетки, как правило, жить не могут (погибают). Например, эритроциты без- ядерные – живут 48 суток.
Форма ядра бывает различная, обычно круглая, шарообразная. Размеры очень варьируют и зависят от функции клеток. Количество ядер различно. Обычно бывают одноядерные клетки, но и 2х и даже многоядерные (в печени, поперечно-полосатых мышечных волокнах).
Хроматин . Хромосомы.
Важнейшей составной частью ядра является хроматин (хрома — цвет)- вещество, хорошо окрашивающееся специальными ядерными красителями. Окрашивание обусловлено присутствием в хроматине ДНК и белков. Хроматин представлен глыбками и гранулами, располагается в кариоплазме (ядерном соке) – наблюдается в рабочем состоянии клетки (жизнь).
В неделящейся клетке (интерфаза) хроматин представляет собой разрыхлившиеся хромосомы (потерявшие свою форму) – это их рабочее состояние, с их участием происходят процессы транскрипции и редупликации ДНК (тонкие нити – диспирализованные). Хромосомы — место хранения наследственной информации.
Во время митотического деления (покой) хроматин представляет собой – хромосомы в виде спирализованных, плотных телец, которые выполняют функцию распределения и переноса генетического материала (ДНК) в дочерние клетки. Хроматин бывает 2х видов:
—эухроматин – окрашивается в покоящемся ядре не ярко (активный);
—гетерохроматин– хорошо окрашивается в состояние покоя и при делении (не активный). Он контролирует степень спирализации хромосом.
В хромосоме участки эу — и гетерохроматина чередуется. Половой хроматин – встречается в интерф. ядре соматических клеток особей женского пола. По периферии ядра клеток располагается глыбка хроматина – тельце Бара, у мужского пола эти отличия отсутствуют.
СТРОЕНИЕ ХРОМОСОМ.
1) Во всех клетках поддерживается постоянный набор хромосом. Обычно в соматических клетках (кл. тела) хромосомы парны (гомологичны)и составляют двойной, или диплоидный набор (2п) или полный. Диплоидный набор клеток человека – 46. Число у различных животных и растений широко отличаются, и не зависит от степени организации: кошка – 38, коровы – 60, реч. рак – 116, овца – 54, шимпанзе – 48.
2) Набор хромосом в клетках организмов, принадлежащих к одному виду, характеризуется определенными размерами, формами, числом и называются – кариотипом (у всех людей 46, у всех кошек 38). В половых клетках количество хромосом вдвое меньше (п) и называется гаплоидным: яйцеклетка =23, сперматозоид = 23.
3) Все хромосомы в клетке можно разделить на 2 группы: (по функции)
— аутосомы – не половые хромосомы,
— гетерохромосомы – половые хромосомы (они определяют половые особенности организма).
Кариотип человека представлен 46 хромосомами, из них 44- аутосомы и 2 половые хромосомы: 46=44ау.+2 пол.
| |
4) Каждая хромосома (в период деления) состоит из 2х половин – хроматид, спирально закрученных нитей (ДНК), связанных между собой центромерой (первичной перетяжкой). Хроматида – состоит из ДНК и вокруг белки. Хроматиды – состоят из хромонем (нитей). В состав хромосом входит 40% ДНК и 60% белка, так же входят РНК, липиды, углеводы. 5) Хромосомы в делящейся клетке имеют форму прямых или изогнутых палочек. Центромера делит хромосому на 2 плеча. В зависимости от места нахождения перетяжки и длины плеча хромосомы бывают нескольких типов: |
МЕТАЦЕНТРИЧЕСКИЕ (равноплечие) – центромера в центре хромосомы, оба плеча равны;
СУБМЕТАЦЕНТРИЧЕСКИЕ (неравноплечие) – центромера ближе к одному из концов хромосомы. Плечи имеют разную длину: короткое (р) и длинное (q);
АКРОЦЕНТРИЧЕСКИЕ (сильно неравноплечие) – центромера занимает место на конце хромосомы. Одно плечо длинное, другое совсем маленькое.
