Первые эукариоты
Симбиогенез — основная гипотеза происхождения эукариот
Существует несколько гипотез о путях возникновения эукариотических клеток. Наиболее популярная — симбиотическая гипотеза (симбиогенез). Согласно ей, эукариоты произошли в результате объединения в одной клетке разных прокариот, которые сначала вступили в симбиоз, а затем, все более специализируясь, стали органоидами единого организма-клетки. Как минимум симбиотическое происхождение имеют митохондрии и хлоропласты (пластиды вообще). Произошли они от бактериальных симбионтов.
Клеткой-хозяином мог быть относительно крупный анаэробный гетеротрофный прокариот, похожий на амебу. В отличие от других, он мог приобрести способность питаться путем фаго- и пиноцитоза, что позволяло ему захватывать других прокариот. Они не все переваривались, а снабжали хозяина продуктами своей жизнедеятельности). В свою очередь, получали от него питательные вещества.
Митохондрии произошли от аэробных бактерий и позволили клетке-хозяину перейти к аэробному дыханию, которое не только намного эффективней, но и облегчает существование в атмосфере, содержащей достаточно большое количество кислорода. В такой среде аэробные организмы получают преимущество над анаэробными.
Позже в некоторых клетках поселились похожие на ныне живущих синезеленых водорослей (цианобактерий) древние прокариоты. Они стали хлоропластами, дав начало эволюционной ветви растений.

Кроме митохондрий и пластид симбиотическое происхождение могут иметь жгутики эукариот. В них превратились симбионты-бактерии наподобие современных спирохет, имеющих жгутик. Считается, что в последствии из базальных тел жгутиков произошли центриоли, столь важные структуры для механизма клеточного деления эукариот.
Эндоплазматическая сеть, комплекс Гольджи, пузырьки и вакуоли могли произойти от наружной мембраны ядерной оболочки. С другой точки зрения, некоторые из перечисленных органелл могли возникнуть путем упрощения митохондрий или пластид.
Во многом неясным остается вопрос происхождения ядра. Могло ли оно также образоваться из прокариота-симбионта? Количество ДНК в ядре современных эукариот во много раз превышает его количество в митохондриях и хлоропластах. Возможно часть генетической информации последних со временем переместилась в ядро. Также в процессе эволюции происходило дальнейшее увеличение размера ядерного генома.
Кроме того в симбиотической гипотезе происхождения эукариот не все так однозначно с клеткой-хозяином. Им мог и не быть один вид прокариот. Используя методы сравнения геномов, ученые делают вывод, что клетка-хозяин близок к археям, при этом сочетает в себе признаки архей и ряда неродственных групп бактерий. Отсюда можно сделать вывод, что появление эукариот происходило в сложном сообществе прокариот. При этом процесс скорее всего начался с метаногенной археи, вступавшей в симбиоз с другими прокариотами, что было вызвано необходимостью обитания в кислородной среде. Появление фагоцитоза способствовало притоку чужих генов, а ядро образовалось для защиты генетического материала.
Молекулярный анализ показал, что различные белки эукариот происходят от разных групп прокариот.
Доказательства симбиогенеза
В пользу симбиотического происхождения эукариот говорит то, что митохондрии и хлоропласты имеют собственную ДНК, причем кольцевую и не связанную с белками (также обстоит дело у прокариот). Однако в генах митохондрий и пластид есть интроны, чего нет у прокариот.
Пластиды и митохондрии не воспроизводятся клеткой с нуля. Они образуются из ранее существующих таких же органелл путем их деления и последующего роста.
В настоящее время существуют амебы, у которых нет митохондрий, а вместо них есть бактерии симбионты. Также есть простейшие, сожительствующие с одноклеточными водорослями, выполняющими в клетке-хозяине роль хлоропластов.
Инвагинационная гипотеза происхождения эукариот
Кроме симбиогенеза существуют и другие взгляды на происхождение эукариот. Например, инвагинационная гипотеза. Согласно ей, предком эукариотической клетки был не анаэробный, а аэробный прокариот. К такой клетке могли прикрепляться другие прокариоты. Потом их геномы объединялись.
