Органоиды общего назначения
Органоиды – постоянные и обязательные компоненты клеток; специализированные участки цитоплазмы клетки, имеющие определенную структуру и выполняющие определенные функции в клетке. Различают органоиды общего и специального назначения.
Органоиды общего назначения имеются в большинстве клеток (эндоплазматическая сеть, митохондрии, пластиды, комплекс Гольджи, лизосомы, вакуоли, клеточный центр, рибосомы). Органоиды специального назначения характерны только для специализированных клеток (миофибриллы, жгутики, реснички, сократительные и пищеварительные вакуоли). Органоиды (за исключением рибосом и клеточного центра) имеют мембранное строение.
Эндоплазматическая ретикулум(ЭПР) – это разветвленная система соединенных между собой полостей, трубочек и каналов, образованных элементарными мембранами и пронизывающая всю толщу клетки. Открыта в 1943 г. Портером. Особенно много каналов эндоплазматической сети в клетках с интенсивным обменом веществ. В среднем объем ЭПС составляет от 30% до 50% общего объема клетки. ЭПС лабильна. Форма внутренних лакун и кана
лов, их размер, расположение в клетке и количество изменяются в процессе жизнедеятельности. Развита сильнее в животных клетка. ЭПС морфологически и функционально связана с пограничным слоем цитоплазмы, ядерной оболочкой, рибосомами, комплексом Гольджи, вакуолями, образуя вместе с ними единую функционально-структурную систему для осуществления обмена веществ и энергии и передвижения веществ внутри клетки. Вблизи эндоплазматической сети накапливаются митохондрии и пластиды.
Выделяют две разновидности ЭПС: шероховатую и гладкую. На мембранах гладкой (агранулярной) ЭПС локализованы ферменты систем жирового и углеводного синтеза: здесь происходит синтез углеводов и почти всех клеточных липидов. Мембраны гладкой разновидности эндоплазматической сети преобладают в клетках сальных желез, печени (синтез гликогена), в клетках с большим содержанием питательных веществ (семена растений). На мембране шероховатой (гранулярной) ЭПС располагаются рибосомы, где осуществляется биосинтез белков. Часть синтезируемых ими белков включается в состав мембраны эндоплазматической сети, остальные поступают в просвет ее каналов, где преобразуются и транспортируются в комплекс Гольджи. Особенно много шероховатых мембран в клетках желез и нервных клетках.

Рис. Шероховатый и гладкий эндоплазматический ретикулум.
Рис. Транспорт веществ по системе ядро – эндоплазматический ретикулум (ЭПР) – комплекс Гольджи.
Функции эндоплазматической сети:
1) синтез белков (шероховатая ЭПС), углеводов и липидов (гладкая ЭПС);
2) транспорт веществ, как поступивших в клетку, так и вновь синтезируемых;
3) деление цитоплазмы на отсеки (компартменты), что обеспечивает пространственное разделение ферментных систем, необходимое для их последовательного вступления в биохимические реакции.
Митохондрии – присутствуют практически во всех типах клеток одно- и многоклеточных организмов (за исключением эритроцитов млекопитающих). Число их в разных клетках варьирует и зависит от уровня функциональной активности клетки. В клетке печени крысы их около 2500, а в мужской половой клетке некоторых моллюсков – 20 – 22. Их больше в грудной мышце летающих птиц, чем в грудной мышце нелетающих.
Митохондрии имеют форму сферических, овальных и цилиндрических телец. Размеры составляют 0,2 – 1,0 мкм и диаметре и до 5 — 7 мкм в длину.
Рис. Митохондрия.
Длина нитевидных форм достигает 15-20 мкм. Снаружи митохондрии ограничены гладкой наружной мембраной, сходной по составу с плазмалеммой.
утренняя мембрана образует многочисленные выросты – кристы – и содержит многочисленные ферменты, АТФ-сомы (грибовидные тела), участвующие в процессах трансформации энергии питательных веществ в энергию АТФ. Количество крист зависит от функции клетки. В митохондриях мышц крист очень много, они занимают всю внутреннюю полость органоида. В митохондриях эмбриональных клеток кристы единичны. В растительных выросты внутренней мембраны чаще имеют форму трубочек. Полость митохондрии заполнена матриксом, в котором содержатся вода, минеральные соли, белки-ферменты, аминокислоты. Митохондрии имеют автономную белоксинтезирующую систему: кольцевую молекулу ДНК, различные виды РНК и более мелкие, чем в цитоплазме рибосомы.
Митохондрии тесно связаны мембранами эндоплазматической сети, каналы которой часто открываются прямо в митохондрии. При повышении нагрузки на орган и усилении синтетических процессов, требующих затраты энергии, контакты между ЭПС и митохондриями становятся особенно многочисленными. Число митохондрий может быстро увеличиваться путем деления. Способность митохондрий к размножению обусловлена присутствием в них молекулы ДНК, напоминающей кольцевую хромосому бактерий.
Функции митохондрий:
1) синтез универсального источника энергии – АТФ;
2) синтез стероидных гормонов;
3) биосинтез специфических белков.
Пластиды – органоиды мембранного строения, характерные только для растительных клеток. В них происходят процессы синтеза углеводов, белков и жиров. По содержанию пигментов их делят на три группы: хлоропласты, хромопласты и лейкопласты.
Хлоропласты имеют относительно постоянную эллиптическую или линзовидную форму. Размер по наибольшему диаметру составляет 4 – 10 мкм. Количество в клетке колеблется от нескольких единиц до нескольких десятков. Их размер, интенсивность окраски, количество и расположение в клетке зависят от условий освещения, вида и физиологического состояния растений.
Рис. Хлоропласт, строение.
Это белково-липоидные тела, состоящие на 35-55% из белка, 20-30% — липидов, 9% — хлорофилла, 4-5% каратиноидов, 2-4% нуклеиновых кислот. Количество углеводов варьирует; обнаружено некоторое количество минеральных веществ Хлорофилл – сложный эфир органической двухосновной кислоты – хлорофиллина и органических спиртов – метилового (СН3ОН) и фитола (С20Н39ОН). У высших растений в хлоропластах постоянно присутствуют хлорофилл а – имеет сине-зеленую окраску, и хлорофилл b – желто-зеленую; причем содержание хлорофилла, а в несколько раз больше.