Встречаются хромосомы со вторичной перетяжкой (участок деления), перетяжка называется спутником (сателлитом).
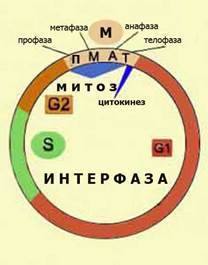
КЛЕТОЧНЫЙ ЦИКЛ И ЕГО ПЕРИОДЫ.
Период жизни клетки от момента её рождения в результате деления материнской клетки до следующего деления или смерти называется жизненным (клеточным) циклом клетки.
| |
Клеточный цикл способных к размножению клеток включает две стадии: — ИНТЕРФАЗУ (стадия между делениями, интеркинез); — ПЕРИОД ДЕЛЕНИЯ (митоз). В интерфазе происходит подготовка клетки к делению – синтез различных веществ, но главным является удвоение ДНК. По продолжительности она составляет большую часть жизненного цикла. Интерфаза состоит из 3–х периодов: 1) Предсинтетический – G1 (джи один) – наступает сразу после окончания деления. Клетка растет, накапливает различные вещества (богатые энергией), нуклеотиды, аминокислоты, ферменты. Готовится к синтезу ДНК. Хромосома содержит 1 молекулу ДНК (1 хроматида). 2) Синтетический – S происходит удвоение материала – репликация молекул ДНК. Усиленно синтезируется белки и РНК. Происходит удвоение числа центриолей. |
3) Постсинтетический G2 – предмитотический, продолжается синтез РНК. Хромосомы содержат 2 свои копии – хроматиды, каждая из которых несет по 1-ой молекуле ДНК (двунитевидная). Клетка готова к делению хромосома сперализуется.
Далее наступает процесс деления. Описано 3 способа деления эукар. клеток:
— амитоз – прямое деление
— митоз – непрямое деление
— мейоз – редукционное деление
АМИТОЗ – встречается редко, особенно у стареющих клеток или при патологических состояниях (репарация тканей), ядро остаётся в интефазном состоянии, хромосомы не сперализуются. Ядро делится путем перетяжки. Цитоплазма может и не делится, тогда образуются двуядерные клетки.
МИТОЗ – универсальный способ деления. В жизненном цикле он составляет лишь малую часть. Цикл эпитемальных клеток кишечника кошки составляет 20 – 22 ч., митоз – 1 час. Митоз состоит из 4-х фаз.
1)ПРОФАЗА – происходит укорочение и утолщение хромосом (спирализация) они хорошо видны. Хромосомы состоят из 2-х хроматид (удвоение в периоде интерфазы). Ядрышко и ядерная оболочка распадаются, цитоплазма и кариоплазма смешиваются. Разделившиеся клеточные центры расходятся по длинной оси клетки к полюсам. Образуется веретено деления (состоящее из упругих белковых нитей).
2)МЕТОФАЗА – хромосомы располагаются в одной плоскости по экватору, образуя метафазную пластинку. Веретено деления состоит из 2-х типов нитей: одни соединяют клеточные центры, вторые – (число их = числу хромосом 46) прикреплены, одним концом к центросоме (клеточному центру), другой к центромере хромосомы. Центромера тоже начинает делиться на 2. Хромосомы (в конце) расщепляются в области центромеры.
3)АНАФАЗА – самая короткая фаза митоза. Нити веретена деления начинают укорачиваться и хроматиды каждой хромосомы удаляются друг от друга по направлению к полюсам. Каждая хромосома состоит только из 1 хроматиды.
4)ТЕЛОФАЗА – хромосомы концентрируются у соответствующих клеточных центров, деспирализуются. Формируются ядрышки, ядерная оболочка, образуется мембрана, отделяющая сестринские клетки друг от друга. Сестринские клетки расходятся.
Биологическое значение митоза состоит в том, что в результате его каждая дочерняя клетка получает точно такой же набор хромосом, а следовательно, и точно такую же генетическую информацию, какими обладала материнская клетка.