Ядро, митохондрии и пластиды возникли путем впячивания и отшнуровывания участков клеточной мембраны. В эти структуры попадала чужеродная ДНК.
Усложнение генома происходило в процессе дальнейшей эволюции.
Инвагинационная гипотеза происхождения эукариот хорошо объясняет наличие двойной мембраны у органелл. Однако она не объясняет, почему система биосинтеза белка в хлоропластах и митохондриях сходна с прокариотической, в то время как таковая в ядерно-цитоплазматическом комплексе имеет ключевые отличия.
Причины эволюции эукариот
Все разнообразие жизни на Земле (от простейших до покрытосеменных и млекопитающих) дали клетки эукариотического, а не прокариотического типа. Возникает вопрос, почему? Очевидно, ряд особенностей, возникших у эукариот, существенно повысили их эволюционные возможности.
Во-первых, у эукариот есть ядерный геном, который во много раз превосходит количество ДНК у прокариот. При этом эукариотические клетки диплоидны, кроме этого в каждом гаплоидном наборе определенные гены многократно повторяются. Все это обеспечивает, с одной стороны, большие масштабы для мутационной изменчивости, а с другой — уменьшает угрозу резкого снижения жизнеспособности в результате вредной мутации. Таким образом, эукариоты, в отличие от прокариот, обладают резервом наследственной изменчивости.
Эукариотические клетки имеют более сложный механизм регуляции жизнедеятельности, у них существенно больше различных регуляторных генов. Кроме того, молекулы ДНК образовали комплексы с белками, что позволило наследственному материалу упаковываться и распаковываться. Все вместе это дало возможность считывать информацию частями, в разных сочетаниях и количестве, в разное время. (Если в клетках прокариот транскрибируется почти вся информация генома, то в эукариотических клетках обычно менее половины.) Благодаря этому эукариоты могли специализироваться, лучше приспосабливаться.
У эукариот появились митоз, а затем и мейоз. Митоз позволяет воспроизводить генетически сходные клетки, а мейоз сильно увеличивает комбинативную изменчивость, что ускоряет эволюцию.
Большую роль в процветании эукариот сыграло приобретенное их предком аэробное дыхание (хотя оно есть и у многих прокариот).
На заре своей эволюции эукариоты обзавелись эластичной оболочкой, обеспечивавшей возможность фагоцитоза, и жгутиками, позволившими им двигаться. Это дало возможность эффективней питаться.
Источник: biology.su
Появление эукариот – важнейшее событие. Изменило структуру биосферы и открыло принципиально новые возможности для прогрессивной эволюции. Эукариотическая клетка является результатом долгой эволюции мира прокариот, мира, в котором разнообразные микробы приспосабливались друг к другу и искали способы эффективной кооперации.
набросок хронологии (повторение)
Фотосинтезирующий прокариотический комплекс Chlorochromatium aggregatum.
Эукариоты возникли в результате симбиоза нескольких видов прокариот. Прокариоты вообще весьма склонны к симбиозу (см. главу 3 в книге «Рождение сложности»). Вот интересная симбиотическая система, известная под названием Chlorochromatium aggregatum. Живет в глубоких озерах, где есть на глубине бескислородные условия. Центральный компонент – подвижная гетеротрофная бета-протеобактерия. Вокруг нее стопками располагаются от 10 до 60 фотосинтезирующих зеленых серных бактерий. Все компоненты соединены выростами наружной мембраны центральной бактерии. Смысл содружества в том, что подвижная бета-протеобактерия перетаскивает всю компанию в места, благоприятные для жизни привередливых серных бактерий, а серные бактерии занимаются фотосинтезом и обеспечивают пищей и себя, и бета-протеобактерию. Может быть, какие-то древние микробные ассоциации примерно такого типа и были предками эукариот.
