Определение мейоз
Мейоз – это деление, при котором получаются половые клетки (у растений – споры). Биологическое значение мейоза:
- рекомбинация (перемешивание наследственной информации)
- редукция (уменьшение количества хромосом в 2 раза).
Отличия мейоза от митоза по итогам
1. После митоза получается две клетки, а после мейоза – четыре.
2. После митоза получаются соматические клетки (клетки тела), а после мейоза – половые клетки (гаметы – сперматозоиды и яйцеклетки; у растений после мейоза получаются споры).
3. После митоза получаются одинаковые клетки (копии), а после мейоза – разные (происходит рекомбинация наследственной информации).
4. После митоза количество хромосом в дочерних клетках остается таким же, как было в материнской, а после мейоза уменьшается в 2 раза (происходит редукция числа хромосом; если бы её не было, то после каждого оплодотворения число хромосом возрастало бы в два раза; чередование редукции и оплодотворения обеспечивает постоянство числа хромосом).
Отличия мейоза от митоза по ходу
1. В митозе одно деление, а в мейозе – два (из-за этого получается 4 клетки).
2. В профазе первого деления мейоза происходит конъюгация (тесное сближение гомологичных хромосом) и кроссинговер (обмен участками гомологичных хромосом), это приводит к перекомбинации (рекомбинации) наследственной информации.
3. В анафазе первого деления мейоза происходит независимое расхождение гомологичных хромосом (к полюсам клетки расходятся двуххроматидные хромосомы). Это приводит к рекомбинации и редукции.
4. В интерфазе между двумя делениями мейоза удвоения хромосом не происходит, поскольку они и так двойные.
…
Второе деление мейоза ничем не отличается от митоза. Как и в митозе, в анафазе II мейоза к полюсам клетки расходятся одинарные сестринские хромосомы (бывшие хроматиды). (сайт)
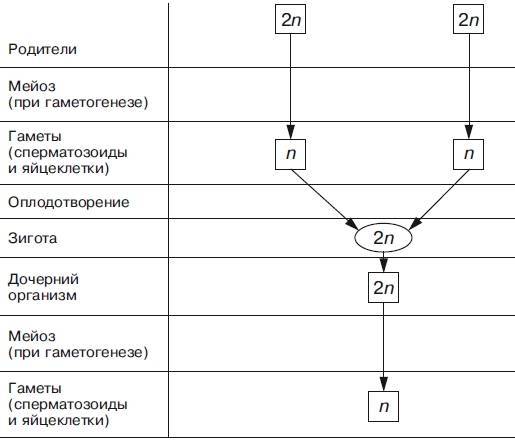
При половом размножении дочерний организм возникает в результате слияния двух половых клеток (гамет) и последующего развития из оплодотворенной яйцеклетки — зиготы.
Половые клетки родителей обладают гаплоидным набором (n) хромосом, а в зиготе при объединении двух таких наборов число хромосом становится диплоидным (2n): каждая пара гомологичных хромосом содержит одну отцовскую и одну материнскую хромосому.
Гаплоидные клетки образуются из диплоидных в результате особого клеточного деления — мейоза.
Мейоз — разновидность митоза, в результате которого из диплоидных (2п) соматических клеток половых желез образуются гаплоидные гаметы (1n). При оплодотворении ядра гаметы сливаются, и восстанавливается диплоидный набор хромосом. Таким образом, мейоз обеспечивает сохранение постоянного для каждого вида набора хромосом и количества ДНК.
Мейоз представляет собой непрерывный процесс, состоящий из двух последовательных делений, называемых мейозом I и мейозом II. В каждом делении различают профазу, метафазу, анафазу и телофазу. В результате мейоза I число хромосом уменьшается вдвое (редукционное деление): при мейозе II гаплоидность клеток сохраняется (эквационное деление). Клетки, вступающие в мейоз, содержат генетическую информацию 2n2хр (рис. 1).