Кроме хлорофилла в состав хлоропластов входят пигменты — каротин С40Н56 и ксантофилл С40Н56О2 и некоторые другие пигменты (каратиноиды). В зеленом листе желтые спутники хлорофилла маскируются более яркой зеленой окраской. Однако осенью, при листопаде, у большинства растений хлорофилл разрушается и тогда обнаруживается присутствие в листе каратиноида – лист становится желтым.
Хлоропласт одет двойной оболочкой, состоящей из наружной и внутренней мембран. Внутреннее содержимое – строма – имеет ламеллярное (пластинчатое) строение. В бесцветной строме выделяют граны – окрашенные в зеленые цвет тельца, 0,3 – 1,7 мкм. Они представляют собой совокупность тилакоидов – замкнутых телец в виде плоских пузырьков или дисков мембранного происхождения. Хлорофилл в виде мономолекулярного слоя располагается между белковым и липидным слоями в тесной связи с ними. Пространственное расположение молекул пигментов в мембранных структурах хлоропластов является весьма целесообразным и создает оптимальные условия для наиболее эффективного поглощения, передачи и использования лучистой энергии. Липиды образуют безводные диэлектрические слои мембран хлоропласта, необходимые для функционирования электронно-транспортной цепи. Роль звеньев цепи переноса электронов выполняют белки (цитохромы, пластохиноны, ферредоксин, пластоцианин) и отдельные химические элементы – железо, марганец и др. Количество гран в хлоропласте от 20 до 200. Между гранами, связывая их друг с другом, располагаются ламеллы стромы. Ламеллы гран и ламеллы стромы имеют мембранное строение.
Внутренне строение хлоропласта делает возможным пространственное разобщение многочисленных и разнообразных реакций, составляющих в своей совокупности содержание фотосинтеза.
Хлоропласты, как и митохондрии, содержат специфическую РНК и ДНК, а также более мелкие рибосомы и весь молекулярный арсенал, необходимый для биосинтеза белка. У этих органоидов имеется достаточное для обеспечения максимальной активности белоксинтезирующей системы количество и-РНК. Вместе с тем в них содержится и достаточно ДНК для кодирования определенных белков. Они размножаются делением, путем простой перетяжки.
Установлено, что хлоропласты могут изменять свою форму, размеры и положение в клетке, т. е. способны самостоятельно двигаться (таксис хлоропластов). В них обнаружено два типа сократительных белков, за счет которых, очевидно, и осуществляется активное движение этих органоидов в цитоплазме.
Хромопласты широко распространены в генеративных органах растений. Они окрашивают лепестки цветков (лютика, георгина, подсолнечника), плоды (томатов, рябины, шиповника) в желтый, оранжевый, красный цвета. В вегетативных органах хромопласты встречаются значительно реже.
Окраска хромопластов обусловлена присутствием каратиноидов – каротина, ксантофилла и ликопина, которые в пластидах находятся в различном состоянии: в виде кристаллов, липоидного раствора или в соединении с белками.
Хромопласты, по сравнению с хлоропластами, имеют более простое строение – в них отсутствует ламеллярная структура. Химический состав также отличен: пигменты – 20–50%, липиды до 50%, белки – около 20%, РНК – 2-3%. Это свидетельствует о меньшей физиологической активности хлоропластов.
Лейкопласты не содержат пигментов, они бесцветны. Эти самые мелкие пластиды имеют округлую, яйцевидную или палочковидную форму. В клетке они часто группируются вокруг ядра.
Внутренне структура, еще менее дифференцирована по сравнению с хлоропластами. В них осуществляется синтез крахмала, жиров, белков. В соответствии с этим выделяют три вида лейкопластов – амилопласты (крахмал), олеопласты (растительные масла) и протеопласты (белки).
Возникают лейкопласты из пропластид, с которыми они сходны по форме и строению, а отличаются лишь размерами.
Все пластиды генетически связаны друг с другом. Они образуются из пропластид – мельчайших бесцветных цитоплазматических образований, сходных по внешнему виду с митохондриями. Пропластиды находятся в спорах, яйцеклетках, в эмбриональных клетках точек роста. Непосредственно из пропластид образуются хлоропласты (на свету) и лейкопласты (в темноте), а из них развиваются хромопласты, являющиеся конечным продуктом в эволюции пластид в клетке.
Комплекс Гольджи – впервые был обнаружен в 1898 г. Итальянским ученым Гольджи в животных клетках. Это система внутренних полостей, цистерн (5-20), располагающихся сближено и параллельно друг другу, и крупных и мелких вакуолей. Все эти образования имеют мембранное строение и являются специализированными участками эндоплазматической сети. В животных клетках комплекс Гольджи развит лучше, чем в растительных; в последних он называется диктиосомы.
Рис. Строение комплекса Гольджи.
Попадающие в пластинчатый комплекс белки и липиды, подвергаются различным преобразованиям, накапливаются, сортируются, упаковываются в секреторные пузырьки и транспортируются по назначению: к различным структурам внутри клетки или за пределы клетки. Мембраны комплекса Гольджи также синтезируют полисахариды и образуют лизосомы. В клетках молочных желез комплекс Гольджи участвует в образовании молока, а в клетках печени – желчи.
Функции комплекса Гольджи:
1) концентрация, обезвоживание и уплотнение синтезированных в клетке белков, жиров, полисахаридов и веществ, поступивших извне;
2) сборка сложных комплексов органических веществ и подготовка их к выведению из клетки (целлюлоза и гемицеллюлоза у растений, гликопротеины и гликолипиды у животных);
3) синтез полисахаридов;
4) образование первичных лизосом.
Лизосомы — небольшие овальные тельца диаметром 0,2-2,0 мкм. Центральное положение занимает вакуоль, содержащая 40 (по разным данным 30-60) гидролитических ферментов, способных в кислой среде (рН 4,5-5) расщеплять белки, нуклеиновые кислоты, полисахариды, липиды и другие вещества.
Вокруг этой полости располагается строма, одетая снаружи элементарной мембраной. Расщепление веществ при помощи ферментов называется лизисом, поэтому органоид назван лизосомой. Образование лизосом происходит в комплексе Гольджи. Первичные лизосомы приближаются непосредственно к пиноцитозным или фагоцитозным вакуолям (эндосомам) и изливают свое содержимое в их полость, образуя вторичные лизосомы (фагосомы), внутри которых и происходит переваривание веществ. Продукты лизиса через мембрану лизосом поступают в цитоплазму и включаются в дальнейший обмен веществ. Вторичные лизосомы с остатками непереваренных веществ называются остаточными тельцами. Примером вторичных лизосом являются пищеварительные вакуоли простейших.
