Наличие целлюлозы в клеточной стенке
КЛЕТОЧНАЯ СТЕНКА, клеточная оболочка одноклеточных, колониальных и многоклеточных грибов и растений. У многоклеточных растений все клеточные стенки и межклетники образуют единую общеорганизменную структуру (апопласт), обеспечивающую передвижение веществ от клетки к клетке. Толщина клеточной стенки варьирует у разных клеток (у растительных, например, от 0,1 мкм до многих десятков мкм). Формирующие её вещества представлены преимущественно полисахаридами. Они образуют матрикс (аморфное вещество) и включённые в него тонкие нити разного размера – микро и макрофибриллы, которые практически нерастяжимы и обеспечивают прочность клеточной стенки на разрыв. Такие микро и макрофибриллы сложены параллельными линейными неразветвлёнными молекулами хитина (у большинства грибов), целлюлозы (у некоторых грибов и большинства растений), ксиланов (у ряда зелёных и красных водорослей), маннанов (у некоторых зелёных водорослей). На их долю приходится от «4% до «90% сухой массы клеточной стенки, что зависит от таксономической принадлежности организма, типа ткани и возраста клетки. Основу матрикса составляют разветвлённые и неразветвлённые молекулы пептидогликанов, маннанов (грибы), пектинов и гемицеллюлоз (высшие растения), альгинатов и фукоидана (бурые водоросли), которые соединены различными связями в ажурную трёхмерную сеть, проницаемую для водных растворов. Матрикс включает также белки (в том числе гликопротеины), роль которых в клеточных стенках плохо изучена.
Образование клеточной стенки начинается с отложения на поверхности плазматической мембраны слоя матрикса, биополимеры которого синтезируются внутри клетки и путём экзоцитоза выделяются на её поверхность. Синтез микро и макрофибрилл происходит из выделяемых протопластом мономерных предшественников с помощью подвижных белковых комплексов. Последние пронизывают насквозь мембрану и связаны с подстилающими её микротрубочками цитоскелета. У одноклеточных и колониальных организмов изначальный слой матрикса лишён микро и макрофибрилл, часто ослизняется; у спор он инкрустируется минеральными веществами и/или полифенолами либо замещается так называемым спорополленином; у многоклеточных организмов изначальный матрикс становится межклеточной пластинкой. В дальнейшем протопласт формирует собственно клеточные стенки, матрикс которой является непосредственным продолжением изначального слоя. У грибов в толще клеточной стенки образуется срединный слой из пептидогликанов без фибрилл. У растений различают первичную и вторичную клеточные стенки.
рвая образуется во время роста клетки и завершается вскоре после его окончания; содержит небольшое число беспорядочно ориентированных микро и макрофибрилл, что позволяет стенке растягиваться по мере клеточного роста. Клетки некоторых типов высших растений на первичной клеточной стенке формируют изнутри вторичную клеточную стенку с микро и макрофибриллами, располагающимися плотно, параллельно друг другу, с хорошо выраженной слоистой структурой. Фибриллы различных слоёв вторичной клеточной стенки ориентированы в разных направлениях, вследствие чего она становится нерастяжимой. Клеточные стенки могут подвергаться обызвествлению (например, у «известковых» зелёных и красных водорослей), окремнению (например, у хвощей), одревеснению (у сосудистых растений), опробковению (у высших растений), замещению пектиново-гемицеллюлозного матрикса каллозой (в ситовидных полях элементов флоэмы).
Клеточная стенка выполняет функции внешнего скелета клетки, защитной оболочки, регулятора водного режима, участвует в обменных процессах, передаче сигналов внутрь клетки из окружающей среды. Гемицеллюлозы клеточной стенки в эндосперме семян являются запасным веществом для зародыша (хурма, пальмы). Некоторые клетки высших растений в зрелом состоянии лишаются протопласта и представлены исключительно клеточной стенкой (например, клетки феллемы, проводящие элементы ксилемы); в основном из клеточной стенки отмерших клеток состоит древесина. Фактически именно клеточные стенки служат сырьём для деревообрабатывающей и целлюлозно-бумажной промышленности, производства вискозы, а также получения агара и альгинатов для медицинской, пищевой и косметической промышленности. О клеточной стенке бактерий смотри в статьях Бактерии, Грамотрицательные и грамположительные бактерии.
Лит.: Ботаника с основами фитоценологии: анатомия и морфология растений. М., 2007.
А. К. Тимонин.