В профазе мейоза I происходит постепенная спирализация хроматина с образованием хромосом. Гомологичные хромосомы сближаются, образуя общую структуру, состоящую из двух хромосом (бивалент) и четырех хроматид (тетрада). Соприкосновение двух гомологичных хромосом по всей длине называется конъюгацией. Затем между гомологичными хромосомами появляются силы отталкивания, и хромосомы сначала разделяются в области центромер, оставаясь соединенными в области плеч, и образуют перекресты (хиазмы). Расхождение хроматид постепенно увеличивается, и перекресты смещаются к их концам. В процессе конъюгации между некоторыми хроматидами гомологичных хромосом может происходить обмен участками — кроссинговер, приводящий к перекомбинации генетического материала. К концу профазы растворяются ядерная оболочка и ядрышки, формируется ахроматиновое веретено деления. Содержание генетического материала остается прежним (2n2хр).
В метафазе мейоза I биваленты хромосом располагаются в экваториальной плоскости клетки. В этот момент спирализация их достигает максимума. Содержание генетического материала не изменяется (2п2хр).
В анафазе мейоза I гомологичные хромосомы, состоящие из двух хроматид, окончательно отходят друг от друга и расходятся к полюсам клетки. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна — число хромосом уменьшается вдвое (происходит редукция). Содержание генетического материала становится 1n2хр у каждого полюса.
В телофазе происходит формирование ядер и разделение цитоплазмы — образуются две дочерние клетки. Дочерние клетки содержат гаплоидный набор хромосом, каждая хромосома — две хроматиды (1n2хр).
Интеркинез — короткий промежуток между первым и вторым мейотическими делениями. В это время не происходит репликации ДНК, и две дочерние клетки быстро вступают в мейоз II, протекающий по типу митоза.

Рис. 1. Схема мейоза (показана одна пара гомологичных хромосом). Мейоз I: 1, 2, 3. 4. 5 — профаза; 6 —метафаза; 7 — анафаза; 8 — телофаза; 9 — интеркинез. Мейоз II; 10 —метафаза; II —анафаза; 12 — дочерние клетки.
В профазе мейоза II происходят тс же процессы, что и в профазе митоза. В метафазе хромосомы располагаются в экваториальной плоскости. Изменений содержания генетического материала не происходит (1n2хр). В анафазе мейоза II хроматиды каждой хромосомы отходят к противоположным полюсам клетки, и содержание генетического метериала у каждого полюса становится lnlxp. В телофазе образуются 4 гаплоидные клетки (lnlxp).
Таким образом, в результате мейоза из одной диплоидной материнской клетки образуются 4 клетки с гаплоидным набором хромосом. Кроме того, в профазе мейоза I происходит перекомбинация генетического материала (кроссинговер), а в анафазе I и II — случайное отхождение хромосом и хроматид к одному или другому полюсу. Эти процессы являются причиной комбинативной изменчивости.
Биологическое значение мейоза:
1) является основным этапом гаметогенеза;
2) обеспечивает передачу генетической информации от организма к организму при половом размножении;
3) дочерние клетки генетически не идентичны материнской и между собой.
Атак же, биологическое значение мейоза заключается в том, что уменьшение числа хромосом необходимо при образовании половых клеток, поскольку при оплодотворении ядра гамет сливаются. Если бы указанной редукции не происходило, то в зиготе (следовательно, и во всех клетках дочернего организма) хромосом становилось бы вдвое больше. Однако это противоречит правилу постоянства числа хромосом. Благодаря мейозу половые клетки гаплоидны, а при оплодотворении в зиготе восстанавливается диплоидный набор хромосом (рис. 2 и 3).
