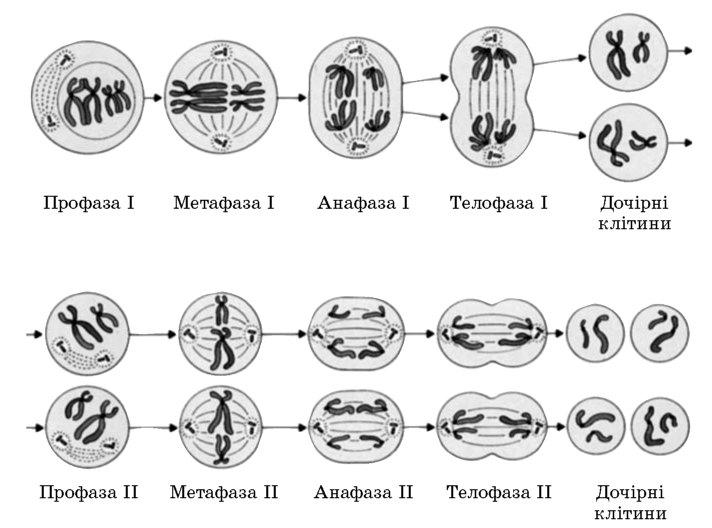
Мейоз та його фази
Мейоз (грец. Meiosis — зменшення, спадання) або редукційний поділ.
В результаті мейозу відбувається зменшення числа хромосом, тобто з диплоїдного набору хромосом (2п) утворюється гаплоїдний (п).
Мейоз складається з 2-х послідовних поділів:
I розподіл називається редукционное або зменшувальне.
II розподіл називається екваціонное або зрівняльний, тобто йде за типом мітозу (значить число хромосом в материнській і дочірніх клітинах залишається колишнім).
Біологічний сенс мейозу полягає в тому, що з однієї материнської клітини з диплоїдним набором хромосом утворюється чотири гаплоїдні клітини, таким чином кількість хромосом зменшується в два рази, а кількість ДНК в чотири рази. В результаті такого поділу утворюються статеві клітини (гамети) у тварин і суперечки у рослин.
Фази називаються також як і в мітозі, а перед початком мейозу клітина також проходить інтерфазу.
Профаза I — найтриваліша фаза і її умовно ділять на 5 стадій:
1) лептонеми (Лептотена) — або стадія тонких ниток.
е спирализация хромосом, хромосома складається з 2-х хроматид, на ще тонких нитках хроматид видно потовщення або згустки хроматину, які називаються — хромомер.
2) зігонеми (Зиготена, грец. Зливаються нитки) — стадія парних ниток. На цій стадії попарно зближуються гомологічні хромосоми (однакові за формою величиною), вони притягуються і прикладаються один до одного по всій довжині, тобто коньюгіруют в області хромомер. Це схоже на замок «блискавка». Пару гомологічниххромосом називають біваленти. Число бівалентов одно гаплоидному набору хромосом.
3) пахінема (Пахітена, грец. Товста) — стадія товстих ниток. Йде подальша спирализация хромосом. Потім кожна гомологичная хромосома розщеплюється в поздовжньому напрямку і стає добре видно, що кожна хромосома складається з двох хроматид такі структури називають тетрадами, тобто 4 хроматиди. В цей час йде кроссинговер, тобто обмін гомологічними ділянками хроматид.
4) Діплонема (діплотена) — стадія подвійних ниток. Гомологічні хромосоми починають відштовхуватися, відходять один від одного, але зберігають взаємозв’язок за допомогою містків — хиазм, це місця де відбудеться кроссинговер. У кожному з’єднанні хроматид (тобто хіазмі), здійснюється обмін ділянками хроматид. Хромосоми спирализуются і коротшають.
5) діакінеза — стадія відокремлених подвійних ниток. На цій стадії хромосоми повністю ущільнені і інтенсивно фарбуються. Ядерна оболонка і ядерця руйнуються. Центриоли переміщаються до полюсів клітини і утворюють нитки веретена поділу.
Хромосомний набір профази I становить — 2п4с.
Таким чином, в профазу I відбувається:
1. кон’югація гомологічних хромосом;
2. освіту бівалентов або тетрад;
3. кроссинговер.