Источник: knowledge.su
Клеточная стенка бактерий — это тонкая бесцветная структура, покрывающая клетку снаружи. У большинства бактерий она невидима в обыкновенный микроскоп без специальной обработки. Однако у крупных форм, например у серобактерии Beg. mirabilis, стенка заметна отчетливо. При явлении плазмолиза, который наступает при помещении клеток в 1-2 %-ный гипертонический раствор NaCl или раствор глюкозы, контуры стенки приобретают четкость и она хорошо видна при фазово-контрастной микроскопии.
Стенка бактериальной клетки составляет до 50 % сухой массы организма, толщина ее колеблется в пределах 20-80 нм. Клеточная стенка — плотная ригидная структура. Она обладает эластичностью и достаточной механической прочностью, выдерживает внутриклеточное осмотическое давление, достигающее 10-30 атм.
Химический состав клеточных стенок различных видов бактерий неодинаков, довольно сложен и отличает их не только от клеток растений и животных, но и друг от друга.
Основным компонентом клеточной оболочки высших растений и водорослей является целлюлоза.
целлюлозы состоят, например, микрофибриллы большинства водорослей — до 50-80 % сухой массы оболочки клетки. В микрофибриллах клеточных оболочек большинства мицелиальных грибов преобладает хитин — полимер N-ацетилглюкозамина.
Совершенно иной химический состав имеют клеточные стенки бактерий. Такие соединения как целлюлоза и хитин не характерны для них. Правда, некоторые виды бактерий способны синтезировать целлюлозу и компоненты хитина. Так, у Sarcina ventriculi целлюлоза составляет толстый внешний слой клеточной стенки. Помимо Acetobacter xylinum, это единственный представитель прокариот, синтезирующий данный полимер. Компонент хитина ацетилглюкозамин обнаруживается у всех видов бактерий, за исключением некоторых архебакгерий.
В клеточных стенках бактерий содержится два класса новых, необычных соединений, присущих только прокариотам. Это пептидогликан и тейхоевые кислоты.
Пептидогликаны и тейхоевые кислоты. Пептидогликан, или муреин (от лат. myrus — стенка) представляет собой гетерополимер, состоящий из цепочек чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты (эфир молочной кислоты и N-ацетилглюкозамина), соединенных р-1,4-гликозидной связью. К карбоксильной группе мурамовой кислоты присоединен пептид, включающий чаще всего четыре аминокислоты — тетрапептид. Аминокислотный состав пептида различных видов бактерий не одинаков: у Staph, aureus содержится а-лизин, у Е. сой — мезо-диаминопимелиновая кислота, у Corynebacterium — 2-4-диаминомасляная (рис. 3.11).

Рис. 3.11. Структура пептидогликана стафилококка:
1- N-ацетилмурамовая кислота; 2 — N-ацетилглюко- замин; 3 — тетрапептид; 4 — глидоновый мостик
На основании аминокислотного состава пептидов и соединяющих их мостиков различают ряд подгрупп пептидогликана. Особенностью пептидной части этого полимера является наличие D-аминокислот (в белках они не встречаются) и высокое содержание диаминокислот. Обе аминогруппы, входящие в состав муреина диаминокислоты, участвуют в образовании пептидных связей — с D-аланином и аминокислотным мостиком. Посредством мостиков осуществляются поперечные сшивки пептидогликановых цепей. В итоге формируется гигантская молекула, напоминающая по виду мешок, состоящая из сети полисахаридных цепей, связанных множеством поперечных пептидных связей. За счет образования поперечных сшивок обеспечивается жесткая трехмерная пространственная организация молекулы, обусловливающая механическую прочность и ригидность клеточной стенки.
Пептидогликан чувствителен к литическому действию лизоцима, который расщепляет p-l-4-гликозидные связи между N-ацетилглюкозамином и N-ацетилмурамовой кислотой. Обработка бактерий лизоцимом приводит к разрушению сформированной клеточной стенки.
гибитором синтеза пептидогликана является ряд антибиотиков: пенициллин, цефалоспорин, бацитрацин,
ванкомицин. К примеру, пенициллин подавляет активность фермента транспептидазы, катализирующего образование поперечных сшивок между образующимися цепями пептидогликана. Не сшитый полимер не используется для образования клеточной стенки бактерий.
Тейхоевые кислоты (от греч. «тейхос» — стенка) представляют собой растворимые в воде полимеры, состоящие из остатков трехатомного спирта глицерола или пятиатомного — рибитола, которые соединены друг с другом фосфодиэфирными связями (рис. 3.12). Цепи тейхоевых кислот могут содержать от 10 до 50 остатков спирта. Большинство тейхоевых кислот включают значительное количество D-аланина, аминогруппы которого придают тейхоевым кислотам амфотерные свойства. Кроме D-аланина свободные гидроксильные группы спиртов могут быть замещены глюкозой, N-ацетилглюкозамином, галактозой. Наличие свободных гидро-
ксилов фосфорной кислоты обусловливает сродство тейхоевых кис лот к двухвалентным катионам.
