Основными компонентами хроматина ядра эукариот являются
1.Ядерный аппарат эукариотических клеток имеет ряд отличий от прокариотических. Во-первых, ДНК-содержащий компонент отделен от цитоплазмы специальной оболочкой (ядерная оболочка) , во-вторых, количество ДНК в ядах эукариот в тысячи раз больше, чем в составе нуклеоидов бактерий, в-третьих, ДНК эукариот представляет собой сложный нуклеопротеидный комплекс, образующий специальную структуру — хроматин, из которого и состоят эукариотические хромосомы. Далее — в состав ядер эукариот входят несколько физически не связанных хромосом, каждая из которых содержит одну линейную гигантскую молекулу ДНК. Каждая хромосомная ДНК представляет собой полирепликонную структуру, т. е. содержит множеств автономно реплицирующихся участков. Синтез и образование транскриптов эукариотических клеток сопровождается процессами вторичной их перестройки, «созревания» , включающих в себя как фрагментацию (процессинг) , так и сращивание отдельных фрагментов ДНК (сплайсинг) . Наконец, в ядрах не происходит синтеза белков, т.
в эукариотических клетках процессы синтеза ДНК и РНК разобщены от процесса синтеза белков.
Клеточное ядро, обычно одно на клетку (есть примеры многоядерных клеток) , состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка и других продуктов синтетической активности, ядерного белкового остова (матрикса) и кариоплазмы (или ядерного сока)) . Эти основные компоненты встречаются практически во всех неделящихся клетках эукариотических одно- или многоклеточных организмов.
Главный компонент ядер, хроматин, является структурой, выполняющей генетическую функцию клетки, в хроматиновой ДНК заложена практически вся генетическая информация. Ядерная оболочка выполняет сложную барьерно-рецепторную, а также транспортную и каркасную функцию. Нехроматиновый ядерный белковый остов (матрикс) обеспечивает не только пространственное расположение хромосом в ядре, но и участвует в их функциональной активности. Одним из хромосомных участков, определяющих синтез рРНК и образование клеточных рибосом, является ядрышко. Кроме того в ядре в связи с хроматином и матриксом обнаруживаются различные рибонуклеопротеидные структуры, содержащие разные типы РНК. Между всеми этими компонентами заключена жидкая фаза клеточного ядра, кариоплазма, в которой протекают многие процессы, связанные как с ядерным метаболизмом, так и с внутриядерным транспортом белков и РНК.
2. Наследственность – это свойство организмов сохранять материальную и функциональную преемственность между их поколениями. Благодаря этому свойству обеспечивается непрерывность живого при смене его поколений. Современной биологией установлено, что наследственность связана с определёнными материальными структурами клетки – молекулами ДНК и их редупликацией, происходящей при делении клеток.
Источник: otvet.mail.ru
Вопрос 1. Опишите строение ядра эукариотической клетки.
Ядро окружено оболочкой, которая состоит из двух мембран. Ядерная мембрана со стороны, обращённой в цитоплазму, покрыта рибосомами, внутренняя мембрана ядра гладкая. Ядерная оболочка – часть мембранной системы клетки. Выросты внешней ядерной мембраны соединяются с каналами эндоплазматической сети, образуя единую систему сообщающихся каналов. Между ядром и цитоплазмой осуществляется постоянный обмен веществами.
Несмотря на активный обмен между ядром и цитоплазмой, ядерная оболочка отграничивает ядерное содержимое от цитоплазмы, обеспечивая тем самым различия в их химическом составе. Это необходимо для нормального функционирования ядерных структур.
В гелеобразном ядерном соке располагаются хроматин и одно или несколько ядрышек.
В живой клетке ядерный сок выглядит бесструктурной массой, заполняющей промежутки между структурами ядра. В состав ядерного сока входят различные белки (в том числе большинство ферментов ядра), свободные нуклеотиды, аминокислоты, а также рибонуклеиновые кислоты (РНК), транспортируемые затем из ядра в цитоплазму.
Вопрос 2. Что такое ядрышко? Как вы считаете, можно ли ядрышко выделить из ядра как самостоятельную единую структуру? Объясните свою точку зрения.
Ядрышко – структура, составленная из расположенных рядом участков нескольких различных хромосом. Эти участки представляют собой большие петли ДНК, содержащие гены рибосомальной РНК (рРНК). Такие петли называются ядрышковым организатором.
Ядрышко — не самостоятельная структура или органоид. Оно — производное
хромосомы, один из ее локусов, активно функционирующий в интерфазе.
Ядрышко является центром образования рибосом, т.к. здесь осуществляется синтез рРНК и соединение этих молекул с белками, т.е. происходит формирование субъединиц рибосом, которые затем поступают в цитоплазму, где и завершается сборка рибосом.
