Що таке органела

Зміст:
Що таке органели клітини
Органели клітини – це спеціалізовані структури власне клітини, що відповідають за різні важливі і життєво необхідні функції. Чому ж все-таки «органели»? Просто тут ці компоненти клітини зіставляються з органами багатоклітинного організму.
Які органели входять до складу клітини
Також деколи під органелами розуміється виключно лише постійні структури клітини, які знаходяться в її цитоплазмі. З цієї ж причини ядро клітини та її ядерце не називають органелами, так само як і не є органелами клітинна мембрана, вії та джгутики. А ось до органел, що входять до складу клітини відносяться: хромосоми, мітохондрії, комплекс Гольджі, ендоплазматична мережа, рибосоми, мікротрубочки, мікрофіламенти, лізосоми. По суті це і є основні органели клітини.
Якщо мова йде про тварині клітини, то в число їх органел також входять центріолі та мікрофібрили. А ось в число органоїдів рослинної клітини ще й входять тільки властиві рослинам пластиди. В цілому склад органоїдів в клітинах може істотно відрізнятися в залежності від виду самої клітини.
Малюнок будови клітини, включаючи її органели.
Двомембрані органели клітини
Також в біології існує таке явище як двомембрані органели клітини, до них відносяться мітохондрії і пластиди. Нижче ми опишемо властиві їм функції, втім як і всіх інших основних органел.
Функції органел клітини
А тепер коротко опишемо основні функції органел тваринної клітини. Отже:
- Плазматична мембрана – тонка плівка навколо клітини, що складається з ліпідів та білків. Дуже важливий органоїд, який забезпечує транспортування в клітину води, мінеральних та органічних речовин, видаляє шкідливі продукти життєдіяльності і захищає клітину.
- Цитоплазма – внутрішнє напіврідке середовище клітини. Забезпечує зв’язок між ядром і органелами.
- Ендоплазматична мережа – вона ж мережа каналів в цитоплазмі. Бере активну участь в синтезі білків, вуглеводів та ліпідів, займається транспортуванням корисних речовин.
- Мітохондрії – органели, в яких окислюються органічні речовини і синтезуються молекули АТФ за участю ферментів. По суті мітохондрії це органоїд клітини, що синтезує енергію.
- Пластида (хлоропласти, лейкопласти, хромопласти) – як ми згадували вище, зустрічаються виключно у рослинних клітин, в цілому їх наявність є головною особливістю рослинного організму. Відіграють дуже важливу функцію, наприклад, хлоропласти, що містять зелений пігмент хлорофіл, у рослини відповідають за явище фотосинтезу.
- Комплекс Гольджі – система порожнин, відмежованих від цитоплазми мембраною. Здійснюють синтез жирів та вуглеводів на мембрані.
- Лізосоми – тільця, відокремлені від цитоплазми мембраною. Наявні в них особливі ферменти прискорюють реакцію розщеплення складних молекул. Також лізосома є органоїдом, що забезпечує складання білка в клітинах.
- Вакуолі – порожнини в цитоплазмі, заповнені клітинним соком, місце накопичення запасних поживних речовин; вони регулюють вміст води в клітині.
В цілому всі органели є важливими, адже вони регулюють життєдіяльність клітини.
Источник: www.poznavayka.org
Які бувають органели?
Органела — це складна структура. Деякі з них можуть навіть мати свою власну ДНК і РНК. У всіх клітинах присутні мітохондрії, рибосоми, лізосоми, клітинний центр, апарат (комплекс) Гольджі, ендоплазматична мережа (ретикулум). Рослини мають також специфічні клітинні органели: вакуолі і пластиди. Деякі відносять до органоидам також мікротрубочки і мікрофіламенти.
Органела — це і рибосома, і вакуоль, і клітинний центр, і багато інших. Давайте розглянемо докладніше будова і функції органел.