Теория симбиогенеза. Мережковский, Маргулис. Митохондрии – потомки альфа-протеобактерий, пластиды – потомки цианобактерий. Труднее понять, кто был предком всего остального, то есть цитоплазмы и ядра. Ядро и цитоплазма эукариот сочетает в себе признаки архей и бактерий, а также имеет множество уникальных особенностей.
О митохондриях. Возможно, именно приобретение митохондрий (а не ядра) было ключевым моментом в становлении эукариот. Большинство генов предков митохондрий было перенесено в ядро, где они попали под контроль ядерных регуляторных систем. Эти ядерные гены митохондриального происхождения кодируют не только белки митохондрий, но и многие белки, работающие в цитоплазме. Это говорит о том, что митохондриальный симбионт сыграл более важную роль в формировании эукариотической клетки, чем предполагалось.
Совместное существование в одной клетке двух разных геномов требовало развития эффективной системы их регуляции. А для того, чтобы эффективно управлять работой большого генома, необходимо изолировать геном от цитоплазмы, в которой протекает обмен веществ и идут тысячи химических реакций. Ядерная оболочка как раз и отделяет геном от бурных химических процессов цитоплазмы. Приобретение симбионтов (митохондрий) могло стать важным стимулом в развитии ядра и генно-регуляторных систем.
То же относится и к половому размножению. Без полового размножения можно жить до тех пор, пока геном у вас достаточно маленький. Организмы с большим геномом, но лишенные полового размножения, обречены на быстрое вымирание, за редчайшими исключениями.
Альфапротеобактерии – к этой группе относились предки митохондрий.
Родоспириллум – удивительный микроорганизм, который может жить и за счет фотосинтеза, в том числе и в анаэробных условиях, и как аэробный гетеротроф, и даже как аэробный хемоавтотроф. Он может, например, расти за счет окисления угарного газа СО, не используя никаких других источников энергии. Вдобавок ко всему этому, он умеет еще и фиксировать атмосферный азот. То есть это в высшей степени универсальный организм.
Иммунная система принимает митохондрии за бактерии. Когда при травме в кровь попадают разрушенные митохондрии, из них высвобождаются характерные молекулы, которые встречаются только у бактерий и у митохондрий (кольцевая ДНК бактериального типа и белки, несущие на одном из своих концов особую модифицированную аминокислоту формилметионин). Это связано с тем, что аппарат синтеза белка в митохондриях остался таким же, как у бактерий. Клетки иммунной системы – нейтрофилы – реагируют на эти митохондриальные вещества точно так же, как на бактериальные, и при помощи тех же рецепторов. Это ярчайшее подтверждение бактериальной природы митохондрий.
Главная функция митохондрий – кислородное дыхание. Скорее всего, стимулом для объединения анаэробного предка ядра и цитоплазмы с «протомитохондрией» была необходимость защититься от токсического действия кислорода.
Откуда взялись у бактерий, и в том числе у альфапротеобактерий, молекулярные системы, необходимые для кислородного дыхания? Похоже, в их основу были положены молекулярные системы фотосинтеза. Электронно-транспортная цепь, сформировавшаяся у бактерий как часть фотосинтетического аппарата, была адаптирована для кислородного дыхания. У некоторых бактерий до сих пор участки электронно-транспортных цепей используются одновременно и в фотосинтезе, и в дыхании. Скорее всего предками митохондрий были аэробные гетеротрофные альфа-протеобактерии, которые, в свою очередь, произошли от фотосинтезирующих альфа-протеобактерий, таких как родоспириллум.
Число общих и уникальных белковых доменов у архей, бактерий и эукариот. Белковый домен – это часть белковой молекулы, имеющая определенную функцию и характерную структуру, то есть последовательность аминокислот. Каждый белок, как правило, содержит один или несколько таких структурно-функциональных блоков, или доменов.
4,5 тысячи белковых доменов, которые есть у эукариот, можно разделить на 4 группы: 1) имеющиеся только у эукариот, 2) общие для всех трех надцарств, 3) общие для эукариот и бактерий, но отсутствующие у архей; 4) общие для эукариот и архей, но отсутствующие у бактерий. Мы рассмотрим две последние группы (они на рисунке выделены цветом), поскольку для этих белков можно с определенной уверенностью говорить об их происхождении: соответственно бактериальном или архейном.