Вопрос 3. Что такое хроматин? Опишите строение и состав хромосомы.
Хроматином (от греч. хрома – окраска, цвет) называют комплекс ДНК и белков, интенсивно окрашивающийся некоторыми красителями и отличающийся по форме от ядрышка. В делящихся клетках молекулы ДНК сильно спирализуются, укорачиваются и приобретают компактные размеры и форму. Такое компактное состояние ДНК называют хромосомами.
Хромосомы – органоиды клеточного ядра, совокупность которых определяет основные наследственные свойства клеток и организмов. Полный набор хромосом в клетке, характерный для данного организма, называется кариотипом. В любой клетке тела большинства животных и растений каждая хромосома представлена дважды: одна из них получена от отца, другая – от матери при слиянии ядер половых клеток в процессе оплодотворения. Такие хромосомы называются гомологичными, набор гомологичных хромосом – диплоидным.
Форма хромосом зависит от положения так называемой первичной перетяжки, или центромеры, – области, к которой во время деления клетки (митоза) прикрепляются нити веретена деления. Центромера делит хромосому на два плеча, которые могут быть одинаковой или разной длины.
Вопрос 4. Как соотносится число хромосом в соматических и половых клетках? Почему число хромосом в половых клетках должно быть вдвое меньше, чем в соматических?
Число хромосом в кариотипе большинства видов живых организмов чётное. Это объясняется тем, что в каждой соматической клетке находятся две одинаковые по форме и размеру хромосомы: одна – из отцовского организма, вторая – из материнского.
Хромосомы, одинаковые по форме и размеру и несущие одинаковые гены, называют гомологичными. Хромосомный набор соматической клетки, в котором каждая хромосома имеет себе пару, носит название двойного (или диплоидного) и обозначается 2n. Из каждой пары гомологичных хромосом в половые клетки попадает только одна хромосома, поэтому хромосомный набор гамет называют одинарным (или гаплоидным) и обозначают 1n.
Вопрос 5. Какие хромосомы называют гомологичными?
Хромосомы, одинаковые по форме и размеру и несущие одинаковые гены, называют гомологичными.
Вопрос 6. Что такое кариотип?
Совокупность количественных (число и размеры) и качественных (форма) признаков хромосомного набора соматической клетки называют кариотипом.
Вопрос 7. Вспомните строение ДНК бактерий. Выберите критерии и сравните наследственный материал про- и эукариотических клеток.
Строение ДНК бактерий аналогично таковому клеток эукариотического типа (растений, животных, грибов). В отличие от бактерий у вирусов геном представлен одной нуклеиновой кислотой – ДНК или РНК. Бактериальные клетки, кроме ДНК, могут иметь генетически полноценные образования функционирующие автономно. Необходимо подчеркнуть, что носителями наследственности бактерий кроме ДНК являются плазмиды и эписомы. В этой связи, любая структура бактериальной клетки, способна к саморепликации.
Бактериальная хромосома представлена одной двунитевой молекулой ДНК кольцевидной формы и называется нуклеотидом. Длина нуклеотида в растянутом виде составляет примерно 1 мм. Нуклеотид – эквивалент ядра. Расположен он в центре бактерии. В отличие от эукариот ядро бактерий не имеет ядерной оболочки, ядрышка и основных белков (гистонов).
Геномы прокариот и эукариот, хотя и имеют определенное сходство, но все же существенно различаются по своей структуре. Геномы прокариот практически целиком состоят из генов и регуляторных последовательностей. В генах прокариот нет интронов. Часто функционально родственные гены прокариот находятся под единым транскрипционным контролем, то есть транскрибируются вместе, составляя оперон.
Геномы эукариот существенно больше геномов бактерий, у дрожжей примерно в 2 раза, а у человека – на три порядка, то есть в тысячу раз. Однако прямой зависимости между количеством ДНК и эволюционной сложностью видов не наблюдается. Достаточно сказать, что геномы некоторых видов амфибий или растений в десять или даже в сто раз превосходят по размеру геном человека. В некоторых случаях близкие виды организмов могут существенно различаться по количеству ДНК. Важным обстоятельством является то, что при переходе от прокариот к эукариотам увеличение генома происходит, главным образом, за счет появления огромного количества некодирующих последовательностей. Действительно, в геноме человека кодирующие области, то есть экзоны, суммарно занимают не более 3%, а по некоторым оценкам около 1% от общей длины ДНК.
Вопрос 8. Используя рисунок 14, расскажите, как осуществляется обмен веществами между ядром и цитоплазмой.
Обмен веществ между ядром и цитоплазмой осуществляется двумя путями. Во-первых, ядерная оболочка пронизана многочисленными порами, через которые происходит обмен молекулами между ядром и цитоплазмой. Во-вторых, поступление веществ из ядра в цитоплазму и обратно может происходить в результате отделения выростов и впячиваний ядерной оболочки.