Рис.3.12. Структура тейхоевых кислот клеточной стенки:
а -глицеролтейхоевая; б — рибитолтеихоевая
Клетки одного штамма бактерий, как правило, содержат тейхоевую кислоту только одного типа: рибитолтейхоевую или глицеролтейхоевую.
и уникальные соединения содержатся в клеточных стенках только грамположительных бактерий, где они прочно связаны с пептидогликаном. Так как тейхоевые кислоты представляют собой длинные линейные молекулы, они могут проходить через весь пептидогликановый слой до наружной части клетки и играть роль поверхностных антигенов, обусловливая, таким образом, антигенную специфичность клеточной поверхности бактерий. Кроме того, создавая в клеточной стенке высокую плотность строго ориентированных зарядов, тейхоевые кислоты оказывают влияние на проникновение ионов в клетку, обеспечивая высокую плотность двухвалентных катионов в области цитоплазматической мембраны. Это благоприятствует поддержанию физической целостности мембраны и ее связи с рибосомами.
У некоторых бактерий тейхоевые кислоты участвуют в регуляции активности автолитических ферментов, осуществляющих
в определенных условиях гидролиз муреина собственной клетки. Так, у пневмококков тейхоевые кислоты ингибируют действие литических ферментов клетки путем связывания с ними. Нарушение этой связи приводит клетки к лизису.
Пептидогликан является основным структурным компонентом клеточных стенок почти всех прокариот, за исключением архебактерий, у которых он либо совсем отсутствует, либо имеет иной химический состав. Например, у метанобразующих бактерий пептидогликан содержит вместо муреиновой кислоты талозоминуроновую, а пептидная часть не содержит D-аминокислот, состоит только из а-форм.
В зависимости от химического состава и структуры клеточной стенки все бактерии разделяют на грамположительные и грамотрицательные. Это основано на способности их окрашиваться фиолетовыми красителями трифенилметанового ряда кристаллвиолетом или генцианвиолетом — и не обесцвечиваться нейтральными растворителями — спиртом, ацетоном. Этот метод окраски введен впервые в 1884 г. датским врачом Христианом Грамом и окраска по Граму используется как важнейший таксономический признак бактерий. Сущность его состоит в следующем. Фиксированные клетки окрашиваются кристаллвиолетом или генцианвиолетом, затем протравливаются 30 с раствором Люголя (1 + KI), промываются спиртом, водой и докрашиваются 1 %-ным водным фуксином. Грамположительные бактерии приобретают синий цвет, грамотрицательные — красный.
По структуре и химическому составу клеточной стенки грамположительные бактерии существенно отличаются от грамотрицательных ( табл. 2).
У грамположительных бактерий клеточная стенка представляет собой гомогенный электронно-плотный слой толщиной 20 — 80 нм. Основную массу (50-90 % сухого вещества) составляет пептидогликан, образующий ригидный толстый слой. Он плотно прилегает к ЦПМ. Пептидогликановый слой пронизан тейхоевыми кислотами, которые могут выходить на поверхность клеточной стенки. Кроме этих основных полимеров в клеточных стенках грамположительных бактерий содержатся в небольших количествах липиды, полисахариды, белки. Липиды и
полисахариды ковалентно связываются с пептидогликаном, образуя сложную, механически прочную структуру.
Таблица 2
Характеристика химического состава клеточных стенок бактерий
| Компоненты | Г рамположительные | Г рамотрицател ьные |
| Пептидогликан | + | + |
| Тейхоевые кислоты | + | — |
| Липиды | + | + |
| Белки | ± | + |
| Полисахариды | + | — |
| Липополисахариды | — | + |
| Липопротеиды | — | + |
точная стенка грамотрицательных бактерий более тонкая (10-15 нм ) и многослойная (рис.3.13). Внутренний слой представлен пептидогликаном, содержание которого значительно меньше (1-10 %), чем в стенках грамположительных бактерий. Толщина данного слоя 2-3 нм. Наружный слой более рыхлый и толстый — 8-10 нм, имеет сложный химический состав. В нем обнаружены белки, фосфолипиды и липополисахариды, расположенные мозаично. По структуре и химическому составу этот слой имеет сходство с цитоплазматической мембраной.
получил название наружной мембраны и имеется только у грамотрицательных бактерий.