Ключевой момент в том, что эукариотические домены, предположительно унаследованные от бактерий и от архей, имеют существенно разные функции. Домены, унаследованные от архей (их функциональный спектр показан на левом графике), играют ключевую роль в жизни эукариотической клетки. Среди них преобладают домены, связанные с хранением, воспроизведением, организацией и считыванием генетической информации. Большинство "архейных" доменов относится к тем функциональным группам, в пределах которых горизонтальный обмен генами у прокариот происходит реже всего. Видимо, эукариоты получили этот комплекс путем прямого (вертикального) наследования от архей.
Среди доменов бактериального происхождения тоже есть белки, связанные с информационными процессами, но их мало. Большинство из них работает только в митохондриях или пластидах. Эукариотические рибосомы цитоплазмы имеют архейное происхождение, рибосомы митохондрий и пластид имеют бактериальное происхождение.
Среди бактериальных доменов эукариот значительно выше доля сигнально-регуляторных белков. От бактерий эукариоты унаследовали многие белки, ответственные за механизмы реагирования клетки на факторы внешней среды. А также – многие белки, связанные с обменом веществ (подробнее см. в главе 3 «Рождения сложности»).
Эукариоты имеют:
· Архейную «сердцевину» (механизмы работы с генетической информацией и синтеза белка)
· Бактериальную «периферию» (обмен веществ и сигнально-регуляторные системы)
· Простейший сценарий: АРХЕЯ проглотила БАКТЕРИЙ (предков митохондрий и пластид) и все свои бактериальные признаки приобрела от них.
· Этот сценарий слишком прост, потому что у эукариот много бактериальных белков, которые не могли быть заимствованы у предков митохондрий или пластид.
У эукариот много «бактериальных» доменов, не характерных ни для цианобактерий (предков пластид), ни для альфапротеобактерий (предков митохондрий). Они были получены от каких-то других бактерий.
Птицы и динозавры. Реконструировать прото-эукариот трудно. Ясно, что та группа древних прокариот, которая дала начало ядру и цитоплазме, обладала рядом уникальных особенностей, которых нет у прокариот, доживших до наших дней. И когда мы пытаемся реконструировать облик этого предка, мы сталкиваемся с тем, что простор для гипотез оказывается слишком большим.
Аналогия. Известно, что птицы произошли от динозавров, причем не от каких-то неизвестных динозавров, а от вполне определенной группы – манирапторых динозавров, которые относятся к тероподам, а тероподы в свою очередь – это одна из групп ящеротазовых динозавров. Найдено много переходных форм между нелетающими динозаврами и птицами.
Но что бы мы могли сказать о предках птиц, если бы ископаемой летописи не было? В лучшем случае мы бы выяснили, что ближайшей родней птиц являются крокодилы. Но смогли бы мы воссоздать облик прямых предков птиц, то есть динозавров? Вряд ли. Но именно в таком положении мы и находимся, когда пытаемся восстановить облик предка ядра и цитоплазмы. Ясно, что это была группа неких прокариотических динозавров, группа вымершая и не оставившая, в отличие от настоящих динозавров, внятных следов в геологической летописи. Современные археи по отношению к эукариотам – это как современные крокодилы по отношению к птицам. Попробуйте восстановить строение динозавров, зная только птиц и крокодилов.
Аргумент в пользу того, что в докембрии жило много всяких микробов, не похожих на нынешние. Протерозойские строматолиты были намного сложнее и разнообразнее современных. Строматолиты – продукт жизнедеятельности микробных сообществ. Не значит ли это, что и протерозойские микробы были разнообразнее современных, и что многие группы протерозойских микробов просто не дожили до наших дней?