Пути обмена веществ между ядром и цитоплазмой.
1 — обмен веществ через ядерные поры,
2 — впячивание цитоплазмы внутрь ядра,
3 — впячивание ядерной оболочки,
4 — продвижение ядерной мембраны в эндоплазматическую сеть;
5 — выведение части каналов во внешнее межклеточное пространство.
Вопрос 9. Используя дополнительные источники информации, приведите примеры числа хромосом у разных видов живых организмов. Сделайте вывод, зависит ли степень сложности организации вида от числа хромосом.
Человек Homo sapiens 46
Гориллы Gorilla 48
Волк Canis lupus 78
Кошка Felis catus 38
Осёл Equus asinus 62
Ананас Ananas comosus 50
Картофель Solanum tuberosum 48
Комар Aedes aegypti 6
Наименьшее число хромосом: самки подвида муровьев Myrmecia pilosula имеют пару хромосом на клетку. Самцы имеют только 1 хрососому в каждой клетке.
Наибольшее число: вид папоротников Ophioglossum reticulatum имеет около 630 пар хромосом, или 1260 хромосом на клетку
Верхний предел числа хромосом не зависит от количества ДНК которое в них входит: у американской амфибии Amphiuma ДНК в ~30 раз больше, чем у человека, которая помещается в 14 хромосомах.
Число хромосом не зависит от уровня организации и не всегда указывает на родство: одно и тоже число их может быть у очень далёких друг от друга систематических групп и может сильно отличаться у близких по происхождению видов.
Например величина генома у эукариот обычно гораздо больше, чем у прокариот. Отклонения в величине генома у эукариот гораздо больше, чем у бактерий: от 8.8*10 в 6 степени нуклеиновых пар до 6.9*10 в 11 степени нуклеиновых пар, т. е. приблизительно в 80 тысяч раз. Огромная межвидовая вариация в размере генома среди эукариот не имеет отношения ни к сложности организма, ни к вероятному числу генов, которые этот организм имеет. Например, некоторые одноклеточные обладают гораздо большим количеством ДНК, чем млекопитающие. Отсутствие соответствия между величиной генома и предполагаемым количеством генетической информации, содержащейся внутри генома, известно как парадокс величины генома. Суть этого парадокса в следующем:
а) размеры генома большинства эукариот настолько велики, что их потенциальная информационная емкость намного превышает реальное число генов;
б) виды одного и того же рода могут существенно (в несколько раз) отличаться по величине генома;
в) так называемые «эволюционно примитивные» реликтовые формы («живые ископаемые») по содержанию ДНК на клетку зачастую превосходят представителей эволюционно преуспевающих таксономических групп: почти 35-кратное превышение генома двоякодышаших рыб над геномом человека.
Таким образом величина геномов у представителей разных таксонов вовсе не согласуется с нашими интуитивными представлениями о том, кто из них «выше», а кто «ниже» на эволюционной лестнице. «Судить о степени эволюционной продвинутости по размерам генома столь же правомочно, как оценивать общественное положение человека по его весу». ))
Избыточность величины генома конкретно выражается в наличии многочисленных семейств повторяющейся ДНК. Разнообразие семейств повторяющейся ДНК с трудом поддается систематизации.
Вопрос 10. Согласны ли вы с утверждением, что ядро является важнейшей частью клетки? Ответ обоснуйте.
Ядро – основной компонент клетки, несущей генетическую информации Ядро – располагается в центре, оно является важнейшей частью клетки. Значение ядра: участвует в образовании белка, РНК, рибосом; регуляция формообразования процессов и функции клеток; хранение генетического кода и его точное воспроизведение в ряду клеточного поколения.
Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал функционирует и воспроизводится. Поэтому выпадение или нарушение любой из перечисленных выше функций губительно для клетки в целом. Так нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не будет обеспечивать клеточные функции, в результате чего клетка погибает.
рушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что также губительно для клеток. К такому же результату приведет нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток. Выпадение в результате поражения ядра или в случае нарушений каких-либо регуляторных процессов синтеза любой формы РНК автоматически приведет к остановке синтеза белка в клетке или к грубым его нарушениям.
Источник: resheba.me
Строение и функции ядра
Как правило, эукариотическая клетка имеет одно ядро, но встречаются двуядерные (инфузории) и многоядерные клетки (опалина). Некоторые высокоспециализированные клетки вторично утрачивают ядро (эритроциты млекопитающих, ситовидные трубки покрытосеменных).
Форма ядра — сферическая, эллипсовидная, реже лопастная, бобовидная и др. Диаметр ядра — обычно от 3 до 10 мкм.