Источник: megalektsii.ru
Длительность клеточного цикла
Длительность клеточного цикла у разных клеток варьируется. Быстро размножающиеся клетки взрослых организмов, такие как кроветворные или базальные клетки эпидермиса и тонкой кишки, могут входить в клеточный цикл каждые 12—36 ч. Короткие клеточные циклы (около 30 мин) наблюдаются при быстром дроблении яиц иглокожих, земноводных и других животных. В экспериментальных условиях короткий клеточный цикл (около 20 ч) имеют многие линии клеточных культур. У большинства активно делящихся клеток длительность периода между митозами составляет примерно 10—24 ч.
Фазы клеточного цикла
Клеточный цикл эукариот состоит из двух периодов:
-
Период клеточного роста, называемый «интерфаза», во время которого идет синтез ДНК и белков и осуществляется подготовка к делению клетки.
-
Периода клеточного деления, называемый «фаза М» (от слова mitosis — митоз).
Интерфаза состоит из нескольких периодов:
-
G1-фазы (от англ. gap — промежуток), или фазы начального роста, во время которой идет синтез мРНК, белков, других клеточных компонентов;
-
S-фазы (от англ. synthesis — синтез), во время которой идет репликация ДНК клеточного ядра, также происходит удвоение центриолей (если они, конечно, есть).
-
G2-фазы, во время которой идет подготовка к митозу.
У дифференцировавшихся клеток, которые более не делятся, в клеточном цикле может отсутствовать G1 фаза. Такие клетки находятся в фазе покоя G0.
Период клеточного деления (фаза М) включает две стадии:
-
кариокинез (деление клеточного ядра);
-
цитокинез (деление цитоплазмы).
В свою очередь, митоз делится на пять стадий.
Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатах световой и электронной микроскопии фиксированных и окрашенных клеток.
Регуляция клеточного цикла
Закономерная последовательность смены периодов клеточного цикла осуществляется при взаимодействии таких белков, как циклин-зависимые киназы и циклины. Клетки, находящиеся в G0 фазе, могут вступать в клеточный цикл при действии на нихфакторов роста. Разные факторы роста, такие как тромбоцитарный, эпидермальный, фактор роста нервов, связываясь со своими рецепторами, запускают внутриклеточный сигнальный каскад, приводящий в итоге к транскрипции генов циклинов ициклин-зависимых киназ. Циклин-зависимые киназы становятся активными лишь при взаимодействии с соответствующими циклинами. Содержание различных циклинов в клетке меняется на протяжении всего клеточного цикла. Циклин является регуляторной компонентой комплекса циклин-циклин-зависимая киназа. Киназа же является каталитическим компонентом этого комплекса. Киназы не активны без циклинов. На разных стадиях клеточного цикла синтезируются разные циклины. Так, содержание циклина B в ооцитах лягушки достигает максимума к моменту митоза, когда запускается весь каскад реакций фосфорилирования, катализируемых комплексом циклин-В/циклин-зависимая киназа. К окончанию митоза циклин быстро разрушается протеиназами.
Источник: StudFiles.net
Интерфаза: период G1 (пресинтетический)
Пресинтетический период G1 наступает сразу после деления клетки. В этом периоде клетка имеет диплоидное содержание ДНК на одно ядро, т.е. в клетке содержится двойной набор хромосом. Генетический материал пребывает в виде слабо спирализованных цепей ДНК или хроматина. В пресинтетическом периоде генетический материал клетки не копирован и клетка только начинает подготовку к репликации ДНК.
В периоде G1 клетка начинает расти и увеличиваться в размерах за счет накопления клеточных белков. Также клетка синтезирует мРНК. Таким образом клетка готовится к синтезу ДНК и последующему митозу.
На развитие клетки в периоде G1 и других фазах могут влиять факторы роста: температура, питательные вещества, пространство для роста. Нормальный синтез мРНК и белков происходит за счет достаточного количества аминокислот. А продуктивный рост клеток обеспечивает нормальная физиологическая температура тела.