Рис. 2. Схема гаметогенеза: à — сперматогенез; á — овогенез
Рис. 3. Схема, иллюстрирующая механизм сохранения диплоидного набора хромосом при половом размножении
Источник: www.sites.google.com
Первое мейотическое деление
В результате первого мейотического (редукционного) деления из диплоидных клеток (2n) образуются гаплоидные (n). Он начинается из профазы І, в которой, также как и в митозе, происходит упаковка наследственного материала (спирализация хромосом). Одновременно гомологические (парные) хромосомы сближаются одинаковыми участками – происходит коньюгация. В результате коньюгации образуются пары хромосом – биваленты.
ждая хромосома, вошедшая в мейоз, состоит из двух хроматид иимеет удвоенный наследственный материал, потому бивалент состоит из 4 нитей. Когда хромосомы находятся в коньюгированном состоянии, их дальнейшая спирализация продолжается. Отдельные хроматиды гомологических хромосом переплетаются и перекрещиваются. В дальнейшем гомологические хромосомы отталкиваются и немного расходятся, потому в местах переплетения хроматид может произойти их разрыв. Как результат в процессе возобновления разрывов у хроматиды гомологических хромосом происходит обмен соответствующими участками. В итоге перешедшая от родителя к данному организму хромосома, содержит часть материнской хромосомы, и наоборот.
После кроссинговера уже изменённые хромосомы, то есть с другими объединениями генов, расходятся.
Поскольку кроссинговер является процессом закономерным, он каждый раз приводит к обмену разными по размеру участками и, таким образом, обеспечивается эффективная рекомбинация материала хромосом гамет.
- В метафазе І завершает формироваться веретено деления. Его нити крепятся к центромерам хромосом, которые соединены в биваленты так, что от каждой центромеры отходит лишь одна нить к одному из полюсов клетки. В результате с помощью связанных с центромерами гомологических хромосом нитей биваленты располагаются по экватору веретена деления.
- В анафазе І гомологические хромосомы рассоединяются и расходятся к полюсам клетки.
В телофазе І возле полюсов веретена собирается одинарный (гаплоидный) набор хромосом, в котором каждый их вид представлен уже не парой, а одной хромосомой, состоящей из двух хроматид. В короткой по длительности телофазе возобновляется оболочка ядра, а материнская клетка делится на две дочерние. Таким образом, благодаря образованию бивалентов при коньюгации гомолологических хромосом в профазе І мейоза создаёт условия для дальнейшей редукции количества хромосом. Формируется гаплоидный набор в гаметах, который обеспечивается расхождением в анафазе І не хроматид, как в митозе, а гомологических хромосом, которые ранее были соединены в биваленты.
Второе мейотическое деление
Второе мейотическое деление происходит сразу же после первого и подобно обычному митозу (потому его ещё называют митозом мейоза), но клетки, которые делятся, содержат гаплоидный набор хромосом.
- Профаза ІІ недлительна.
- В метафазе ІІ снова образуется веретено деления, хромосомы располагаются в экваториальной плоскости, а центромеры соединяются с микротрубочками веретена деления.
- В анафазе ІІ их центромеры рассоединяются и каждая хроматида превращается в самостоятельную хромосому. Дочерние хромосомы, которые отделяются друг от друга, направляются к полюсам клетки.
- В телофазе ІІ завершается расхождение хромосом и клетки делятся: из двух гаплоидных клеток образуются четыре гаплоидные дочерние клетки.
Значение мейоза
Благодаря редукционному делению регулируется непрерывное увеличение числа хромосом в процессе слиянии гамет. Если бы не было этого механизма, то во время полового размножения число хромосом удваивалось бы из поколения в поколение.
Ещё одно важное значение мейоза: обеспечение большого разнообразия генетического состава гамет как в результате кроссинговера, так и в результате различного объединения отцовских и материнских хромосом при их расхождении в анафазе І мейоза. Это обеспечивает разнообразие и разнокачественность потомства во время полового размножения.