Ядро отграничено от цитоплазмы двумя мембранами (каждая из них имеет типичное строение). Между мембранами — узкая щель, заполненная полужидким веществом. В некоторых местах мембраны сливаются друг с другом, образуя поры (3), через которые происходит обмен веществ между ядром и цитоплазмой. Наружная ядерная (1) мембрана со стороны, обращенной в цитоплазму, покрыта рибосомами, придающими ей шероховатость, внутренняя (2) мембрана гладкая. Ядерные мембраны являются частью мембранной системы клетки: выросты наружной ядерной мембраны соединяются с каналами эндоплазматической сети, образуя единую систему сообщающихся каналов.
Кариоплазма (ядерный сок, нуклеоплазма) — внутреннее содержимое ядра, в котором располагаются хроматин и одно или несколько ядрышек. В состав ядерного сока входят различные белки (в том числе ферменты ядра), свободные нуклеотиды.
Ядрышко (4) представляет собой округлое плотное тельце, погруженное в ядерный сок. Количество ядрышек зависит от функционального состояния ядра и варьирует от 1 до 7 и более. Ядрышки обнаруживаются только в неделящихся ядрах, во время митоза они исчезают. Ядрышко образуется на определенных участках хромосом, несущих информацию о структуре рРНК. Такие участки называются ядрышковым организатором и содержат многочисленные копии генов, кодирующих рРНК. Из рРНК и белков, поступающих из цитоплазмы, формируются субъединицы рибосом. Таким образом, ядрышко представляет собой скопление рРНК и рибосомальных субъединиц на разных этапах их формирования.
Хроматин — внутренние нуклеопротеидные структуры ядра, окрашивающиеся некоторыми красителями и отличающиеся по форме от ядрышка. Хроматин имеет вид глыбок, гранул и нитей. Химический состав хроматина: 1) ДНК (30–45%), 2) гистоновые белки (30–50%), 3) негистоновые белки (4–33%), следовательно, хроматин является дезоксирибонуклеопротеидным комплексом (ДНП). В зависимости от функционального состояния хроматина различают: гетерохроматин (5) и эухроматин (6). Эухроматин — генетически активные, гетерохроматин — генетически неактивные участки хроматина. Эухроматин при световой микроскопии не различим, слабо окрашивается и представляет собой деконденсированные (деспирализованные, раскрученные) участки хроматина. Гетерохроматин под световым микроскопом имеет вид глыбок или гранул, интенсивно окрашивается и представляет собой конденсированные (спирализованные, уплотненные) участки хроматина. Хроматин — форма существования генетического материала в интерфазных клетках. Во время деления клетки (митоз, мейоз) хроматин преобразуется в хромосомы.
Функции ядра: 1) хранение наследственной информации и передача ее дочерним клеткам в процессе деления, 2) регуляция жизнедеятельности клетки путем регуляции синтеза различных белков, 3) место образования субъединиц рибосом.
Хромосомы

Хромосомы — это цитологические палочковидные структуры, представляющие собой конденсированный хроматин и появляющиеся в клетке во время митоза или мейоза. Хромосомы и хроматин — различные формы пространственной организации дезоксирибонуклеопротеидного комплекса, соответствующие разным фазам жизненного цикла клетки. Химический состав хромосом такой же, как и хроматина: 1) ДНК (30–45%), 2) гистоновые белки (30–50%), 3) негистоновые белки (4–33%).
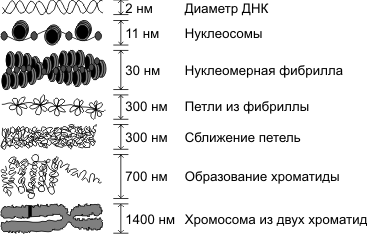
Основу хромосомы составляет одна непрерывная двухцепочечная молекула ДНК; длина ДНК одной хромосомы может достигать нескольких сантиметров. Понятно, что молекула такой длины не может располагаться в клетке в вытянутом виде, а подвергается укладке, приобретая определенную трехмерную структуру, или конформацию. Можно выделить следующие уровни пространственной укладки ДНК и ДНП: 1) нуклеосомный (накручивание ДНК на белковые глобулы), 2) нуклеомерный, 3) хромомерный, 4) хромонемный, 5) хромосомный.
В процессе преобразования хроматина в хромосомы ДНП образует не только спирали и суперспирали, но еще петли и суперпетли. Поэтому процесс формирования хромосом, который происходит в профазу митоза или профазу 1 мейоза, лучше называть не спирализацией, а конденсацией хромосом.