Клетка готова к переходу в следующую фазу после достижения ею определенных размеров и синтезирования всех необходимых белков.
Интерфаза: период S (синтез ДНК)
В периоде S происходит репликация (синтез или копирование) ДНК. Точность синтеза ДНК важна для предотвращения появления генетических аномалий, которые ведут к нарушению работы клетки или ее гибели. Поэтому регуляторные процессы периода S клеточного цикла очень консервативны.
В течение периода S интенсивно синтезируются РНК и белки, которые связанны с ДНК. Они необходимы для строения новой хроматиды. Осуществляют синтез этих белков, свободные рибосомы, которые не связанны с эндоплазматической сетью.
Целью репликации ДНК является создание двух идентичных хроматид. Дочерние копии ДНК синтезируются на каждой материнской нити. Репликация ДНК начинается в конкретных участках, которые называются ori. У млекопитающих насчитывается около 100 ori на каждой ДНК. Синтез распространяется от точек ori в две стороны по нитям ДНК.
В клетке в периоде G1 закладываются специальные пререпликационные комплексы, которые препятствуют синтезу более одной хромосомы в ДНК. Эти комплексы разбираются перед началом репликации в периоде S. На нитях ДНК разрушаются специальные белки, которые предотвращают прикрепление репликационного аппарата к ДНК после начала синтеза.
Кроме репликации ДНК, в периоде S также происходит удвоение центриолей клеточного центра. Материнская центриоль строит в клетке новую центриоль, дочернюю. Материнская центриоль задействована в сборке микротрубочек.
Интерфаза: период G2 (постсинтетический)
Окончание стадии S является началом стадии G2. Постсинтетический период – это финальная стадия интерфазы клеточного цикла. В этом периоде происходит интенсивное деление митохондрий и процессы биосинтеза, а также концентрация энергетических запасов. В периоде G2 клетка заканчивает приготовления к митозу.
В периоде G2 синтезируется иРНК, необходимая для прохождения митоза. Также синтезируется РНК рибосом, которые определяют деление клетки. Кроме того, в это же время синтезируются белки митотического веретена — тубулина. Синтез РНК резко снижается в конце периода G2 и полностью прекращается во время митоза.
Перед митозом в конце периода G2 происходит окончательная проверка размеров клетки, полнота репликации и целостности ДНК.
Митоз
Деление клетки наступает после успешного прохождения ею всех этапов интерфазы. Деление клетки подразумевает образование двух идентичных дочерних клеток из родительской. Во время митоза происходит делений ядра клетки с сохранением числа хромосом. В каждом ядре дочерних клеток находится одинаковый с родительским набор хромосом. После деления ядра происходит деление самой клетки. В митозе выделяют четыре стадии, основываясь на внешний вид хромосом: профаза, метафаза, анафаза, телофаза.
Профаза. Для профазы характерно утолщение и укорочение хромосом как результат их спирализации. В этот период двойные хромосомы состоят их связанных между собой сестринских хроматид. Параллельно со спирализацией пропадает ядрышко и распадается на отдельные фрагменты ядерная оболочка. После ликвидации ядерной оболочки хромосомы свободно лежать в цитоплазме. Во время профазы центриоли клетки расходятся к ее полюсам. Веретено деления начинает формироваться в конце профазы. Оно образуется из микротрубочек с помощью полимеризации белковых субъединиц
Метафаза. Завершение формирования веретена деления происходит в метафазе. Веретено деления состоит их хромосомных и центросомных микротрубочек. Хромосомные микротрубочки связываются с центромерами хромосом, а центросомные – тянутся от полюса к полюсу клетки. К микротрубочкам веретена деления прикреплена каждая двойная хромосома, и они выталкиваются микротрубочками, стремясь расположиться на равных расстояниях от полюсов клетки. Находясь в одной плоскости, они образуют метафазную пластинку. В этот период очень хорошо видно двойное строение хромосом.