Источник: spravochnick.ru
(от греч. méiosis ≈ уменьшение), редукционное деление, деления созревания, способ деления клеток, в результате которого происходит уменьшение (редукция) числа хромосом в два раза и одна диплоидная клетка (содержащая два набора хромосом) после двух быстро следующих друг за другом делений даёт начало 4 гаплоидным (содержащим по одному набору хромосом). Восстановление диплоидного числа хромосом происходит в результате оплодотворения. М. ≈ обязательное звено полового процесса и условие формирования половых клеток ( гамет ). Биологическое значение М. заключается в поддержании постоянства кариотипа в ряду поколений организмов данного вида и обеспечении возможности рекомбинации хромосом и генов при половом процессе. М. ≈ один из ключевых механизмов наследственности и наследственной изменчивости . Поведение хромосом при М. обеспечивает выполнение основных законов наследственности (см. Менделя законы ).
В зависимости от места М. в жизненном цикле организмов различают 3 типа М. Гаметный, или терминальный, М. (у всех многоклеточных животных и ряда низших растений), происходит в половых органах и приводит к образованию гамет. Зиготный, или начальный, М. (у многих грибов и водорослей), происходит в зиготе сразу после оплодотворения и приводит к образованию гаплоидного мицелия или таллома , а затем спор и гамет. Споровый, или промежуточный, М. (у высших растений), имеет место накануне цветения и приводит к образованию гаплоидного гаметофита , в котором позднее образуются гаметы. У простейших (Protozoa) встречаются все 3 типа М. Перед М. происходит удвоение количества ДНК в клетке. В ходе двух делений М. (рис.) ДНК делится поровну между 4 клетками. В результате первого (редукционного) деления М. пары гомологичных хромосом разъединяются и члены пар расходятся в 2 клетки (редукция числа хромосом). Каждая хромосома сохраняет две продольные половины ≈ хроматиды . В результате второго (эквационного) деления хроматиды расходятся в разные клетки и каждая из 4 сестринских клеток получает по одной хроматиде. Т. о., первое деление М. принципиально отличается от митоза , а второе ≈ это митоз в клетках с гаплоидным числом хромосом. Во время М., перед редукцией числа хромосом, происходит обмен участками гомологичных хромосом ≈ кроссинговер , приводящий к перераспределению аллельных генов (см. Аллели ). М. длится много дольше митоза: например, у пшеницы он продолжается 24 часа, у лилии ≈ 9≈12 суток, у мыши ≈ 11≈14 суток, у человека ≈ 24. У ряда животных и человека во время образования женских половых клеток (см. Оогенез ) М. останавливается на срок до нескольких лет и завершается только во время оплодотворения.
Первая фаза М. ≈ профаза I, наиболее сложная и длительная (у человека 22,5, у лилии 8≈10 суток), подразделяется на 5 стадий. Лептотена ≈ стадия тонких нитей, когда хромосомы слабо спирализованы и наиболее длинны, видны утолщения ≈ хромомеры. Зиготена ≈ стадия начала попарного, бок о бок соединения (синапсиса, конъюгации) гомологичных хромосом; при этом гомологичные хромомеры взаимно притягиваются и выстраиваются строго друг против друга. Пахитена ≈ стадия толстых нитей; гомологичные хромосомы стабильно соединены в пары ≈ биваленты, число которых равно гаплоидному числу хромосом; под электронным микроскопом видна сложная ультраструктура в месте контакта двух гомологичных хромосом внутри бивалента: т. н. синаптонемальный комплекс, который начинает формироваться ещё в зиготене; в каждой хромосоме бивалента обнаруживаются 2 хроматиды; т. о., бивалент (тетрада, по старой терминологии) состоит из 4 гомологичных хроматид; на этой стадии происходит кроссинговер, осуществляющийся на молекулярном уровне; цитологические последствия его обнаруживаются на