Метафазная хромосома (хромосомы изучаются в метафазу митоза) состоит из двух хроматид (8). Любая хромосома имеет первичную перетяжку (центромеру) (5), которая делит хромосому на плечи. Некоторые хромосомы имеют вторичную перетяжку (6) и спутник (7). Спутник — участок короткого плеча, отделяемый вторичной перетяжкой. Хромосомы, имеющие спутник, называются спутничными (3). Концы хромосом называются теломерами (9). В зависимости от положения центромеры выделяют: а) метацентрические (равноплечие) (1), б) субметацентрические (умеренно неравноплечие) (2), в) акроцентрические (резко неравноплечие) хромосомы (3, 4).
Соматические клетки содержат диплоидный (двойной — 2n) набор хромосом, половые клетки — гаплоидный (одинарный — n). Диплоидный набор аскариды равен 2, дрозофилы — 8, шимпанзе — 48, речного рака — 196. Хромосомы диплоидного набора разбиваются на пары; хромосомы одной пары имеют одинаковое строение, размеры, набор генов и называются гомологичными.
Кариотип — совокупность сведений о числе, размерах и строении метафазных хромосом. Идиограмма — графическое изображение кариотипа. У представителей разных видов кариотипы разные, одного вида — одинаковые. Аутосомы — хромосомы, одинаковые для мужского и женского кариотипов. Половые хромосомы — хромосомы, по которым мужской кариотип отличается от женского.
Хромосомный набор человека (2n = 46, n = 23) содержит 22 пары аутосом и 1 пару половых хромосом. Аутосомы распределены по группам и пронумерованы:
| Группа | Число пар | Номер | Размер | Форма |
|---|---|---|---|---|
| A | 3 | 1, 2, 3 | Крупные | 1, 3 — метацентрические, 2 — субметацентрические |
| B | 2 | 4, 5 | Крупные | Субметацентрические |
| C | 7 | 6, 7, 8, 9, 10, 11, 12 | Средние | Субметацентрические |
| D | 3 | 13, 14, 15 | Средние | Акроцентрические, спутничные (вторичная перетяжка в коротком плече) |
| E | 3 | 16, 17, 18 | Мелкие | Субметацентрические |
| F | 2 | 19, 20 | Мелкие | Метацентрические |
| G | 2 | 21, 22 | Мелкие | Акроцентрические, спутничные (вторичная перетяжка в коротком плече) |
Половые хромосомы не относятся ни к одной из групп и не имеют номера. Половые хромосомы женщины — ХХ, мужчины — ХУ. Х-хромосома — средняя субметацентрическая, У-хромосома — мелкая акроцентрическая.
В области вторичных перетяжек хромосом групп D и G находятся копии генов, несущих информацию о строении рРНК, поэтому хромосомы групп D и G называются ядрышкообразующими.
Функции хромосом: 1) хранение наследственной информации, 2) передача генетического материала от материнской клетки к дочерним.
Источник: licey.net
ХРОМАТИН — материальный субстрат хромосом, представляющий собой многокомпонентную систему молекул, находящихся в определенных пространственных, химических и физических взаимоотношениях.
Основным структурным и химическим компонентом хроматина служит комплекс дезоксирибонуклеиновой кислоты (см.) с гистонами (см.) и негистоновыми белками (см. Нуклеопротеиды), иногда — с протаминами (см.). Другие компоненты хроматина — РНК (см. Рибонуклеиновые кислоты), липиды (см.), углеводы (см.), неорганические вещества прямо или косвенно связаны с белками (см.). Количественные соотношения компонентов хроматина существенно зависят от типа клеток; их относительное содержание чаще всего соответствует следующим величинам: ДНК 30—45%, гистоны 30—50%, негистоновые белки 2—35%, РНК и другие компоненты 1 —10%.
Термин «хроматин» был введен в 1880 году немецким ученым В. Флеммингом для обозначения окрашивающихся структур фиксированных ядер клеток (за исключением ядрышек). Преобладающая часть таких структур окрашивается основными красителями (базохроматин), а некоторые — кислотными (оксихроматин). Выделяемые в ядре хроматиновые структуры имеют вид глыбок или сети фибрилл, различающихся в одной клетке и в клетках разных объектов по степени дисперсности. Наиболее интенсивно окрашивающиеся глыбки — хромоцентры (кариосомы) иногда называли ложными ядрышками. Хромо центры имеют, по-видимому, повышенную адгезивную способность, поскольку они легко входят в контакт с ядерной оболочкой, а также агрегируют друг с другом.
Структура хроматина в ядрах интерфазных клеток, то есть клеток, находящихся в периоде между следующими друг за другом митозами (см.), зависит от стадии развития организма (см. Онтогенез, Эмбриональное развитие). У ряда исследованных объектов в первых 2—4-х бластомерах хроматиновые структуры не выявляются, на стадии 8—10 бластомеров в ядре выявляются мелкие хроматиновые глыбки, приобретающие в неделящихся дифференцированных клетках высокоспецифичный характер для каждого типа клеток (см. Деление клетки). В процессе старения этих клеток наблюдают усиление конденсации хроматина.