Наружная мембрана является дополнительным барьером, препятствующим проникновению в клетку крупных молекул. Так, она препятствует поступлению в клетку антибиотиков, в частности пенициллина, актиномицина Д. Вполне возможно, что по этой причине грамотрицательные бактерии менее чувствительны к антибиотикам, чем грамположительные.
Липополисахариды наружной мембраны определяют антигенную специфичность бактерий, а также служат рецепторами для адсорбции фагов.
Белки наружной мембраны выполняют разные функции. Одни из них, так называемые белки матриксапорины, формируют в
мембране гидрофильные поры, через которые осуществляется диффузия аминокислот, небольших олигосахаридов и пептидов (молекулярная масса от 600 до 900 Да6). Транспорт веществ через поры, образованные поринами, лишен специфичности. Порины являются также рецепторами для фагов и колицинов.
Вторая группа белков — минорные белки, как и предыдущая группа, выполняют транспортные и рецепторные функции. Важная роль отводится им в транспорте железосодержащих соединений в клетке разных видов грамотрицательных бактерий.
Таким образом, структура клеточной стенки грамотрицательных бактерий намного сложнее, чем грамположительных. Структурные особенности и химический состав клеточных стенок лежат в основе механизма окрашиваемости бактерий по Г раму.
Да — дальтон, или единица аггомной массы, равен 1,66033 х 10 1 кг.
Ответственность за окраску по Граму несут муреин и частично липиды, оказывающие влияние на проницаемость клеточной стенки. Обработка бактерий спиртом вызывает разбухание муреина и уменьшение диаметра пор клеточной стенки, что в целом приводит к снижению ее проницаемости. Так как грамположительные бактерии характеризуются высоким содержанием муреина, то в результате обработки спиртом стенки их становятся почти непроницаемыми для красителей и вымывание краски не происходит. У грамотрицательных слой муреина тонкий и не играет существенной роли в проницаемости стенки. Кроме того, проницаемость клеточной стенки у грамположительных бактерий увеличивается за счет растворения и вымывания липидов спиртом, содержание которых довольно высокое ( до 22 % ), и к тому же они хорошо растворяются в нейтральных органических растворителях. Все это способствует обесцвечиванию клетки. Доказательством того, что в окраске по Граму основную роль играет клеточная стенка, является тот факт, что при удалении ее с окрашенных клеток протопласты грамположительных бактерий при промывании спиртом обесцвечиваются, превращаясь в грамотрицательные. Следовательно, окрашенный комплекс удерживает клеточная стенка.
Клеточная стенка у грамотрицательных бактерий отделена от цитоплазматической мембраны электронно-прозрачным промежутком, получившим название периплазматического пространства, или периплазма. В нем содержатся кроме тонкого слоя муреина (2-3 нм) специфические белки, так называемые связывающие, или транспортные белки. Это водорастворимые белки, обладающие высоким сродством к определенным питательным субстратам — аминокислотам, сахарам, неорганическим ионам. Они являются составной частью систем активного транспорта, но самостоятельно осуществлять этот процесс они не могут и функционируют только в сочетании со специфическими пермеазами, локализованными в цитоплазматической мембране. Транспортные белки связывают соответствующие субстраты и переносят их от внешней мембраны к цитоплазматической. В периплазматическом пространстве содержится также ряд гидролитических ферментов — нуклеазы, щелочная и кислая фосфатазы, пенициллианаза. У грамположительных бактерий эти ферменты являются типичными экзоферментами, у грамотрицательных выход их из клеток задерживается наружной мембраной, которая является барьером для белков и некоторых других соединений. Наличие в периплазме ферментов позволяет клетке использовать более широкий круг веществ, поступающих извне. Так как данные ферменты изолированы от цитоплазмы, то содержание их не угрожает содержимому клетки подвергнуться автолизу, или самоперевариванию.
Важнейшие функции клеточной стенки заключаются в следующем. Она обеспечивает клетке определенное постоянство формы, защищает содержимое от ее внешних воздействий, определяет способность к адсорбции фагов, так как на ее поверхности расположены фагочувствительные рецепторы, играет важную роль и в реакции иммунитета. Установлено, что между фагоцитарной активностью лейкоцитов и поверхностной структурой бактериальных клеток существует определенная зависимость. Особенности структуры клеточной стенки определяют чувствительность бактерий к повреждающему действию сыворотки крови и форменных элементов.
Таким образом, клеточная стенка бактерий — сложная полифункциональная система, обладающая необходимыми реологическими свойствами (упругость, пластичность, прочность) и обеспечивающая анатомическую целостность клетки, геометрическую форму ее и контакт с внешней средой.