Залежно від кон’югированія хроматид можуть бути різні види кросинговеру: 1 — правильний або неправильний; 2 — рівний або нерівний; 3 — цитологічний або ефективний; 4 — одиничний або множинний.
Метафаза I — спирализация хромосом досягає максимуму. Біваленти вишиковуються уздовж екватора клітини, утворюючи метафазну пластинку. До Центромера гомологічниххромосом кріпляться нитки веретена поділу. Біваленти виявляються з’єднаними з різними полюсами клітини.
Хромосомний набір метафази I становить — 2п4с.
Анафаза I — центромери хромосом не діляться, фаза починається з розподілу хіазм. До полюсів клітини розходяться цілі хромосоми, а не хроматиди. У дочірні клітини потрапляє тільки по одній з пари гомологічних хромосом, тобто йде їх випадкове перерозподіл. На кожному полюсі, виявляється, по набору хромосом — 1п2с, а в цілому хромосомний набір анафази I становить — 2п4с.
Телофаза I — по полюсах клітини знаходиться цілі хромосоми, що складаються з 2-х хроматид, але кількість їх стало в 2 рази менше.
У тварин і деяких рослин хроматиди деспирализуются. Навколо них на кожному полюсі формується ядерна мембрана.
Потім йде цитокинез.
Хромосомний набір утворилися після першого поділу клітин становить — п2с.
Між I і II поділами немає S-періоду і не йде реплікація ДНК, тому що хромосоми вже подвоєні і складаються з сестринських хроматид, тому інтерфазу II називають інтеркінез — тобто відбувається переміщення між двома поділами.
Профаза II — дуже коротка і йде без особливих змін, якщо в телофазу I не утворюється ядерна оболонка, то відразу утворюються нитки веретена поділу.
Метафаза II — хромосоми вишиковуються уздовж екватора. Нитки веретена поділу кріпляться до Центромера хромосом.
Хромосомний набір метафази II становить — п2с.
Анафаза II — центромери діляться і нитки веретена поділу розводять хроматиди до різних полюсів. Сестринські хроматиди називаються дочірніми хромосомами (або материнські хроматиди це і будуть дочірні хромосоми).
Хромосомний набір анафази II становить — 2п2с.
Телофаза II — хромосоми деспирализуются, розтягуються і після цього погано помітні. Утворюються ядерні оболонки, ядерця. Телофаза II завершується цітокінезом.
Хромосомний набір після телофази II становить — пс.
Значення мейозу.
1. Підтримується постійний число хромосом у видів, що розмножуються статевим способом, тому що при злитті гаплоїдних клітин відновлюється диплоїдний набір хромосом.
2. Утворюється велика кількість різних комбінацій батьківських і материнських хромосом, за рахунок незалежного розходження гомологічних хромосом в анафазу I. Число комбінацій пар хромосом визначається як 2n, де n — гаплоїдний набір хромосом. У людини число комбінацій одно 223 = 8388608.
3. Відбувається перекомбінація генетичного матеріалу, за рахунок кросинговеру, який йде в профазу I, на стадії пахінема.
Схема мейотичного поділу.
Источник: blog.r13-r21.com.ua
Первое мейотическое деление
В результате первого мейотического (редукционного) деления из диплоидных клеток (2n) образуются гаплоидные (n). Он начинается из профазы І, в которой, также как и в митозе, происходит упаковка наследственного материала (спирализация хромосом). Одновременно гомологические (парные) хромосомы сближаются одинаковыми участками – происходит коньюгация. В результате коньюгации образуются пары хромосом – биваленты. Каждая хромосома, вошедшая в мейоз, состоит из двух хроматид иимеет удвоенный наследственный материал, потому бивалент состоит из 4 нитей. Когда хромосомы находятся в коньюгированном состоянии, их дальнейшая спирализация продолжается. Отдельные хроматиды гомологических хромосом переплетаются и перекрещиваются. В дальнейшем гомологические хромосомы отталкиваются и немного расходятся, потому в местах переплетения хроматид может произойти их разрыв. Как результат в процессе возобновления разрывов у хроматиды гомологических хромосом происходит обмен соответствующими участками. В итоге перешедшая от родителя к данному организму хромосома, содержит часть материнской хромосомы, и наоборот.
После кроссинговера уже изменённые хромосомы, то есть с другими объединениями генов, расходятся.