Функции лизосом:
1) внутриклеточное переваривание макромолекул пищи и чужеродных компонентов, поступающих в клетку при пино- и фагоцитозе, обеспечивая клетку дополнительным сырьем для биохимических и энергетических процессов;
2) при голодании лизосомы переваривают некоторые органоиды и на какое-то время пополняют запас питательных веществ;
3) разрушение временных органов эмбрионов и личинок (хвост и жабры у лягушки) в процессе постэмбрионального развития;
Рис. Образование лизосом
Вакуоли – полости в цитоплазме растительных клеток и протист, заполненные жидкостью. Имеют форму пузырьков, тонких канальцев и другую. Вакуоли образуются из расширений эндоплазматической сети и пузырьков комплекса Гольджи как тончайшие полости, затем по мере роста клетки и накопления продуктов обмена объем их увеличивается, а количество сокращается. Развитая сформировавшаяся клетка имеет обычно одну большую вакуоль, занимающую центральное положение.
Вакуоли растительных клеток заполнены клеточным соком, который представляет собой водный раствор органических (яблочная, щавелевая, лимонная кислоты, сахара, инулин, аминокислоты, белки, дубильные вещества, алкалоиды, глюкозиды) и минеральных (нитраты, хлориды, фосфаты) веществ.
У протист встречаются пищеварительные вакуоли и сократительные.
Функции вакуолей:
1) хранилища запасных питательных веществ и вместилища выделений ( у растений);
2) определяют и поддерживают осмотическое давление в клетках;
3) обеспечивают внутриклеточное пищеварение у протист.
Рис. Клеточный центр.
Клеточный центр обычно находится вблизи ядра и состоит из двух центриолей, расположенных перпендикулярно друг другу и окруженных лучистой сферой. Каждая центриоль представляет собой полое цилиндрическое тельце длиной 0,3-0,5 мкм и длиной 0,15 мкм, стенка которого образована 9 триплетами микротрубочек. Если центриоль лежит в основании реснички или жгутика, то ее называют базальным тельцем.
Перед делением центриоли расходятся к противоположным полюсам и возле каждой из них возникает дочерняя центриоль. От центриолей, расположенных на разных полюсах клетки, образуются микротрубочки, растущие навстречу друг другу. Они формируют митотическое веретено, способствующее равномерному распределению генетического материала между дочерними клетками, являются центром организации цитоскелета. Часть нитей веретена прикрепляется к хромосомам. В клетках высших растений клеточный центр центриолей не имеет.
Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы. Они возникают в результате дупликации уже имеющихся. Это происходит при расхождении центриолей. Незрелая центриоль содержит 9 одиночных микротрубочек; по-видимому, каждая микротрубочка является матрицей при сборке триплетов, характерных для зрелой центриоли.
Центросома характерна для клеток животных, некоторых грибов, водорослей, мхов и папоротников.
Функции клеточного центра:
1) образование полюсов деления и формирование микротрубочек веретена деления.
Рибосомы — мелкие сферические органоиды, от 15 до 35 нм. Состоят из двух субъединиц большой (60S) и малой (40S). Содержат около 60% белка и 40% рибосомальной РНК. Молекулы рРНК образуют ее структурный каркас. Большинство белков специфически связано с определенными участками рРНК. Некоторые белки входят в состав рибосом только во время биосинтеза белка. Субъединицы рибосом образуются в ядрышках. и через поры в ядерной оболочке поступают в цитоплазму, где располагаются либо на мембране ЭПА, либо на наружной стороне ядерной оболочки, либо свободно в цитоплазме. Сначала на ядрышковой ДНК синтезируются рРНК, которые затем покрываются поступающими из цитоплазмы рибосомальными белками, расщепляются до нужных размеров и формируют субъединицы рибосом. Полностью сформированных рибосом в ядре нет. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка. По сравнению с митохондриями, пластидами, клетками прокариот рибосомы в цитоплазме эукариотических клеток крупнее. Могут объединяться по 5-70 единиц в полисомы.
Функции рибосом:
1) участие в биосинтезе белка.
Рис. 287. Рибосома: 1 — малая субъединица; 2 — большая субъединица.
Реснички, жгутики – выросты цитоплазмы, покрытые элементарной мембраной, под которой находится 20 микротрубочек, образующих 9 пар по периферии и две одиночные в центре. У основания ресничек и жгутиков расположены базальные тельца. Длина жгутиков достигает 100 мкм. Реснички – это короткие – 10-20 мкм – жгутики. Движение жгутиков винтовое, а ресничек – веслообразное. Благодаря ресничкам и жгутикам передвигаются бактерии, протисты, ресничные, перемещаются частицы или жидкости (реснички мерцательного эпителия дыхательных путей, яйцеводов), половые клетки (сперматозоиды).
Рис. Строение жгутиков и ресничек эукариот
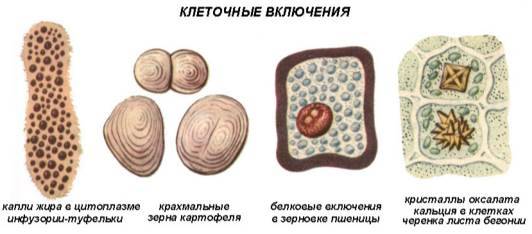
Включения — временные компоненты цитоплазмы, то возникающие, то исчезающие. Как правило, они содержатся в клетках на определенных этапах жизненного цикла. Специфика включений зависит от специфики соответствующих клеток тканей и органов. Включения встречаются преимущественно в растительных клетках. Они могут возникать в гиалоплазме, различных органеллах, реже в клеточной стенке.
В функциональном отношении включения представляют собой либо временно выведенные из обмена веществ клетки соединения (запасные вещества — крахмальные зерна, липидные капли и отложения белков), либо конечные продукты обмена (кристаллы некоторых веществ).
Крахмальные зерна. Это наиболее распространенные включения растительных клеток. Крахмал запасается у растений исключительно в виде крахмальных зерен. Они образуются только в строме пластид живых клеток. В процессе фотосинтеза в зеленых листьях образуется ассимиляционный, или первичный крахмал. Ассимиляционный крахмал в листьях не накапливается и, быстро гидролизуясь до сахаров, оттекает в части растения, в которых происходит его накопление. Там он вновь превращается в крахмал, который называют вторичным. Вторичный крахмал образуется и непосредственно в клубнях, корневищах, семенах, то есть там, где он откладывается в запас. Тогда его называют запасным. Лейкопласты, накапливающие крахмал, называют амилопластами. Особенно богаты крахмалом семена, подземные побеги (клубни, луковицы, корневища), паренхима проводящих тканей корней и стеблей древесных растений.