Изоэлектрическая точка (см.) хроматина зависит от количества белков, входящих в комплекс с ДНК, и находится в интервале значений pH 3,0—5,0. Патологические изменения хроматина как морфологические структуры сопровождаются изменениями pH, при которых находится изоэлектрическая точка. Это отмечают, например, при воздействиях ионизирующего излучения, старении и др. При различных патологических состояниях может меняться и степень дисперсности хроматина. Так, опухолевые клетки характеризуются наличием большого числа хромоцентров, имеющих иногда достаточно крупные размеры; при болезни Дауна (см. Дауна болезнь) хроматин по сравнению с нормой более конденсирован, изменены константы его связывания с красителями; при синдроме Блума (см. Пойкилодермия) хроматин имеет пылевидную или сегментированную структуру. При некоторых видах патологии отмечено усиление конденсации хроматина и концентрация его крупных глыбок на внутренней поверхности ядерной оболочки (гиперхроматоз).
Структуры хроматина и половой хроматин (см.) наблюдают с помощью световой микроскопии. Форма и размер этих структур зависят от способа фиксации клеток. Это свидетельствует о том, что выявляемая после фиксации морфология хроматина отражает не его истинную структуру в живой клетке, а лишь возможность разных способов его организации. В ядрах живых клеток, как правило, компоненты, соответствующие хроматиновым структурам, не выявляются. Однако незначительные повреждения (раздражения) в ряде случаев приводят к обратимому появлению таких структур в прежде гомогенном ядре (напр., при воздействии наркотических анальгетиков и др.). Известен и противоположный эффект — обратимая «гомогенизация» структур, выявляемая в норме в ядрах живых клеток. Естественно, что оптическая гомогенность ядра не тождественна структурной гомогенности хроматина на уровнях более низких, чем позволяет видеть разрешающая способность световой микроскопии. Поэтому сейчас термин «хроматин» утрачивает свое морфологическое содержание, его чаще относят к химическому субстрату хромосом (см.) — сложному комплексу биополимеров. Организующие этот комплекс в единую систему в основном слабые (нековалентные) взаимодействия, равно как и конформация (см.) образующих его молекул, существенным образом зависят от хим. состава, количественного соотношения взаимодействующих компонентов и внешних факторов. Это определяет возможность различных способов организации комплекса в целом и (или) благодаря структурной динамике организации его отдельных структурных компонентов. Полагают, что набор таких способов организации (состояний) ограничен, а переходы между ними имеют характер фазовых переходов. Реализация состояния хроматина, по тем или иным причинам не соответствующего состоянию данной клетки в норме, является признаком патологии.
Установлено существование, по крайней мере, двух классов хроматина: 1) эухроматина, который деконденсируется во время интерфазы и конденсируется в митозе; 2) гетерохроматина, который остается компактным не только в митозе, но и в интерфазе, где его микроскопически идентифицируют в виде хромоцентров. Эухроматин является основной информационной частью генома, в которой преимущественно локализованы структурные гены с соответствующими регуляторными областями. Для гетерохроматина характерна поздняя репликация (см.) ДНК, входящей в его состав. В отличие от эухроматина гетерохроматин в структурном отношении более лабилен: иногда наблюдают его деконденсацию при голодании, действии низких температур и др. Установлено, что при воздействии ряда мутагенных факторов (см. Мутагены) химической и физической природы структурные повреждения чаще локализуются в гетерохроматиновых областях хромосом. Различают два типа гетерохроматина. Первым из них является структурный, постоянно конденсированный хроматин. Как правило, в нем не содержится генов (см. Ген), его ДНК представлена в основном короткими повторяющимися нуклеотидными последовательностями (у некоторых организмов — сателлитной ДНК). При пространственном сближении в результате хромосомных перестроек участков структурного гетерохроматина и эухроматина в ряде случаев ингибируется фенотипическое проявление генов (так называемый эффект положения гена). Активация генов, локализованных в эухроматине, при пространственном разобщении последнего с гетерохроматином может быть, согласно некоторым представлениям, одной из причин активации онкогенов, локализованных в ДНК хромосомы. В целом роль структурного гетерохроматина недостаточно ясна. Полагают, что он существен для процессов конъюгации хромосом (см.), взаимного расположения хромосом в ядре, прикрепления участков хромосом к ядерной оболочке, укладки хроматиновых фибрилл, защиты жизненно важных элементов хромосом, сближения ядрышкообразующих хромосом, эволюции кариотипа и др. Таким образом, предполагаемая роль структурного гетерохроматина заключается в регуляции пространственной организации и соответственно — функциональной активности хромосом.