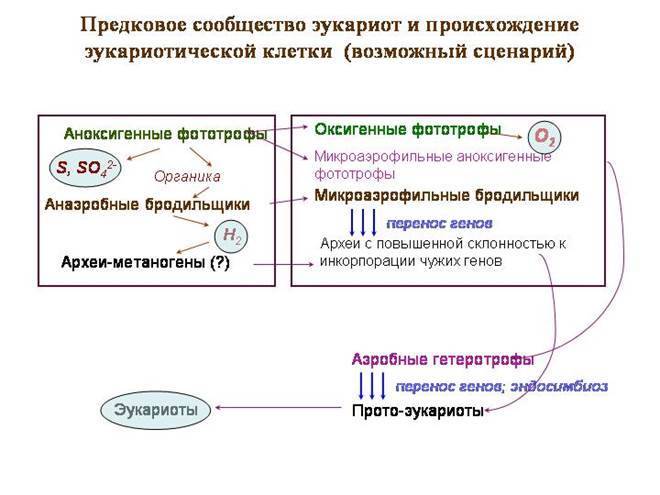
Предковое сообщество эукариот и происхождение эукариотической клетки (возможный сценарий)
Гипотетическое «предковое сообщество» — типичный бактериальный мат, только в его верхнем жили предки цианобактерий, еще не перешедшие к оксигенному фотосинтезу. Они занимались аноксигенным фотосинтезом.Донором электронов служила не вода, а сероводород. В качестве побочного продукта выделялись сера и сульфаты.
Во втором слое обитали пурпурные фотосинтезирующие бактерии, в том числе – альфапротеобактерии, предки митохондрий. Пурпурные бактерии используют длинноволновой свет (красный и инфракрасный). Эти волны обладают лучшей проникающей способностью. Пурпурные бактерии и сейчас часто живут под слоем цианобактерий. Пурпурные альфапротеобактерии тоже используют в качестве донора электрона сероводород.
В третьем слое были бактерии-бродильщики, перерабатывающие органику; некоторые из них в качестве отходов выделяли водород. Это создавало базу для сульфатредуцирующих бактерий. Там могли быть и метаногенные археи. Среди обитавших здесь архей были и предки ядра и цитоплазмы.
Начало кризисным событиям положил переход цианобактерий к кислородному фотосинтезу. В качестве донора электрона цианобактерии начали использовать вместо сероводорода обычную воду. Это открывало большие возможности, но имело и негативные последствия. Вместо серы и сульфатов при фотосинтезе стал выделяться кислород – вещество крайне токсичное для всех древних обитателей земли.
Первыми с этим ядом столкнулись его производители – цианобактерии. Они же, вероятно, первыми стали вырабатывать средства защиты от него. Электронно-транспортные цепи, служившие для фотосинтеза, были модифицированы и начали служить для аэробного дыхания. Изначальная цель, видимо, заключалась не в получении энергии, а только в нейтрализации кислорода.
Вскоре и обитателям второго слоя сообщества – пурпурным бактериям – пришлось вырабатывать аналогичные системы защиты. Так же, как и цианобактерии, они сформировали системы аэробного дыхания на основе фотосинтетических систем. Именно у пурпурных альфапротеобактерий развилась наиболее совершенная дыхательная цепь, которая ныне функционирует в митохондриях эукариот.
В третьем слое сообщества появление свободного кислорода должно было вызвать кризис. Метаногены и многие сульфатредукторы утилизируют молекулярный водород при помощи ферментов-гидрогеназ. Такие микробы не могут жить в аэробных условиях, потому что кислород ингибирует гидрогеназы. Многие бактерии, выделяющие водород, в свою очередь, не растут в среде, где нет микроорганизмов, его утилизирующих. Из бродильщиков в составе сообщества, по-видимому, остались формы, выделяющие в качестве конечных продуктов низкоорганические соединения (пируват, лактат, ацетат и т.п.). Эти бродильщики выработали свои средства защиты от кислорода, менее эвффективные. К числу выживших относились и археи – предки ядра и цитоплазмы.