Источник: www.med24info.com
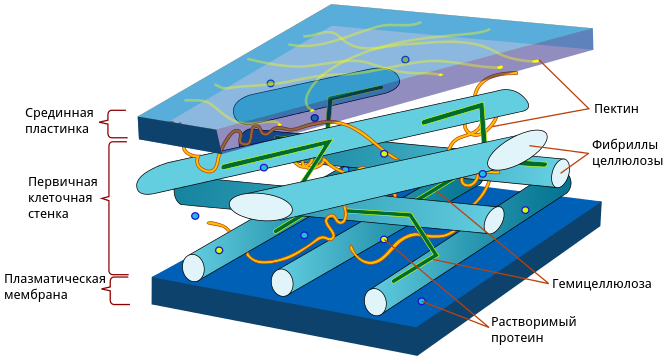
Состав клеточной стенки растений
У растений стенка дочерних клеток образуется уже во время деления родительской. Впоследствии она называется первичной. У многих клеток позже образуется вторичная оболочка.
Первичная клеточная оболочка состоит из микрофибрилл целлюлозы, погруженных в матрикс из других полисахаридов. Отличительной особенностью волокон целлюлозы является их прочность. Молекула целлюлозы представляет собой длинную полисахаридную цепь. Отдельные молекулы соединяются друг с другом водородными связями в пучок, который называется микрофибриллой. Такие фибриллы образуют каркас клеточной стенки.
Матрикс клеточной стенки составляют полисахариды пектины и гемицеллюлозы, а также ряд других веществ (например, белков). Пектиновые вещества представляют собой группу кислых полисахаридов, их молекулы могут быть не только линейными, но и разветвленными. Гемицеллюлозы также смешанная группа полисахаридов. Длина их линейных молекул короче, чем у целлюлозы.
Оболочки соседних клеток растений соединены между собой срединной пластинкой, состоящих из пектатов магния и кальция, для которых характерна клейкость.
В состав стенок растений входит вода (составляет более половины массы), обуславливая ряд физических и химических свойств полисахаридов.
Жесткий каркас растения во многих местах пронизан каналами (плазмодесмами), по которым цитоплазма одной клетки соединяется с цитоплазмой соседних.
Клетки мезофилла листа (а также некоторые другие) на протяжении всей своей жизни имеют только первичную стенку. У большинства же клеток на первичную оболочку с внутренней стороны отлагается вторичная стенка, составленная из дополнительных слоев целлюлозы. Обычно в это время клетка уже дифференцирована и не растет (исключение составляют, например, клетки колленхимы).
В каждом отдельном слое вторичного утолщения микрофибриллы целлюлозы располагаются под одним углом (параллельно друг другу). Однако разные слои имеют разный угол, что обеспечивает большую прочность.
Часть клеток растений одревесневают (трахеальные элементы ксилемы, склеренхима и др.). В основе этого процесса лежит интенсивная лигнификация стенок (в небольших количествах лигнин есть во всех оболочках). Лигнин не является полисахаридом, а представляет собой сложное полимерное вещество. Отложения лигнина могут иметь различную форму (сплошную, кольцевую, спиральную, сетчатую). Он скрепляет целлюлозу, не дает ей смещаться. Лигнин не только обеспечивает прочность, но и дает дополнительную защиту от неблагоприятных физических и химических факторов.
Функции клеточной стенки
Оболочки разных клеток совместно обеспечивают всему растению и его отдельным частям механическую прочность и опору. Это функция клеточной стенки аналогична одной из функций скелета животных. Однако она не единственная.
Жесткость стенок препятствует растяжению клеток и их разрыву. В результате по физическим законам в клетки может путем осмоса поступать вода. Для травянистых растений тургоцентричность клеток является единственной их опорой.
Микрофибриллы целлюлозы ограничивают рост клеток и определяют их форму. Если микрофибриллы окольцовывают клетку, то она будет расти в длину (поперек направления волокон).
Связанные клеточные стенки образуют апопласт, по которому передвигается вода и минеральные вещества. Плазмодесмы связывают содержимое разных клеток в единую систему — симпласт.
Стенки сосудов ксилемы, трахеид, ситовидных трубок выполняют транспортную функцию.
Наружные клеточные стенки эпидермальных клеток покрыты воском (кутикулой). С одной стороны, он препятствует испарению воды, с другой – проникновению вредных микроорганизмов.
У некоторых растений в определенных клетках оболочки видоизменяются и служат местом запаса питательных веществ.
Источник: biology.su