Анафаза. В периоде анафазы дочерние хромосомы растягиваются к полюсам клетки с помощью микротрубочек веретена деления. Дочерние хромосомы несколько изгибаются, что позволяет им расходиться к полюсам. Анафаза характеризуется наличием в клетке двух диплоидных наборов хромосом.
Телофаза. В телофазе происходит раскручивание хромосом. Они раздуваются, и их строение становятся плохо видно под микроскопом. У каждого полюса вокруг хромосом формируется ядерная оболочка из мембранных структур цитоплазмы. В ядрах появляются ядрышки. Веретено деления начинает разрушаться. В телофазе осуществляется разделение цитоплазмы и формирование двух клеток.
Регуляторы клеточного цикла
Фазы клеточного цикла регулируются с помощью вне- и внутриклеточных механизмов. Внеклеточные механизмы обусловлены влиянием факторов роста, цитокинов, нейрогенных и гормональных стимуляторов. Каждая фаза клеточного цикла и их последовательная смена осуществляются под действием особых белков цитоплазмы: циклинов и циклин-зависимых киназ.
Циклины – это специфические ферменты, которые активируют циклин-зависимые киназы. Внутриклеточная концентрация циклинов меняется в зависимости от стадии прохождения клеточного цикла. Циклины активируют циклин-зависимые киназы в результате взаимодействия их молекул. Образование молекулярного комплекса происходит при максимальной концентрации циклина. Существует несколько видов циклинов, которые имеют ключевое значение в смене фаз клеточного цикла. Увеличение концентрации каждого из них является сигналом для прохождения в очередной период клеточного цикла. Максимальная концентрация циклина Е происходит при переходе в период S; концентрация циклина А – при переходе в период G2; концентрация циклина В – при переходе в период М. В процессе клеточного цикла каждый циклин в свое время разрушается протеасомами.
Циклин-зависимые киназы – это разновидность белков, которые регулируются циклином и циклиноподобными молекулами. Циклин-зависимые киназы регулируют транскрипцию и процессинг мРНК, а также участвуют в процессе смены фазы клеточного цикла. Каждая циклин-зависимая киназа активируется соответствующим циклином. При этом циклин-зависимые киназы, в основном, имеют схожее строение и отличаются только способом сочетания циклинов. В отличие от циклинов, уровень циклин-зависимых киназ остается более-менее стабильным на протяжении всех периодов клеточного цикла.
Совместная работа циклинов и циклин-зависимых киназ выглядит следующим образом:
1.Циклин Д синтезируется в ответ на пролиферативный стимул.
2.Циклин Д связывается со специальной циклин-зависимой киназой, и запускает работу транскрипционных факторов, которые участвуют в синтезе циклина Е, циклина А и синтезе ДНК.
3.Циклин Е связывается с соответствующей циклин-зависимой киназой и запускает период S.
4. Циклин В с активированной циклин-зависимой киназой инициируют переход в период митоза.
5.Период митоза заканчивается после разрушения комплекса циклина В с циклин-зависимой киназой.
Источник: www.oncoforum.ru
КЛЕ́ТОЧНЫЙ ЦИКЛ, набор строго скоординированных во времени процессов, приводящих к воспроизведению клеток; важнейший механизм, с помощью которого репродуцируется всё живое. В основе способности клеток к самовоспроизведению лежит уникальное свойство ДНК реплицироваться с образованием двух точных копий, которые в ходе деления распределяются между двумя дочерними клетками. Помимо удвоения ДНК в К. ц. большинства клеток должно удваиваться число их органелл и макромолекул, в противном случае с каждым делением клетки становились бы меньше. Т. о., для поддержания своих размеров клетка должна координировать рост клеточной массы с её делением. У одноклеточных организмов К. ц. совпадает с жизненным циклом. В непрерывно размножающихся клетках многоклеточных организмов он состоит из чётко сменяющих друг друга периодов: интерфазы (подготовки к делению) и митоза (собственно деления).