Может быть, в этот кризисный момент и произошло ключевое событие – ослабление генетической изоляции у предков эукариот и начало активного заимствования чужих генов. Прото-эукариоты инкорпорировали гены разных бродильщиков до тех пор, пока не стали сами микроаэрофильными бродильщиками, сбраживающими углеводы до пирувата и молочной кислоты.
Обитатели третьего слоя – предки эукариот – теперь непосредственно контактировали с новыми обитателями второго слоя – аэробными альфапротеобактериями, которые научились использовать кислород для получения энергии. Метаболизм прото-эукариот и альфапротеобактерий стал взаимодополнительным, что создавало предпосылки для симбиоза. Да и само расположение альфапротеобактерий в сообществе (между верхним, выделяющим кислород, и нижним слоем) предопределяло их роль как «защитников» предков эукариот от избытков кислорода.
Вероятно, прото-эукариоты заглатывали и приобретали в качестве эндосимбионтов многих разных бактерий. Экспериментирование такого рода и сейчас продолжается у одноклеточных эукариот, обладающих огромным разнообразием внутриклеточных симбионтов. Из этих экспериментов союз с аэробными альфапротеобактериями оказался наиболее удачным.
Источник: studopedia.ru
Проблема распространенности жизни во Вселенной
Вопрос о распространенности жизни во Вселенной не решен современной наукой. Постулируя, что в условиях, сходных с теми, которые имелись на молодой Земле, развитие живого вполне вероятно, можно прийти к выводу о том, что в бесконечной Вселенной должны встречаться формы жизни, сходные с земными. На этой принципиальной позиции стоят многие ученые. Тем самым подхватывается мысль Джордано Бруно о множественности обитаемых миров.
Во-первых, в метагалактике есть огромное число звезд, похожих на наше Солнце, следовательно, планетные системы могут существовать не только у Солнца. Более того, исследования показали, что некоторые звезды определенных спектральных классов вращаются медленно вокруг своей оси, что может быть вызвано наличием вокруг этих звезд планетных систем. Во-вторых, молекулярные соединения, необходимые для начальной стадии эволюции неживой природы, достаточно распространены во Вселенной и открыты даже в межзвездной среде. При соответствующих условиях жизнь могла возникнуть на планетах у других звезд по типу эволюционного развития жизни на Земле. В-третьих, нельзя исключать возможность существования небелковых форм жизни, принципиально отличных от тех, которые распространены на Земле.
С другой стороны, многие ученые считают, что даже примитивная жизнь представляетсобой настолько сложную структурно и функционально систему, что даже при наличии на какой-либо планете всех необходимых для ее возникновения условий, вероятность ее спонтанного зарождения крайне низка. Если эти соображения справедливы, то жизнь должнабыть крайне редким и возможно, в пределах наблюдаемой Вселенной, уникальным явлением.
Исходя из данных астрономии, можно однозначно заключить, что в Солнечной системе и других ближайших к нам звездных системах условий для образования цивилизаций не существует. Но не исключается существование примитивных форм жизни. Так, группа американских ученых на основе анализа структуры так называемого “марсианского метеорита” считает, что ими обнаружены свидетельства примитивной одноклеточной жизни, существовавшей на Марсе в далеком прошлом. Ввиду скудности подобного материала сейчас нельзя сделать однозначных выводов по данной проблеме. Возможно, в этом помогут будущие марсианские экспедиции.
Источник: StudFiles.net
Прокариоты
Прокариоты — организмы, состоящие из клеток, которые не имеют клеточного ядра или любых мембранных органелл. Это означает, что генетический материал ДНК у прокариот не связан в ядре. Кроме того, ДНК прокариот менее структурирована, чем у эукариот. В прокариотах ДНК одноконтурная. ДНК эукариот организована в хромосомы. Большинство прокариот состоят только из одной клетки (одноклеточные), но есть несколько и многоклеточных. Ученые разделяют прокариот на две группы: бактерии и археи.