следующей стадии. Диплотена ≈ стадия раздвоившихся нитей; гомологичные хромосомы начинают отталкиваться друг от друга, но оказываются связанными, обычно в 2≈3 точках на бивалент, где видны хиазмы (перекресты хроматид) ≈ цитологическое проявление кроссинговера. Диакинез ≈ стадия отталкивания гомологичных хромосом, которые по-прежнему соединены в биваленты хиазмами, перемещающимися на концы хромосом (терминализация); хромосомы максимально коротки и толсты (за счёт спирализации) и образуют характерные фигуры: кресты, кольца и др. Следующая фаза М. ≈ метафаза I, во время которой хиазмы ещё сохраняются; биваленты выстраиваются в средней части веретена деления клетки, ориентируясь центромерами гомологичных хромосом к противоположным полюсам веретена. В анафазе I гомологичные хромосомы с помощью нитей веретена расходятся к полюсам; при этом каждая хромосома пары может отойти к любому из двух полюсов, независимо от расхождения хромосом др. пар. Поэтому число возможных сочетаний при расхождении хромосом равно 2n, где n ≈ число пар хромосом. В отличие от анафазы митоза, центромеры хромосом не расщепляются и продолжают скреплять 2 хроматиды в хромосоме, отходящей к полюсу. В телофазе I у каждого полюса начинается деспирализация хромосом и формирование дочерних ядер и клеток. Далее следует короткая интерфаза без редупликации ДНК ≈ интеркинез, и начинается второе деление М. Профаза II, метафаза II, анафаза II и телофаза II проходят быстро; при этом в конце метафазы II расщепляются центромеры, и в анафазе II расходятся к полюсам хроматиды каждой хромосомы. Эта классическая схема М. имеет исключения. Например, у растений рода ожика (Luzula) и насекомых семейства кокцид (Coccidae) в первом делении М. расходятся хроматиды, а во втором ≈ гомологичные хромосомы, однако и в этих случаях в результате М. происходит редукция числа хромосом. Различия между сперматогенезом и оогенезом у животных и образованием микроспор и мегаспор у растений не отражаются на поведении хромосом в ходе М., хотя размеры и судьбы сестринских клеток оказываются разными. Известны аномалии М. У межвидовых гибридов все хромосомы, а у анеуплоидов (см. Анеуплоидия ) непарные хромосомы не способны конъюгировать и остаются в виде унивалентов; у автополиплоидов (см. Автополиплоидия ) образуются объединения более чем из 2 хромосом ≈ т. н. мультиваленты. В каждом из этих случаев невозможна правильная редукция числа хромосом в анафазе I; образующиеся гаметы (с несбалансированными наборами хромосом) либо сами нежизнеспособны, либо дают нежизнеспособное или уродливое потомство (см. Хромосомные болезни ). Отсутствие хиазм (ахизматия) обычно приводит к тем же результатам, однако у самцов некоторых видов мух, в том числе у дрозофилы, хиазмы всегда отсутствуют, хотя гаметы образуются нормальные. Причины перехода клеток от деления путём митоза к М. в жизненном цикле каждого организма, а также молекулярные механизмы конъюгации гомологичных хромосом и кроссинговера исследуются.
Лит.: Соколов И. И., Цитологические основы полового размножения многоклеточных животных, в кн.: Руководство по цитологии, т. 2, М. ≈ Л., 1966, с. 390≈460; Райков И. Б., Карпология простейших, Л., 1967; Богданов Ю. Ф., Гомологичная конъюгация хромосом, в кн.: Успехи современной генетики, т. 3, М., 1971, с. 134≈61; Swanson С. P., Cytology and cytogenetics, Englewood Cliffs, 1957; Rhoades М. М., Meiosis, в кн.: The cell. Biochemistry. Physiology. Morphology, v. 3, N. Y., 1961, p. 1≈75; John B., Lewis K. R., The meiotic system, W. ≈ N. Y., 1965; (Protoplasmatologia, Bd 6, fasc. 1); Westergaard М., Wettstein D. von, The Synaptinemal complex, в кн.: Annual Review of Genetics, v. 6, 1972, p. 71≈110.
Ю. Ф. Богданов.
Источник: xn--b1algemdcsb.xn--p1ai