Липидные капли. Встречаются практически во всех растительных клетках. Наиболее богаты ими семена и плоды. Жирные масла в виде липидных капель — вторая по значению (после крахмала) форма запасных питательных веществ. Семена некоторых растений (подсолнечник, хлопчатник и т.д.) могут накапливать до 40% масла от массы сухого вещества.
Липидные капли, как правило, накапливаются непосредственно в гиалоплазме. Они представляют собой сферические тела обычно субмикроскопического размера. Липидные капли могут накапливаться и в лейкопластах, которые называют элайопластами.
Белковые включения образуются в различных органеллах клетки в виде аморфных или кристаллических отложений разнообразной формы и строения. Наиболее часто кристаллы можно встретить в ядре — в нуклеоплазме, иногда в перинуклеарном пространстве, реже в гиалоплазме, строме пластид, в расширениях цистерн ЭПР, матриксе пероксисом и митохондриях. В вакуолях встречаются как кристаллические, так и аморфные белковые включения. В наибольшем количестве кристаллы белка встречаются в запасающих клетках сухих семян в виде так называемых алейроновых3 зерен или белковых телец.
Запасные белки синтезируются рибосомами во время развития семени и откладываются в вакуоли. При созревании семян, сопровождающемся их обезвоживанием, белковые вакуоли высыхают, и белок кристаллизуется. В результате этого в зрелом сухом семени белковые вакуоли превращаются в белковые тельца (алейроновые зерна).
Включения классифицируют на трофические, представляющие собой запасы питательных веществ (углеводные – гликоген в клетках печени и мышцах, белковые – желточные включения в яйцеклетках, жировые – капли жира в клетках подкожной жировой клетчатки), секреторные продукты жизнедеятельности клеток желез внешней и внутренней секреции (ферменты, гормоны, слизь и др.), пигментные – содержат различные пигменты (меланин, липофусцин, гемоглобин и др.) и экскреторные – продукты обмена веществ, подлежащие удалению из клетки, (кристаллы щавелевой кислоты, щавелевокислого кальция, мочевина)
Источник: helpiks.org
Органоиды специального назначения содержатся во многих растительных и животных клетках. К ним относятся органоиды движения (миофибриллы, реснички, жгутики, стрекательные капсулы и др.), опорные структуры (тонофибриллы), органоиды, воспринимающие внешние раздражения (например, фоторецепторы, статорецепторы и фонорецепторы), нейрофибриллы, а также структуры клеточной поверхности, связанные с всасыванием и перевариванием пищи (микроворсинки, кутикула и др. виды.)
Реснички и жгутики — это выступающие из клетки органеллы, имеющие диаметр около 0,25 мкм и содержащие в середине пучок параллельно расположенных микротрубочек . Главная функция этих органелл состоит в передвижении самих клеток или в продвижении вдоль клеток окружающей их жидкости и частиц. Реснички и жгутики имеются на поверхности клеток многих типов и встречаются у большинства животных и некоторых растений. У человека множество ресничек имеют клетки эпителия бронхов (до 10#9 на 1 см2). Они заставляют постоянно двигаться вверх слой слизи с частицами пыли и остатками отмерших клеток. С помощью ресничек клеток яйцевода яйцеклетки продвигаются по нему. Жгутики отличаются от ресничек лишь длиной. Так, сперматозоиды млекопитающих имеют по одному жгутику длиной до 100 мкм. Обычно реснички короче жгутиков более чем в 10 раз. Тысячи ресничек одной клетки движутся координированно, образуя на поверхности плазмалеммы бегущие волны Каждая ресничка работает подобно хлысту: удар вперед, при котором ресничка полностью выпрямляется и передает в окружающую жидкость максимальное усилие, проталкивая ее, а затем, изгибаясь, чтобы уменьшить сопротивление среды, она возвращается в исходное положение). На всю длину реснички или жгутика тянутся микротрубочки — полые белковые цилиндры с внешним диаметром 25 нм. Микротрубочки, как и микрофиламенты, полярны, они удлиняются с одного конца вследствие полимеризации глобулярного белка. В ресничках и жгутиках они располагаются по системе 9+2; девять двойных микротрубочек (дублетов) образуют стенку цилиндра, в центре которого находятся две одиночные микротрубочки ю.Дублеты способны скользить друг относительно друга, что заставляет ресничку или жгутик изгибаться.
Микротрубочки
Микротрубочки — белковые внутриклеточные структуры, входящие в состав цитоскелета.Микротрубочки представляют собой полые внутри цилиндры диаметром 25 нм. Длина их может быть от нескольких микрометров до, вероятно, нескольких миллиметров в аксонах нервных клеток. Их стенка образована димерами тубулина. Микротрубочки, подобно актиновым микрофиламентам, полярны: на одном конце происходит самосборка микротрубочки, на другом — разборка. В клетках микротрубочки играют роль структурных компонентов и участвуют во многих клеточных процессах, включая митоз, цитокинез и везикулярный транспорт.Содержание [показать]
Строение Микротрубочки — это структуры, в которых 13 тубулиновых α-/β-гетеродимеров уложены по окружности полого цилиндра. Внешний диаметр цилиндра около 25 нм, внутренний — около 15.Один из концов микротрубочки, называемый плюс-концом, постоянно присоединяет к себе свободный тубулин. От противоположного конца — минус-конца — тубулиновые единицы отщепляются.
ФункцияМикротрубочки в клетке используются в качестве «рельсов» для транспортировки частиц. По их поверхности могут перемещаться мембранные пузырьки и митохондрии. Транспортировку по микротрубочкам осуществляют белки, называемые моторными. Это высокомолекулярные соединения, состоящие из двух тяжёлых (массой около 300 кДа) и нескольких лёгких цепей. В тяжёлых цепях выделяют головной и хвостовой домены. Два головных домена связываются с микротрубочками и являются собственно двигателями, а хвостовые — связываются с органеллами и другими внутриклеточными образованиями, подлежащими транспортировке.
Помимо транспортной функции, микротрубочки формируют центральную структуру ресничек и жгутиков — аксонему. Типичная аксонема содержит 9 пар объединённых микротрубочек по периферии и две полных микротрубочки в центре. Из микротрубочек состоят также центриоли и веретено деления, обеспечивающее расхождение хромосом к полюсам клетки при митозе и мейозе. Микротрубочки участвуют в поддержании формы клетки и расположения органоидов (в частности, аппарата Гольджи) в цитоплазме клеток.