Типичная клетка прокариота включает:
- клеточную стенку;
- плазматическую (клеточную) мембрану;
- цитоплазму;
- рибосомы;
- жгутики и пили;
- нуклеоид;
- плазмиды;
Эукариоты
Эукариоты — живые организмы, клетки которых содержат ядро и мембранные органеллы. Генетический материал у эукариот находится в ядре, а ДНК организована в хромосомы. Эукариотические организмы могут быть одноклеточными и многоклеточными. Все животные являются эукариотами. Также эукариоты включают растения, грибы и простейших.
Типичная клетка эукариота включает:
- плазматическую (клеточную) мембрану;
- ядрышко;
- ядро;
- хромосомы;
- рибосомы;
- эндоплазматический ретикулум;
- аппарат (комплекс) Гольджи;
- цитоскелет;
- цитоплазму;
- лизосомы;
- центриоль;
- митохондрии.
Понравилась статья? Поделись с друзьями:
Источник: NatWorld.info
|
ЗАЧЕМ? Появление эукариотической клетки является вторым по значимости (после зарождения самой жизни) событием биологической эволюции. Важнейшее отличие эукариотических организмов от прокариотических состоит в более совершенной системе регуляции генома (именно в этом смысл появления клеточного ядра: область активного метаболизма – цитоплазма – отделилась от области хранения, считывания, репликации генетической информации и, главное, регуляции транскрипции и посттранскрипционных модификаций РНК). Благодаря этому резко возросла приспособляемость одноклеточных организмов, их способность адаптироваться к меняющимся условиям БЕЗ внесения наследственных изменений в геном, т.е. оставаясь «самими собой». Рост приспособляемости, устойчивости живых систем – основной закон биологической эволюции; в Фанерозое, например, он проявляется в закономерном и неуклонном росте средней продолжительности существования родов (см. об этом наш обзор и статью А.Маркова). Именно благодаря возможности адаптироваться, т.е. изменяться в зависимости от внешних условий, эукариоты смогли стать многоклеточными: ведь в многоклеточном организме клетки с одним и тем же геномом, в зависимости от условий, образуют совершенно разные как по морфологии, так и по функции ткани. |
Фотоальбом «Прокариоты»
Фотоальбом «Одноклеточные эукариоты»
Марков А.В. Ядро земное и ядро клеточное: что между ними общего? (популярная статья)
В.В.Малахов. Основные этапы эволюции эукариотных организмов. 2003.
М. А. Федонкин. Сужение геохимического базиса жизни и эвкариотизация биосферы: причинная связь. 2003.
С. В. Шестаков. О ранних этапах биологической эволюции с позиции геномики. 2003.
Марков А.В. Проблема происхождения эукариот
А.В.Марков, А.М.Куликов. Происхождение эвкариот: выводы из анализа белковых гомологий в трех надцарствах живой природы
О происхождении эукариот. Из популярной книги А.Ю.Журавлева.
Г.А.Заварзин. Эволюция микробных сообществ.
Н.А.Колчанов. Эволюция регуляторных генетических систем.
Рассуждения А.С.Раутиана (из комментариев к кн. Тейяра де Шардена «Феномен человека») о разделении организма на герму (часть, в которой заключена наследственная информация) и сому (все остальное тело) помогают понять значение выделения клеточного ядра: «Разделение организма на сому и герму обусловлено прежде всего тем, что задачи хранения наследственной информации и функционирования предъявляют к своей материальной основе противоположные требования. Любая динамика понижает устойчивость гермы и содержащейся в ней наследственной информации (книга лучше всего сохраняется, если ее не читают). Все живые системы принципиально динамичны. Находясь вдали от состояния термодинамического равновесия, они поддерживают свои существенные параметры благодаря постоянному обмену веществм и энергией с внешней средой. Иными словами, для поддержания сомы необходима динамика, а для сохранения гермы с ее наследственной информацией — покой. Компромисс между этими противоположными требованиями достигается путем пространственного разделения сомы и гермы внутри организма»
А.Ю.Розанов, М.А.Федонкин. Проблема первичного биотопа эвкариот. 1994.