Поскольку кроссинговер является процессом закономерным, он каждый раз приводит к обмену разными по размеру участками и, таким образом, обеспечивается эффективная рекомбинация материала хромосом гамет.
- В метафазе І завершает формироваться веретено деления. Его нити крепятся к центромерам хромосом, которые соединены в биваленты так, что от каждой центромеры отходит лишь одна нить к одному из полюсов клетки. В результате с помощью связанных с центромерами гомологических хромосом нитей биваленты располагаются по экватору веретена деления.
- В анафазе І гомологические хромосомы рассоединяются и расходятся к полюсам клетки.
В телофазе І возле полюсов веретена собирается одинарный (гаплоидный) набор хромосом, в котором каждый их вид представлен уже не парой, а одной хромосомой, состоящей из двух хроматид. В короткой по длительности телофазе возобновляется оболочка ядра, а материнская клетка делится на две дочерние. Таким образом, благодаря образованию бивалентов при коньюгации гомолологических хромосом в профазе І мейоза создаёт условия для дальнейшей редукции количества хромосом. Формируется гаплоидный набор в гаметах, который обеспечивается расхождением в анафазе І не хроматид, как в митозе, а гомологических хромосом, которые ранее были соединены в биваленты.
Второе мейотическое деление
Второе мейотическое деление происходит сразу же после первого и подобно обычному митозу (потому его ещё называют митозом мейоза), но клетки, которые делятся, содержат гаплоидный набор хромосом.
- Профаза ІІ недлительна.
- В метафазе ІІ снова образуется веретено деления, хромосомы располагаются в экваториальной плоскости, а центромеры соединяются с микротрубочками веретена деления.
- В анафазе ІІ их центромеры рассоединяются и каждая хроматида превращается в самостоятельную хромосому. Дочерние хромосомы, которые отделяются друг от друга, направляются к полюсам клетки.
- В телофазе ІІ завершается расхождение хромосом и клетки делятся: из двух гаплоидных клеток образуются четыре гаплоидные дочерние клетки.
Значение мейоза
Благодаря редукционному делению регулируется непрерывное увеличение числа хромосом в процессе слиянии гамет. Если бы не было этого механизма, то во время полового размножения число хромосом удваивалось бы из поколения в поколение.
Ещё одно важное значение мейоза: обеспечение большого разнообразия генетического состава гамет как в результате кроссинговера, так и в результате различного объединения отцовских и материнских хромосом при их расхождении в анафазе І мейоза. Это обеспечивает разнообразие и разнокачественность потомства во время полового размножения.
Источник: spravochnick.ru
6. Цитологічна та генетична характеристика стадій мейозу. Випадкова комбінація гомологів (батьківських і материнських) з різних пар в гаметах при мейозі.
Мейоз складається з двох слідуючих один за одним поділів при одній редуплікації ДНК в інтерфазі перед першим поділом гаметоцита першого порядку. Кожен з двох поділів складається з 4-х фаз: профаза, метафаза, анафаза, телофаза. Найбільш складний з них перший поділ, а найбільш складна з фаз — перша профаза.
Перший мейотичний поділ називають ще редукційним (редукція — зменшення), тому що вже після нього дочірні клітини (гаметоцити ІІ порядку) стають гаплоїдними.
I профаза складніша, тривала; у людини вона триває близько 2-х діб. Виділяють 5 морфологічно помітних стадій: 1) лептонема (лепто — тонкий) — стадія тонких ниток; 2) зигонема (зигос — разом) — стадія ниток, що злилися; 3) пахінема (пахи — товстий) — стадія товстих ниток; 4) диплонема (дипло — подвійний) — стадія подвійних ниток; 5) діакінез — стадія відособлення шляхом відходження один від одного двох ниток.
Головні події в профазі І:
-
кон’югація гомологічних хромосом (лептонема, зигонема);
-
кросинговер між сестринськими хроматидами гомологів (пахінема);
-
вибірковий синтез іРНК, рРНК (диплонема);
-
остаточне формування бівалентів (діакінез).
І метафаза.
Спарені гомологи, тобто біваленти вишиковуються в області екватора (перша відмінність від мітозу). Кінетохорні нитки обох хроматид одного бівалента обернені тільки до одного полюса, до якого він орієнтований (друга відмінність від мітозу). При цьому вони вступають в зв’язок з полюсними мікротрубочками.