ОРГАНОИДЫ СПЕЦИАЛЬНОГО НАЗНАЧЕНИЯ
| Микротрубочки – длинные тонкие полые цилиндры диаметром 25 нм . стенки микротрубочек состоят из белков | 1.опорная функция образуют внутренний каркас помогающий клеткам сохранять форму 2.двигательная-входят в состав ресничек и жгутиков |
| Мвыросикронити – тонкие структуры состоящие из тысяч молекул белков соединенных друг с другом | Образуют опорно-двигательную систему называемую цитоскелетом . способствует току цитоплазмы в клетках |
| Реснички –многочисленные цитоплазматические выросты на поверхности мембраны образованы микротрубочками покрытыми мембраной | Обеспечивают передвижение некоторых одноклеточных организмов и ток жидкости в организмах удаление частичек пыли |
| Жгутики –— поверхностная структура, присутствующая у многих прокариотических и эукариотических клеток и служащая для их движения в жидкой среде или по поверхности твёрдых сред. Жгутики прокариот и эукариот резко различаются: бактериальный жгутик имеет толщину 10—20 нм и длину 3—15 мкм, он пассивно вращается расположенным в мембране мотором; жгутики же эукариот толщиной до 200 нм и длиной до 200 мкм, они могут самостоятельно изгибаться по всей длине. У эукариот часто также присутствуют реснички, идентичные по своему строению жгутику, но более короткие (до 10 мкм). | Служат для движения одноклеточным организмам сперматозоидам и зооспорам |
Вопрос 17.
Включения – необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клетки.
Это скопление веществ в клетке.
Классификация:
Трофические (нейтральные липиды, полисахариды, белки)
Секреторные (вакуоли, выводящие вещества из клетки)
Экскреторные (продукты метаболизма)
Пигментные – экзогенные (каротин, пыль, красители)
— эндогенные (гемоглобин, меланин)
Источник: megaobuchalka.ru
Лекция №4
СПЕЦИАЛЬНЫЕ ОРГАНОИДЫ И ВКЛЮЧЕНИЯ
Органоиды специального назначения
Органоиды специального назначения содержатся во многих животных и растительных клетках. От общих органоидов они отличаются тем, что характерны только для определенных высоко дифференцированных клеток и выполняют строго определенную функцию, характерную для этих клеток.
Классификация органоидов специального назначения:
1. Органоиды движения: реснички, жгутики, миофибриллы.
2. Опорные структуры: тонофибриллы.
3. Органоиды, участвующие в передаче возбуждения: нейрофибриллы.
4. Органоиды, воспринимающие внешние раздражения: фоторецепторы, статорецепторы, фонорецепторы.
5. Органоиды поверхности клеток: микроворсинки, кутикула.
6. Органоиды защиты и нападения у одноклеточных: трихоцисты – у инфузорий; коноид, роптрии – у представителей класса Споровиков.
Рассмотрим более подробно основные из этих органоидов.
Реснички и жгутики
Реснички и жгутики – это нитевидные или волосковидные выросты свободной поверхности клеток. С помощью ресничек и жгутиков клетки могут передвигаться в жидкой среде, так как эти органоиды способны совершать ритмические движения. Если же реснички и жгутики имеются у прикрепленных к какому-либо субстрату клеток, то они вызывают движение окружающей жидкости.
Различий в тонкой организации этих структур нет. Если на поверхности клетки имеется большое количество волосковидных выростов небольшой длины, то их называют ресничками, если же таких выростов мало и длина их значительная, то они называются жгутиками.
У животных реснички и жгутики встречаются: а) в клетках ресничного эпителия (эпителий трахеи, некоторых отделов полового тракта); б) у сперматозоидов (у нематод и десятиногих раков спермии не имеют жгута); в) у простейших (жгутиконосцы, инфузории, корненожки). В мире растений они имеются у подвижных зооспор водорослей, мхов, папоротников, низших грибов, миксомицетов. Клетки высших растений и высших грибов, а также споровики не имеют ресничек и жгутиков даже у мужских половых клеток.
Толщина ресничек и жгутиков составляет около 200 нм (0,2 мкм). Поскольку принципиальных различий в строении ресничек и жгутиков нет, рассмотрим ультраструктуру этих образований на примере реснички. Снаружи ресничка покрыта цитоплазматической мембраной. Внутри нее расположена аксонема (или осевой цилиндр), состоящая из микротрубочек. Нижняя проксимальная часть реснички, базальное тельце, погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы.
Базальное тельце по своей структуре совершенно сходно с центриолью и состоит из 9 триплетов микротрубочек. Аксонема в своем составе, в отличие от базального тельца, имеет 9 пар (дублетов) микротрубочек, образующих внешнюю стенку цилиндра аксонемы. Дублеты микротрубочек слегка повернуты (около 100) по отношению к радиусу аксонемы. Кроме периферических дублетов микротрубочек в центре аксонемы располагается пара центральных микротрубочек. Эти две центральные микротрубочки, в отличие от периферических, не доходят до базальных телец. Поскольку в базальных тельцах содержится сократимый белок типа актомиозина, периферические микротрубочки выполняют двигательную функцию, а центральные – только опорную.
В основании ресничек и жгутиков часто встречаются корешки или кинетодесмы, представляющие собой пучки тонких (6 нм) фибрилл, обладающих поперечных исчерченностью. Часто такие исчерченные кинетодесмы простираются от базальных телец в глубь цитоплазмы в направлении к ядру. Роль этих структур еще недостаточно выяснена.
Отклонения от вышеизложенного плана строения встречаются редко, но у некоторых клеток, например, в жгутиках сперматозоидов и некоторых жгутиконосцев, обнаружены 9 дополнительных фибрилл, расположенных между центральными и периферическими микротрубочками. Эти дополнительные фибриллы соединены с трубочками аксонемы с помощью очень тонких волокон.
Миофибриллы
Миофибриллы представляют собой особые дифференцированные сократимые элементы клетки, за счет которых происходят сложные и совершенные движения мышц. Различают два типа миофибрилл: гладкие и поперечнополосатые. Оба типа миофибрилл широко распространены у многоклеточных животных и у простейших.
Поперечнополосатые миофибриллы широко известны в составе соматической и сердечной мускулатуры членистоногих и хордовых животных. Гладкие миофибриллы типичны для мускулатуры внутренних органов позвоночных и для соматических мышц многих низших беспозвоночных.