Именно в интерфазе, а не в митозе, как считали ранее, происходит репликация ДНК (удвоение хромосом). Сама интерфаза подразделяется на три фазы. Пресинтетическая G1-фаза предшествует синтезу ядерной ДНК и следует сразу за предыдущим делением. В этой фазе в клетке синтезируются рибонуклеиновые кислоты (РНК), разл. белки, увеличивается число рибосом, митохондрий. В целом наблюдается активный рост клетки. В фазе синтеза ДНК (S-фазе) продолжается образование РНК и белков, происходит репликация ядерной ДНК. Постсинтетическая G2-фаза характеризуется окончательной подготовкой клетки к делению, в т. ч. сборкой микротрубочек, которые в дальнейшем будут формировать веретено деления. За фазами интерфазы следует митоз (М-фаза). Понятие «К. ц.» включает в себя не только интервал времени от одного клеточного деления до другого. Иногда клетка, завершившая митоз, прекращает подготовку к очередным синтезу ДНК и митозу и входит в состояние «вне цикла», т. е. в состояние пролиферативного покоя, обозначаемое как G0-фаза. Из этой фазы она может вновь вернуться в состояние К. ц. под влиянием стимула к размножению. Существуют также клетки с удлинённой продолжительностью постсинтетической фазы. Их называют G2-популяцией.
Длительность К. ц. варьирует у разных организмов в широких пределах – от нескольких минут до нескольких суток. Показано также, что разные клеточные популяции могут сильно различаться и по продолжительности отд. фаз цикла. Наиболее коротким является К. ц. бластомеров на ранних стадиях эмбрионального развития. В К. ц. этих клеток практически отсутствуют G1- и G2-фазы, S-фаза может длиться всего неск. минут. На заключит. этапе эмбрионального развития животных, напр. у млекопитающих, происходит массовое вступление клеток в состояние покоя, связанное с их дифференцировкой. Длительность К. ц. различается в пределах одного взрослого многоклеточного организма. Напр., клетки регенерирующей печени мыши делятся примерно через каждые 48 ч, а крипт кишки (углублений эпителия в её слизистой) – через 10,5 ч. Лимфоциты человека, помещённые в среду для культивирования клеток и обработанные фитогемагглютинином, начинают размножаться, и их К. ц. оказывается равным примерно 24 часам. Наиболее вариабельной является G1-фаза. При дифференцировке одни типы клеток взрослого организма, выполняя специфич. функции, находятся в G0-фазе, но сохраняют способность к размножению, другие – необратимо утрачивают её и называются терминально дифференцированными.
Программа осуществления процессов К. ц. включает сложную систему контроля, которая обеспечивается работой комплексов регуляторных белков. При этом контролирующие системы действуют как внутри клетки (эндогенные регуляторы), так и вне её (экзогенные регуляторы). Эндогенная регуляция клеточного цикла основана на циклич. активации протеинкиназ, известных как циклин-зависимые киназы. Активность последних увеличивается или падает в ходе К. ц. Такие колебания активности ведут к циклич. изменениям степени фосфорилирования внутриклеточных белков, инициирующих или регулирующих важнейшие события К. ц.: репликацию ДНК, сегрегацию хромосом и цитокинез (собственно деление тела клетки на две). У разных организмов в разных фазах К. ц. активируются разл. варианты комплексов циклинов и киназ. Прекращение работы таких комплексов связано с действием ингибирующих белков или с протеолизом циклинового компонента комплекса. В целом эндогенная регуляция в ходе К. ц. обеспечивается сложной системой процессов синтеза и деградации определённых субъединиц регуляторных комплексов, процессами фосфорилирования и дефосфорилирования этих и др. регуляторых белков. Экзогенная регуляция включает действие на клетки внеклеточных регуляторных сигналов. Для одноклеточных организмов такими сигналами являются компоненты окружающей среды, для многоклеточных организмов – синтезируемые разл. клетками небольшие белковые молекулы, к которым относятся факторы роста и цитокины. Нарушение механизмов эндогенной и экзогенной регуляции К. ц. может приводить к неконтролируемому размножению клеток, что служит причиной образования опухолей.
Источник: bigenc.ru