І анафаза.
Відбувається розпад хіазм, що зв’язують батьківські і материнські хромосоми, відходження цілих гомологів, що складаються з двох хроматид. Тому з генетичної точки зору в анафазі відбувається розходження алельних генів, розташованих в різних гомологах.
I телофаза.
Коли хромосоми досягають полюсів поділу, вони деконденсуються, навколо них утворюється ядерна оболонка, наступає цитокінез і клітини переходять в коротку інтерфазу. В результаті першого мейотичного поділу утворюється дві клітини з гаплоїдним набором хромосом (1n), оскільки в них один гомолог з пари, але диплоїдний набір ДНК, оскільки гомологи складаються з двох хроматид, призначених для двох клітин. Тому після короткої інтерфази слідує другий мейотичний поділ.
Другий мейотичний поділ. Його ще називають екваційним (эквел — рівний), оскільки його суть полягає в розходженні хроматид, внаслідок чого утворюються генетично рівні клітини (якщо не враховувати результати кросинговера). Отже цей поділ за своїми генетичними і цитологічними закономірностями нічим не відрізняється від мітозу, хіба що відбувається в гаплоїдній клітині. В результаті другого поділу утворюються 4 клітини з 1n хромосом і 1n ДНК.
Біологічне значення мейозу і статевого розмноження:
1) в результаті мейозу утворюються гамети з гаплоїдним числом хромосом, що забезпечує диплоїдний набір хромосом і його постійність у ряді поколінь при статевому розмноженні, яке супроводжується злиттям гаплоїдних гамет з утворенням диплоїдної зиготи;
2) мейоз забезпечує перекомбінацію алельних генів в гаметах, внаслідок чого кожна гамета має унікальний набір генів. Перекомбінація алельних генів відбувається завдяки трьом цитологічним процесам мейозу.
2.1. Кросинговер. Кросинговер відбувається в пахінемі профази I між хроматидами гомологічних хромосом. Кросинговер забезпечує перекомбінацію алельних генів в групах зчеплення.
2.2. Випадкова комбінація гомологів (батьківських і материнських) з різних пар в гаметах.
Батьківські і материнські гомологи різних бівалентів випадково орієнтуються до полюсів поділу в метафазі I, оскільки рухи бівалентів щодо один одного незалежні. Тому розбіжність жіночих і чоловічих гомологів з різних пар в анафазі I і потрапляння їх в різні клітини випадкове. Внаслідок чого в гаметах відбувається незалежна комбінація гомологів з різних пар, тобто перекомбінація різних груп зчеплення генів. Випадкова (незалежна) комбінація жіночих і чоловічих гомологів різних пар в гаметах є цитологічною основою 3-го закону Менделя — закону незалежної комбінації ознак.
Число гамет з різною комбінацією батьківських і материнських гомологів (Уг) з різних пар можна обчислити за формулою степеневої функції Уг = 2х, де 2 — число комбінацій, яке утворює пара гомологічних хромосом (у одну клітину відходить жіночий гомолог, а в іншу — чоловічий), х — число пар гомологічних хромосом, що комбінуються, в клітині (бівалентів в метафазі I).
2.3. Запліднення алгебраїчно можна звести до множення комбінацій в гаметах. Тому число комбінацій жіночих і чоловічих гомологів з різних пар в зиготах виражається формулою: Уз = 2х ♀ х 2х ♂ = 22х, де 2х ♀ та 2х ♂ — число комбінацій у відповідних гаметах. Приклади для видів: 2n = 4; 2n = 8; 2n = 46. Якщо до комбінацій різних груп зчеплення генів, додати число гомологів з різних пар, тобто до комбінацій комбінацій, які утворюються в результаті кросинговера, тобто перекомбінації генів усередині групи зчеплення, то число комбінацій гамет і зигот безкінечне. Отже, практично можна вважати, що кожна гамета і кожна зигота унікальні по набору алельних генів. Звідси розширення біологічного значення статевого розмноження полягає в наступному.