У человека структурный гетеро-хроматин локализован в центромерных участках всех хромосом, в районах вторичных перетяжек хромосом 1, 9, 16-й пар, коротких плечах акроцентрических хромосом, в дистальной части длинного плеча Y -хромосомы и обрамляет блоки генов рибосомной РНК (ядрышкообразующие районы). На долю структурного гетерохроматина у человека приходится 10—15% всего хроматина. У разных лиц количество структурного гетерохроматина варьирует даже в пределах гомологичных хромосом. Обнаружено, что полиморфные варианты структурного гетерохроматина (см. Полиморфизм в генетике) у людей могут коррелировать с некоторыми наследственными заболеваниями, а возможно определять их или указывать на предрасположенность к ним.
Вторым типом гетерохроматина принято считать факультативный гетерохроматин, или инактивированный эухроматин. Этот тип хроматина сходен с гетерохроматином только в морфол. отношении: микроскопически он выявляется в интерфазном ядре в виде интенсивно красящихся глыбок разного размера. Основываясь на молекулярной организации и функциях, его правильнее считать одним из типов эухроматина. Он содержит структурные гены, фенотипически инактивированные путем конденсации (гетерохроматини-зации) эухроматина. Одним из типичных примеров факультативного гетерохроматина являются тельца Барра (X-хроматин).
Таким образом, функционирование хроматина как системы, в которой происходит начальный этап реализации наследственной информации, в значительной степени определяется пространственным распределением ее взаимозависимых конденсированных и де-конденсированных зон (согласно представлениям о физических процессах, лежащих в основе самоорганизации пространственной структуры хроматина,— микрофазовое расслоение системы). Распределение конденсированных и деконденсированных зон является отражением состояния системы в целом, что не исключает, однако, относительной автономности этих участков в ряде процессов. Известны случаи, когда путем конденсации хроматина осуществляется инактивация целых хромосом (например, одной из X-хромосом у женщин) или почти всего генома (напр., в эритроцитах птиц). В большинстве типов клеток доля активного хроматина составляет 2— 15%. По данным молекулярно-биол. анализа, в ряде случаев инактивация связана с появлением определенных подфракций гистона Н1 или замещением последнего другими гистонами, в частности гистоном Н5 (см. Нуклеопротеиды). В сперматозоидах некоторых животных репрессия генома реализуется на фоне замещения гистонов протаминами или подобными им белками.
Существенную роль в организации транскрипции (см.), в том числе через дифференциальную деконденсацию хроматина, отводят негистоновым белкам хроматина (НГБ). В их число входят также ферментные комплексы, ответственные за репарацию (см. Репарация генетических повреждений), репликацию, транскрипцию и модификацию нуклеиновых кислот (см.) и за некоторые ферментативные превращения ряда хромосомных белков. В ядрах клеток, в которых не происходит активной транскрипции, количество негистоновых белков хроматина существенно уменьшено. Например, зрелые гаметы в значительной степени освобождены от таких белков. Полагают, что в организации или поддержании транскрипции принимают участие негистоновые белки хроматина, прочно связанные с ДНК, среди которых, по-видимому, находится компонент, специфически связывающий комплекс гормон — рецептор, а также тесно связанные с нуклеосомами белки HMG14 и HMG17. Последние способны ингибировать деацетилирование гистонов, а этот процесс наряду с недометилированием ДНК представляет собой модификации, характерные для компонентов активных участков хроматина.
Важным для структурных переходов хроматина является способность белка хроматина А24 к расщеплению на гистон Н2а и полипептид убиквитин. Общей характеристикой участков транскрипционно активного хроматина из разных источников является повышенная чувствительность их ДНК к воздействию ряда нуклеаз (см.). При активации транскрипции такая чувствительность распространяется на участок молекулы ДНК в составе хроматина по протяженности примерно на два порядка больше, чем занимает ген. Все изложенное выше свидетельствует о значении в организации транскрипции более высоких уровней упаковки хроматина, чем его элементарная фибрилла, видимая в электронный микроскоп. Последняя при участии гистона Н1, расположенного наряду с негистоновыми белками хроматина HMG1 и HMG2 в основном на межнуклеосомной ДНК, представляет волокно диаметром около 10 нм. При этом монотонность нуклеосомной организации дезоксирибонуклеопротеидного (ДНП) волокна может нарушаться благодаря структурной динамике нуклеосом (см. Клетка), модификации гистонов при их фосфорилировании, ацетилировании, метилировании и рибозилировании.