Строение миофибрилл наиболее подробно изучено в поперечно-полосатых мышечных волокнах. Миофибрилла имеет толщину 0,5 мкм и длину, которая равна от 10-20 мкм до нескольких миллиметров и даже сантиметров. В световой микроскоп видно, что пучки миофибрилл окрашиваются неравномерно: через равные промежутки длины в них видно чередование темных и светлых участков. Темные участки имеют двойное лучепреломление и называются анизотропными дисками (А-диски). Светлые участки двойного лучепреломления не обнаруживают и называются изотропными дисками (I-диски).
Каждый А-диск разделяется на две половины менее плотной, чем остальные его участки, полосой, называемой Н-зоной (полоска Ханзена). Посередине каждого I-диска проходит темная линия, называемая Z-линией (телофрагма). Участок миофибриллы между двумя Z-линиями называется саркомером. Он является единицей строения и функционирования миофибриллы.
Подробности строения саркомера были получены только при изучении миофибрилл в электронном микроскопе. Каждая миофибрилла состоит из пучка очень тонких нитей – миофиламентов. Различают два типа миофиламентов: толстые и тонкие. Тонкие миофиламенты имеют диаметр около 7 нм и длину около 1 мкм; они состоят в основном из белка актина. Они располагаются в пределах I-диска и заходят в А-диск до Н-зоны. Толстые миофиламенты длиной до 1,5 мкм и толщиной около 15 нм состоят из белка миозина; они расположены только в пределах А-диска. В тонких миофиламентах кроме актина находятся также белки тропомиозин и тропонин. Z-линии имеют в своем составе белок α-актинин и десмин.
Ни актин, ни миозин по отдельности не обладают сократительной способностью. Актин, белок с молекулярным весом 43,5 тысяч, является глобулярным белком размером около 3 нм. В присутствии АТФ и некоторых белковых факторов он способен к агрегации в виде нитчатых структур толщиной до 7 нм. Такие актиновые фибриллы состоят из двух спиралей, обвивающих друг друга. Миозин, входящий в состав толстых нитей, – очень крупный белок (мол. вес 470 тысяч), состоящий из шести цепей: двух длинных, спирально обвивающихся одна вокруг другой, и четырех коротких, которые связываются с концами длинных цепей и образуют глобулярные «головки». Последние обладают АТФ-азной активностью, могут реагировать с фибриллярным актином, образуя актомиозиновый комплекс, способный к сокращению.
Актиновые миофиламенты связаны на одном конце с Z-линией, которая состоит из ветвящихся молекул белка α-актинина, образующих фибриллярную сеть, идущую поперек миофибриллы. С двух сторон к Z-линии прикрепляются концы актиновых нитей соседних саркомеров. Функция Z-линий заключается как бы в связывании соседних саркомеров друг с другом; Z-линии не являются сократимыми структурами.
Механизм мышечного сокращения заключается в одновременном укорачивании всех саркомеров по всей длине миофибриллы. Г. Хаксли показал, что в основе сокращения лежит перемещение относительно друг друга толстых и тонких нитей. При этом толстые миозиновые нити как бы входят в пространство между актиновыми нитями, приближая друг к другу Z-линии. Эта модель скользящих нитей может объяснить не только сокращение поперечнополосатых мышц, но и любых сократимых структур.
В гладких мышечных клетках также имеются актиновые и миозиновые нити, но они не так правильно расположены, как в исчерченных мышцах. Здесь нет саркомеров, а просто среди пучков актиновых миофиламентов без особого порядка располагаются миозиновые молекулы.
Тонофибриллы
Тонофибриллы характерны для клеток одноклеточных организмов и для эпителиальных клеток многоклеточных животных. Электронно-микроскопическое исследование показало, что они состоят из пучка тонофиламентов – тончайших нитей с диаметром 6-15 нм. В одном пучке может быть от 3 до нескольких сотен тонофиламентов.
Тонофибриллы располагаются пучками в клетке в разных направлениях, прикрепляются либо к десмосомам, либо к любому участку цитоплазматической мембраны и никогда не переходят из одной клетки в другую.
Тонофибриллы выполняют в клетке опорную функцию.
Нейрофибриллы
Нейрофибриллы открыты в 1855 г. Ф.В. Овсянниковым. Они характерны для нервных клеток (нейронов). Состоят из более тонких нитей – нейрофиламентов.
В теле нейрона нейрофибриллы расположены беспорядочно, а в отростках образуют пучок параллельно длине отростка. Из этого правила имеется всего лишь два исключения: параллельное, упорядоченное расположение нейрофибрилл в теле нейрона впервые обнаружено у бешеных животных, а затем у животных, которые впадают в спячку.
Открытие нейрофибрилл привело к возникновению нейрофибриллярной теории проведения нервного возбуждения. Сторонники этой теории считали, что нейрофибриллы являются беспрерывным проводящим элементом нервной системы. Однако в дальнейшем было установлено, что нейрофибриллы не переходят из одного нейрона в другой. В настоящее время мы придерживаемся нейронной теории, согласно которой в проведении нервного импульса основная роль принадлежит плазмалемме нейрона, а по нейрофибриллам из тела нейрона к его окончанию передаются вещества, участвующие в образовании нервных импульсов. А с одной клетки на другую возбуждение передается с помощью синапса (строение синапса описывалось ранее при рассмотрении коммуникационных межклеточных контактов). В синапсе возбуждение передается химическим путем с помощью медиатора.
Непостоянные включения в клетке
В отличие от органоидов, как общего, так и специального назначения, включения представляют собой непостоянные образования, то возникающие, то исчезающие в процессе жизнедеятельности клетки. Основное место локализации включений – это цитоплазма, но они иногда встречаются и в ядре.
По своему характеру все включения – это продукты клеточного метаболизма. По химическому составу и по выполняемым функциям они классифицируются следующим образом:
1. трофические (белковые, углеводные, жировые);
2. секреторные;
3. экскреторные;
4. пигментные.
Трофические включения
Белковые включения. Имеют форму зерен, гранул, дисков. Они могут присутствовать во всех клетках, но встречаются реже, чем жиры и углеводы. Примером белковых включений служит желток в яйцеклетках, алейроновые зерна в эндосперме семян. В этом случае белковые гранулы служат запасным питательным материалом для зародыша; в других клетках – это трофический (строительный) материал для дальнейшего построения элементов клетки. Энергетическим запасом белковые включения могут служить в самом крайнем случае, когда углеводные и жировые запасы полностью израсходовались.