1. З виникненням мейозу, який призводить до утворення гаплоїдних гамет, стала можливою диплофаза організму, тобто диплоїдний набір хромосом (2n). Диплофаза у вищих організмів (вищий рослин, багатоклітинних тварин) стала основною в їх життєвому циклі. Перевага диплофази над гаплофазою полягає в тому, що, по-перше, збільшується доза алельних генів, що супроводжується прискоренням обмінних процесів; по-друге, приводить до появи нових морфологічних варіантів; по-третє, в диплофазі приховуються, тобто не виявляються шкідливі рецесивні гени, а, отже, підвищується стійкість організмів в зовнішньому середовищі.
2. Статеве розмноження в цілому забезпечує перекомбінацію генетичної інформації між особинами популяції, тобто комбінативну мінливість. Це призводить до нових морфологічних варіантів особин. Генетична різноманітність особин сприяє ефективності рушійного природного відбору, який діє в умовах середовища, що змінюються. У цих умовах відбираються особини з новим поєднанням ознак, адекватних новим умовам, які склалися, що призводить надалі до появи нового виду. Отже, статеве розмноження, з його комбінативною мінливістю сприяє прогресивній еволюції. Треба підкреслити, що комбінативна мінливість прискорює еволюційний процес. Вона є основною для диплоїдних організмів при прогресивній еволюції в порівнянні з мутаційною мінливістю. Одних мутацій недостатньо для еволюції, тобто вони спостерігаються в середньому 10-5 — 10-7 ген на покоління. За допомогою статевого розмноження швидко утворюються потрібні комбінації ознак, відповідні новим умовам середовища.
Контрольні питання:
-
Чому при вегетативному розмноженні утворюються організми з ідентичним генотипом, тоді як при статевому завжди з різним?
-
Що загального при власне статевому розмноженні (копуляції) і його аналогах?
-
Перерахувати головні цитологічні перетворення мітозу при його еволюції в мейоз.
-
Що таке синаптонемальний комплекс і його значення.
-
Як цитологічно виглядає кросинговер? На якій стадії профази він відбувається і коли цитологічно проявляється?
-
При якій формі природного добору (стабілізуючий, рушійний) превалює вегетативне розмноження, а при якому статеве?
-
Назвіть три механізми комбінативної мінливості, обумовленої генетичними закономірностями мейозу.
Література: основна — 1-5, 8; додаткова — 1-9.
Источник: StudFiles.net
Мітоз
Мітоз — це непрямий поділ клітин. Є основним способом поділу соматичних клітин. Процес мітозу має чотири стадії — профаза, метафаза, анафаза й телофаза.
Схема мітоза

Стадії мітозу
|
Стадія |
Характеристика |
|
Профаза |
До початку профази клітина зазвичай округлюється, в’язкість її цитоплазми зростає. Морфологічно хромосоми помітні внаслідок спіралізації. У міру перебігу профази хромосоми коротшають, товщають, спадковий матеріал щільно впаковується за довжиною. Оболонка клітини товщає. Ядерна оболонка розчиняється, хромосоми переміщуються в центр клітини, утворюючи екваторіальну пластинку. Центріолі клітинного центру розходяться до полюсів, між ними формується веретено поділу. Генетична характеристика — 2n4c |
|
Метафаза |
Морфологічно хромосоми впорядковано лежать в зоні екватора. У цей момент вони добре помітні. Надщеплення хромосом на хроматиди доходить до центромери (центральна перетяжка хромосом). Каріотип організму вивчають саме на препараті мета-фазної клітини. Генетична характеристика — 2n4c |
|
Анафаза |
Це стадія повного роз’єднання сестринських хромосом і розходження їх до полюсів. Результатом є утворення двох груп хромосом. У перебігу анафази важливу роль відіграють основні компоненти цитоскелета: мікротрубочки та мікрофіламенти. Генетична характеристика — 4n4c (на полюсах по 2n2c) |
|
Телофаза |
На другій стадії відбуваються процеси цитокінезу. Хромосоми деспіралізуються, навколо них концентрується ядерна речовина й формується ядерна оболонка. Генетична характеристика — 4n4c. Цитотомія здійснюється утворенням внутрішньоклітинної пластинки в рослинних клітинах або утворенням борозни поділу у тваринних клітинах. У результаті останньої фази мітозу утворюються дві клітини, кожна з яких має повний диплоїдний набір: 2n2c |