Существенную роль отводят меж-молекулярным контактам, способным регулировать конденсацию ДНК на уровне нуклеосом. Нек-рые структурные переходы нуклеосом происходят при изменении ионной силы среды. В ядре клетки количество низкомолекулярных противоионов (ионов К+, Na+ и др.) по порядку величины равно числу фиксированных на макромолекулах (например, фосфатные группы ДНК) зарядов. Поэтому небольшие колебания в абсолютном количестве низкомолекулярных противоионов в ядре (например, при увеличении или уменьшении объема последнего) должны вызвать структурные переходы нуклеосом. Наконец, гистон Н1 может замещаться другими гистонами или их комплексами, имеющими большее сродство к ДНК, с соответствующей реорганизацией структуры фибриллы. Таким образом, возможность различных способов упаковки хроматина заложена уже на уровне различных полиморфных структурных вариантов элементарной фибриллы хроматина. Стабильность следующего уровня организации хроматина — неравномерных по диаметру (20—30 нм) фибрилл — обеспечивается, по-видимому, и гистоном Н1. Дальнейшая упаковка хроматиновых фибрилл реализуется, как полагают, путем самоорганизации системы с образованием конденсированных (глобулярных) зон и петель или независимых суперспирализованных областей (доменов). Домены характеризуются участком двойной спирали ДНК, специальным образом расположенным в пространстве, концы этой двойной спирали фиксированы, что ограничивает или исключает возможность ее вращения. Длина петли ДНК по контуру для разных объектов соответствует мол. весу (массе) ДНК порядка 10 000000— 100 000000. Изменение степени суперспирализации ДНК является еще одним важным фактором регуляции процессов экспрессии генов (см. Экспрессивность гена) через модификацию надмолекулярных систем хроматина. Суперспирализация ДНК изменяется также при действии ионизирующего излучения, некоторых химических соединений, активации нуклеаз и др. Указанные факторы вызывают однонитевые разрывы в молекулах ДНК, что приводит к релаксации в отдельных петлях ее исходной суперспиральной структуры. Этот процесс может вызывать перераспределение белков хроматина, поскольку ряд белков имеет различные константы связывания с линейной, кольцевой и суперспиральной ДНК.
Воздействие агентов, вызывающих диссоциацию белков, в частности гистонов хроматина (некоторые химимечсие мутагены, ионизирующие излучение, высокие концентрации солей, ионов водорода и др.), также приводит к изменению степени суперспиральности, поскольку сам процесс образования нуклеосом связан с реорганизацией суперспирали ДНК.
Полагают, что динамические возможности структуры хроматина нельзя рассматривать только как один из факторов, регулирующих транскрипцию. Действие всех остальных факторов регуляции, как внутри-, так и внеклеточных, реализуется через создание структуры хроматина, специфичной для каждого типа клеток, различающихся по характеру синтеза РНК. В этой связи все воздействия, изменяющие нормальные взаимоотношения между компонентами хроматина и тем самым — его структуру, должны приводить к патологическому функционированию этой системы. Существенное значение имеют изменения структуры хроматина, предрасполагающие к последующему генетическому неблагополучию. Так, полагают, что важное значение может иметь реализация состояний хроматина, при которых снижена вероятность узнавания ферментами репарации повреждений ДНК — явления, которое, по-видимому, служит одной из ведущих причин феномена нестабильности хромосом и характерной для них группы наследственных болезней (см. Хромосомные болезни). Отмечена связь некоторых изменений структуры хроматина с увеличением частоты конъюгации негомологичных хромосом — одной из возможных причин анеуплоидий (см. Мутация). При действии генетически опасных агентов на клетки и организмы кроме генетических повреждений самой ДНК (генные мутации) и указанных выше перестроек структуры хроматина как системы возникают многочисленные нарушения во взаимодействиях между компонентами хроматина: частичная диссоциация белков хроматина, образование межмолекулярных «сшивок» между ДНК и белками, распад фибриллы хроматина на нуклеосомы и др., что в свою очередь усиливает патологический эффект такого агента.
Библиогр.: Андрееве. Г. и Спитковский Д. М. Биофизические модели самоорганизации пространственной структуры хроматина, Докл. АН СССР, т. 269, № 6, с. 1500, 1983; Г е о р г и е в Г. П. и БакаевВ. В. Три уровня структурной организации хромосом эукариот, Молек. биол., т. 12, № 6, с. 1205, 1978, библиогр.; H е й ф а х А. А. и Т и м о ф e е в а М. Я. Проблемы регуляции в молекулярной биологии развития, М., 1978; Прокофье-ва-Бе льговская А. А. Значение негистоновых белков в преобразованиях и генетическом функционировании хромосом, Молек. биол., т. 16, Na 4, с. 771, 1982; Теоретические проблемы медицинской генетики, под ред. А. Ф. Захарова, с. 52, М., 1979; Chromatin structure and function, ed. by C. A. Nicolini, N. Y. —L., 1979.
Д. М. Спитковский, H. А. Ляпунова.
Источник: xn--90aw5c.xn--c1avg