Углеводные включения также запасаются в твердом виде (глыбки, зерна разнообразных размеров и форм). У многоклеточных животных и простейших в цитоплазме клеток встречаются отложения гликогена. Обычно большие скопления гликогена сосредоточены в цитоплазме поперечнополосатых мышечных волокон, клетках печени, нейронах, у эндопаразитических организмов – гельминтов и простейших. Гранулы гликогена могут располагаться не только в цитоплазме, но и в ядрах многих клеток, в частности, в ядрах клеток печени человека (более мелкие гранулы, чем в цитоплазме).
В клетках растений наиболее часто откладывается крахмал в виде зерен различной формы и размеров, причем форма крахмальных зерен специфична для каждого вида растений и для определенных тканей. Отложениями крахмала богата цитоплазма клубней картофеля, зерен злаков, бобовых растений и др. У низших растений встречаются другие полисахариды: парамилоид, крахмал красных водорослей.
Углеводные включения являются основным энергетическим запасом клетки. При распаде 1 г углевода выделяется 17,6 кДж энергии, которая накапливается в виде АТФ.
Жировые включения. Жиры в цитоплазме откладываются в виде мелких капель. Они встречаются как у животных, так и у растений. В одних клетках жировых включений очень мало и они постоянно используются самой клеткой в процессе обмена веществ, в других клетках они накапливаются в большом количестве, например, жировые клетки соединительной ткани, клетки эпителия печени рыб и амфибий. Большое количество жировых капель встречается и в цитоплазме многих видов простейших, например, инфузорий. Очень много жира содержится в семенах растений, причем количество его может доходить до 70% сухого веса семян (масличные культуры).
Процесс отложения жиров не связан с какими-либо органоидами клетки; они откладываются в основном веществе цитоплазмы. При определенных условиях жировые капли могут сливаться друг с другом, увеличиваясь в размерах, в конечном итоге гигантская жировая капля заполняет собой всю клетку, цитоплазма с ядром отмирают и клетка превращается в мешочек с жиром. Это явление называется жировое перерождение клетки. Этот процесс может носить патологический характер (например, при жировом перерождении печени, сердечной мышцы и т.д.) или являться естественным процессом в жизнедеятельности организма (например, клетки сальных желез, клетки подкожной жировой клетчатки китов, тюленей).
Жировые включения могут выполнять следующие функции:
1) являются долговременным энергетическим запасом клетки (при распаде 1 г жира выделяется 38,9 кДж энергии);
2) терморегуляция (например, у животных, обитающих в холодном климате слой жира в подкожной клетчатке достигает 1 м);
3) амортизация при движении (например, прослойки жира на подошвах ног, на лапах у наземных животных, ладонях рук, вокруг внутренних органов);
4) запас питательных веществ у животных, впадающих в спячку (например, медведь, барсук, еж);
5) источник метаболической воды в организме у животных, обитающих в засушливых условиях (при распаде 1 кг жира образуется 1,1 кг воды).
Секреторные включения
Секреты – это продукты анаболических реакций клетки, которые выполняют в организме различные жизненно важные функции.
Секреторные включения накапливаются в секреторных клетках в виде зерен, гранул, капель. Химическая природа их весьма разнообразна. Это могут быть белки, липиды, кетоны, спирты, соляная кислота и другие. В клетках многих растений встречаются и кристаллические включения, причем чаще всего это оксалаты кальция.
Функции секреторных включений:
1) гуморальная регуляция жизнедеятельности организма (гормоны в клетках желез внутренней секреции);
2) катализация процессов переваривания пищи (ферменты в клетках желез пищеварительного тракта);
3) передача возбуждения в синапсах (медиаторы в пресинаптических окончаниях нейронов);
4) питательные вещества для детенышей (молоко в млечных железах млекопитающих);
5) защитная функция (слизь у земноводных защищает кожу от пересыхания; яды, токсины у животных защищают от врагов и помогают умерщвлять добычу).
Из клеток секреты удаляются различными способами. По способу удаления секрета из клетки выделяют 3 типа секреции:
1) мерокриновая – секрет удаляется через поры без повреждения клетки; такая клетка функционирует непрерывно (например, железы дна желудка);
2) апокриновая – капли секрета отшнуровываются с частью цитоплазмы; такая клетка функционирует с перерывами, необходимыми для ее восстановления (например, слюнные железы, часть потовых)
3) голокриновая – секрет заполняет клетку целиком, цитоплазма отмирает, клетка гибнет и превращается в мешочек с секретом; такая клетка функционирует всего один раз (например, сальные железы).
Экскреторные включения
Экскреторные включения – это продукты катаболических реакций, которые клеткой и организмом не используются, часто являются ядовитыми и должны удаляться. Экскреты могут накапливаться в жидком (капли) и в твердом (зерна, гранулы) состоянии.
Примерами экскреторных включений могут служить капли пота в клетках потовых желез, моча в клетках почечных канальцев. У многих беспозвоночных животных существуют специальные клетки – нефроциты, которые функционируют как почки накопления. Они накапливают экскреты, а затем либо выносят их в кишечник или на поверхность тела, либо оставляют в составе своей цитоплазмы. Важную роль в обособлении ядовитых экскретов играет комплекс Гольджи. Примерами нефроцитов являются хлорагогенные клетки у кольчатых червей, перикардиальные клетки у моллюсков и насекомых, экскретофоры у ресничных червей и асцидий.
Пигментные включения
Пигментные включения могут существовать в виде гранул, зерен, изредка в виде капель. Основная их функция – придание окраски растительным и животным клеткам и организму в целом. Но в ряде случаев пигментные включения выполняют более сложные функции. Рассмотрим в качестве примера некоторые пигменты животного и растительного мира.
Пигменты животного мира:
1). Меланин – пигмент коричневого цвета, расположен в клетках базального слоя кожи, придает окраску эпителию кожи и всем ее производным (волосы у человека, шерсть у животных, ногти, когти, перья у птиц, чешуи у рептилий), а также радужной оболочке глаза. У животных меланин создает различные виды защитной окраски, а у человека выполняет функцию защиты от ультрафиолетового излучения.
2). Липофусцин – пигмент желтого цвета, гранулы которого накапливаются в процессе жизнедеятельности клеток и, особенно, по мере старения их, а также при разных дистрофических процессах («пигмент старения»).
3). Лютеин – желтый пигмент, содержащийся в желтом теле беременности.
4). Ретинин – характерный пигмент, входящий в состав зрительного пурпура сетчатки глаза.