|
Біологічне значення мітозу полягає в точному розподілі генетичної інформації між дочірніми клі- |
тинами й у підтриманні сталості кількості хромосом. |
Метафазна хромосома
Мейоз
|
Мейоз — форма ядерного поділу, що супроводжується зменшенням кількості хромосом від диплоїдної (2n) до гаплоїдної (n). При цьому в материнській клітині відбувається одноразове подвоєння хромосом (реплікація ДНК, як під час |
мітозу), за яким ідуть два цикли клітинних і ядерних поділів (перший і другий поділ мейозу). Таким чином, одна диплоїдна клітина дає початок чотирьом гаплоїдним клітинам. |
Мейоз
Стадії мейозу
|
Стадія |
Характеристика |
|
Профаза I |
Це найбільш тривала і складна стадія мейозу. У ній розрізняють п’ять послідовних періодів. 1. Лептотена — стадія довгих, тонких, слабко спіралізованих хромосом. У цей час закінчується реплікація ДНК. 2. Зиготена — стадія попарного з’єднання гомологічних хромосом. Це явище називається кон’югацією. 3. Пахітена — стадія товстих ниток. Гомологічні хромосоми поєднані в пари — біваленти. У цей час хромосоми в процесі кон’югації переплітаються й відбувається обмін гомологічними ділянками хроматид — кросинговер (crossing-over — перехрест). 4. Диплотена — період, коли гомологічні хромосоми починають відштовхуватись одна від одної, але в ділянках кросинговеру залишаються з’єднаними. 5. Діакінез — період, у який відштовхування гомологічних хромосом триває. Безпосередньо після діакінезу ядерна оболонка руйнується |
|
Метафаза I |
Об’єднані попарно хромосоми розміщуються в центрі клітини, утворюючи екваторіальну пластинку. Хромосоми з’єднуються з полюсами випадковим чином, тому в анафазі щоразу виникає нова комбінація хромосом, а надалі утворюються й різні гамети. Генетична характеристика, як і раніше,— 2n4c |
|
Анафаза I |
У результаті цієї фази мейозу відбувається поділ хромосом на два гаплоїдні набори, що потрапляють у дочірні клітини. Причому, на відміну від мейозу, розходяться не хроматиди, а цілі хромосоми. Генетична характеристика клітини — 2n4c (у цілому), на полюсах — по n2c |
|
Телофаза I |
Хромосоми розійшлися до протилежних полюсів. Їхня кількість стає вдвічі меншою, кожна складається з двох хроматид. Навколо хромосом концентрується ядерна речовина й утворюється ядерна оболонка. Далі відбувається поділ цитоплазми, аналогічний тому, що спостерігається під час мітозу. Генетична характеристика клітин, що утворилися,— n2c |
|
Інтерфаза II |
Головна особливість інтерфази другого поділу мейозу полягає у відсутності реплікації ДНК. Це спричиняє появу гаплоїдного набору хромосом у клітинах, що утворюються. Генетична характеристика — n2c |
|
Профаза II |
У цій фазі руйнуються ядерця та ядерні мембрани, хромосоми коротшають і товщають. Центріолі переміщуються до протилежних полюсів клітини. Генетична характеристика — n2c |
|
Метафаза II |
Хромосоми розташовуються вздовж екватора клітини. Генетична характеристика — n2c |
|
Анафаза II |
Відбувається поділ центромер, хроматиди відокремлюються одна від одної й тепер називаються хромосомами, вони містять у цілому гаплоїдний набір генів. Генетична характеристика клітини — 2n2c (у цілому), на полюсах — nc |
|
Телофаза II |
Ідентична телофазі мітозу. Генетична характеристика клітин, що утворюються,— nc |
|
В організмів, що розмножуються статевим шляхом, у результаті мейозу утворюються гаплоїдні клітини, хромосомні набори яких зливаються, формуючи ди-плоїдний набір зиготи. Якби не було мейозу, злиття гамет вело б до подвоєння кількості хромосом у кожному новому поколінні, народженому статевим шляхом. Отже, мейоз забезпечує збереження кількісної сталості генетичного матеріалу. |
Мейоз створює також можливості для виникнення в гаметах нових комбінацій генів, що є причиною появи нових ознак у потомства. Цьому сприяють: 1. випадкове злиття яйцеклітини і сперматозоїда під час запліднення, 2. кросинговер у профазі першого поділу мейозу, 3. незалежне розходження гомологічних хромосом в анафазі першого поділу мейозу. |
Источник: narodna-osvita.com.ua