Симбиоз. «Блочный» принцип сборки живых систем. См. об этом в нашем обзоре «Проблема эволюционных новообразований».
Тут есть о чем подумать и в философском плане. Мы свыклись с мыслью о том, что произошли от обезьяны. Обезьяна — зверь, нам близкий и понятный. Сложнее, но все-таки можно представить, что мы произошли от бактерии. Какой-никакой, а все же организм, клетка, что-то конкретное и осязаемое. Но нам становится жутковато, когда мы пытаемся осознать следующую истину в ее неприкрытой бесчеловечности. Наш предок — сообщество бактерий. Мы произошли от кишащей массы разных мельчайших тварей, постепенно сливавшихся в единый организм…
|
|
КОГДА И ПОЧЕМУ? Этот крупнейший ароморфоз произошел, по-видимому, не позднее, чем 2,6 – 2,7 млрд. лет назад, на рубеже Архея и Протерозоя (это определили по биомаркерам – остаткам хим. соединений, свойственных только эукариотам, см. наш обзор «древнейшие следы жизни»). Появление эукариот (точнее, тот момент, когда их присутствие становится заметным в летописи) совпадает по времени с самой крупной за всю историю Земли геофизической перестройкой. Первопричиной этой перестройки, по одной из последних моделей, стало выделение у Земли железного ядра, которое привело к целому комплексу последствий: исключительно сильным конвективным течениям в мантии, образованию «Моногеи» (единого континента), максимуму тектонической активности, смене тектоники тонких базальтовых пластин тектоникой литосферных плит, резкому снижению CO2 в атмосфере и резкому похолоданию (кислород в атмосфере стал накапливаться гораздо позже). Такие катастрофические события могли способствовать развитию эукариот двумя способами. Во-первых, они не могли не привести к разрушению, хотя бы частичному, сложившихся ранее прокариотных сообществ, в частности, цианобактериальных «матов». В ходе кризиса и после стали складываться новые микробные сообщества, уже не чисто прокариотные, а смешанные – прокариотно-эукариотные. Такие сообщества были более устойчивыми. Таким образом, возможно, величайший в истории Земли кризис «помог» эукариотам занять прочное положение в биосфере точно так же, как «массовое вымирание» на рубеже Мезозоя и Кайнозоя помогло млекопитающим и птицам занять множество ниш, которые раньше были заняты рептилиями (пока Мезозойские динозавровые сообщества не были разрушены кризисом, млекопитающие и птицы были вынуждены оставаться второстепенными, подчиненными группами). Во-вторых, очевидно, что в эпоху чрезвычайно резких (катастрофических) колебаний внешних условий более приспособляемые формы должны были получить огромное адаптивное преимущество, должен был идти «отбор на приспособляемость». |
|
|
КАК? Общепризнано, что эукариоты появились в результате симбиоза нескольких разновидностей прокариот (бактерий). По-видимому, митохондрии произошли от альфа-протеобактерий (аэробных эубактерий), пластиды – от цианобактерий, а основная клетка – цитоплазма – от какой-то архебактерии. Пока нет общепринятой теории возникновения ядра, цитоскелета, жгутиков. Приведенная ниже подборка рефератов показывает, как много различных гипотез и моделей сейчас обсуждается. Очевидно, имеющиеся фактические данные пока недостаточны для того, чтобы отдать предпочтение какой-то одной из гипотез или выработать новую, которая устроила бы большинство ученых. Моя точка зрения, основанная на анализе белковых доменов архей, бактерий и эукариот, изложена здесь . Согласно предлагаемой модели, ядерно-цитоплазматический компонент будущих эукариот (перед приобретением митохондриальных симбионтов) представлял собой химерный организм, возникший в результате активного поглощения архебактерией чужеродного (в основном эубактериального) наследственного материала из внешней среды. Возможно, главным стимулом для возникновения такой стратегии у архебактерии стал кризис, вызванный переходом цианобактерий к кислородному фотосинтезу. |
Источник: www.evolbiol.ru