5). Дыхательные пигменты животных:
– гемоцианин – пигмент, содержащий в своем составе медь; он может изменять свою окраску от синей (в окисленном состоянии) до бесцветной (в восстановленном состоянии); встречается у ракообразных, некоторых улиток, головоногих моллюсков (растворен в плазме крови или гемолимфе);
– гемоэритрин – пигмент, содержащий в своем составе железо; он может изменять свою окраску от красной (в окисленном состоянии) до бесцветной (в восстановленном состоянии); встречается у некоторых кольчатых червей (находится в клетках крови);
– хлорокруорин – пигмент, также содержащий в своем составе железо; он может изменять свою окраску от красной (в окисленном состоянии) до зеленой (в восстановленном состоянии); встречается у некоторых многощетинковых червей (растворен в плазме крови);
– гемоглобин – железосодержащий пигмент, меняет свою окраску от оранжево-красной (в окисленном состоянии) до пурпурно-красной (в восстановленном состоянии). Это наиболее широко распространенный в природе дыхательный пигмент, встречается у некоторых моллюсков (растворен в плазме крови), у некоторых кольчатых червей (в плазме или в клетках), у всех позвоночных животных (в эритроцитах крови).
Пигменты растительного мира:
1). Хлорофилл – пигмент зеленого цвета, находится в гранах хлоропластов и участвует в процессе фотосинтеза.
2). Группа каротиноидов – каротин (оранжевый), ксантофилл (красный), ликопин (желтый); эти пигменты содержатся в хромопластах и обеспечивают окраску плодов, семян и других органов растений.
5). Фикобилины – это пигменты низших растений; в состав сине-зеленых водорослей входит фикоциан (пигмент синего цвета), а в состав красных водорослей – фикоэритрин (красный пигмент).
Изменение окраски клеток обусловлено перераспределением пигментов.
Источник: refleader.ru
К органоидам специального назначения относятся микроворсинки, реснички, жгутики, миофибриллы, псевдоподии.
Реснички и жгутики –это подвижные цитоплазматические выросты, в которых находится осевая нить (или аксонема), а у основания – базальное тельце. Базальное тельце служит матрицей для организации аксонемы. Само базальное тельце состоит из 9 триплетов микротрубочек, располагается в основании реснички или жгутика и напоминает по строению центриоль.
Аксонема состоит из 9 периферических и двух центрально расположенных пар микротрубочек. Центральная пара окружена центральной оболочкой, от которой к периферическим парам в виде лучей расходятся радиальные спицы. Периферические пары микротрубочек связаны между собой белком нексином. Кроме того, от микротрубочек соседних пар отходят ручки из белка динеина, обладающего активностью АТФ-азы. Аксонема формируется путем самосборки белковых субъединиц. Матрицей для самосборки служит базальное тельце.
Микроворсинки – это цитоплазматические выросты длиной около 1 мкм. Они многократно увеличивают поверхность клетки. Каждая микроворсинка имеет внутренний каркас, образованный пучком около 40 микрофиламентов, расположенных вдоль клеточной оси и фиксирующихся на внутренней поверхности плазмолеммы.
Миофибриллы– органоиды специального назначения мышечных волокон. Они имеют вид тонких нитей, вытянутых вдоль мышечных волокон и отделяющихся друг от друга рядами вытянутых митохондрий и эндоплазматической сети. Сами миофибриллы состоят из сократительных белков актина, миозина и других.
Псевдоподии (ложноножки) –это временные цитоплазматические выпячивания у некоторых одноклеточных организмов и некоторых клеток (например, лейкоцитов) многоклеточных животных. Служат для амебоидного движения и фагоцитоза.
Цитоскелет
Цитоскелет – это трехмерный цитоплазматический комплекс волокнистых и трубчатых структур, придающий клетке определенную форму и выполняющий другие функции. Он представлен микротрубочками, микрофиламентами и промежуточными филаментами.
Микротрубочки – неветвящийся структурный компонент цитоплазмы, присутствующий практически во всех типах эукариотических клеток. Имеют диаметр 24 нм, а в длину –несколько мкм. Это динамические структуры, постоянно растущие с одного конца и деполяризующиеся с другого. Они построены из белка тубулина. Имеют вид полых цилиндров.
Их главная функция заключается в создании эластичного и устойчивого внутриклеточного каркаса, необходимого для поддержания формы клеток. Кроме того, они принимают участие в транспорте макромолекул и органоидов, обеспечивают подвижность жгутиков и ресничек. Входя в состав веретена деления, обеспечивают расхождение хромосом при делении клеток. Они могут быть факторами ориентированного движения клетки в целом.
Разрушение микротрубочек колхицином приводит к нарушениям транспорта веществ (например, к блокаде секреции) и механического переноса отдельных внутриклеточных компонентов. Кроме того, при делении клеток блокируется веретено деления.
Микрофиламенты – относятся к фибриллярным компонентам эукариотических клеток. Они располагаются в кортикальном слое цитоплазмы, прямо под плазмолеммой, в виде пучков или слоями. В среднем их длина составляет 6 нм.
В зависимости от химического состава микрофиламенты могут выполнять функции цитоскелета и участвовать в обеспечении движения. В их состав входят сократительные белки: актин, миозин, тропомиозин и др.
Основные функции микрофиламентов – это создание внутриклеточного сократительного аппарата, который обеспечивает амебоидное движение клеток, большинство движений органоидов и деление клеток.
Промежуточные филаменты(микрофибриллы) – это тонкие неветвящиеся, нередко лежащие пучками нити, которые присутствуют в клетках разных тканей и состоят из разных веществ, например, из кератина – в эпителии, десмина – в мышечных волокнах и др. Они выполняют опорно-каркасную функцию.
Включения
Включения – это непостоянные компоненты, которые могут появляться и исчезать в зависимости от функционального состояния клетки. Их классифицируют на: трофические, секреторные, экскреторные, пигментные, кристаллы. Трофические включения – это, как правило, резервные скопления гранул белка, гликогена, капельки жиров. Секреторные гранулы и пузырьки содержат биологически активные вещества и поступают по протокам желез в те или иные органы. Экскреторные включения не являются биологически активными веществами. Обычно это продукты метаболизма клеток, подлежащие удалению. Пигментные включения могут изменить цвет ткани временно или постоянно (примеры: гемоглобин, меланин, липофусцин – пигмент изнашивания, билирубин и др.). В растительных клетках встречаются включения кристаллов (чаще всего это оксалат кальция).
Источник: poznayka.org