мітохондрії
Ці органели забезпечують клітину енергією — вонивідповідають за клітинне дихання. Вони є і у рослин, і у тварин, і у грибів. Дані клітинні органели мають дві мембрани: зовнішню і внутрішню, між якими є межмембранное простір. Те, що знаходиться всередині оболонок, називається матриксом. У ньому знаходяться різноманітні ферменти — речовини, необхідні для прискорення хімічних реакцій. Внутрішня мембрана володіє складками — кристами. Саме на них і відбувається процес клітинного дихання. Крім того, в матриксі мітохондрій знаходиться мітохондріальна ДНК (мднк) і мРНК, а також рибосоми, практично аналогічні тим, якими володіють прокариотические клітини.
рибосома
Цей органоид відповідає за процес трансляції, приякому з окремих амінокислот синтезується білок. Будова органели рибосоми простіше, ніж мітохондрії, — вона не володіє мембранами. Даний органоид складається з двох частин (субодиниць) — малої і великої. Коли рибосома не діє, вони знаходяться окремо, а коли вона починає синтезувати білок — об'єднуються. Також збиратися разом можуть і кілька рибосом, якщо полипептидная ланцюжок, синтезируемая ними, дуже довга. Така структура називається "полірібосомамі".
лізосоми
Функції органел цього виду зводяться доздійсненню клітинного травлення. Лізосоми володіють однією мембраною, усередині якої перебувають ферменти — каталізатори хімічних реакцій. Іноді ці органели не тільки розщеплюють поживні речовини, але і переварюють цілі органели. Таке може відбуватися при тривалій голодування клітини і дозволяє їй жити ще деякий час. Хоча якщо поживні речовини все ще не почнуть надходити, клітина вмирає.
Клітинний центр: будова і функції
Ця органела складається з двох частин — центріолей. Це утворення в формі циліндрів, що складаються з мікротрубочок. Клітинний центр — дуже важливий органоид. Він бере участь в процесі формування веретена поділу. Крім того, він є центром організації мікротрубочок.
апарат Гольджі
Це комплекс дископодібних мембранних мішечків,званих цистернами. Функції цього органоида полягають в сортуванні, запасанні і перетворенні деяких речовин. Синтезуються тут в основному вуглеводи, які входять до складу глікокаліксу.
Будова і функції ендоплазматичної
Це мережа трубочок і кишень, оточених однієїмембраною. Існує два види ЕПР: гладкий і шорсткий. На поверхні останнього розташовані рибосоми. Гладкий і шорсткий ретикулум виконують різні функції. Перший відповідає за синтез гормонів, зберігання і перетворення вуглеводів. Крім того, в ньому формуються зачатки вакуолей — органоїдів, характерних для рослинних клітин. Шорсткий ендоплазматичнийретикулум містить на своїй поверхні рибосоми, які виробляють полипептидную ланцюжок з амінокислот. Далі вона потрапляє в ендоплазматичну мережу, і тут формується певна вторинна, третинна і четвертинна структура білка (ланцюжок правильним чином закручується).
вакуолі
Це органели клітини рослин. Вони володіють однією мембраною. У них накопичується клітинний сік. Вакуоль необхідна для підтримки тургору. Також вона бере участь в процесі осмосу. Крім того, існують скоротливі вакуолі. Вони містяться в основному в одноклітинних організмах, що живуть у водоймах, і служать в якості насосів, викачують з клітки зайву рідину.
Пластида: різновиди, будова і функції
Це також органели клітини рослин. Вони бувають трьох видів: лейкопласт, хромопласти і хлоропласти. Перші служать для зберігання запасних поживних речовин, в основному це крохмаль. Хромопласти містять в собі різні пігменти. Завдяки їм пелюстки рослин різнокольорові. Це потрібно організму в першу чергу для того, щоб залучати комах-запилювачів.
Хлоропласти — найважливіші пластиди. Найбільше їх кількість знаходиться в листі і стеблах рослин. Вони відповідають за фотосинтез — ланцюг хімічних реакцій, в процесі яких з неорганічних речовин організм отримує органічні. Ці органели володіють двома мембранами. Матрикс хлоропластів називається "строма". У ній знаходяться пластидних ДНК, РНК, ферменти, а також крохмальні включення. У хлоропластах знаходяться тилакоїди — мембранні освіти у вигляді монети. Усередині їх і відбувається фотосинтез. Тут же міститься і хлорофіл, службовець каталізатором для хімічних реакцій. Тилакоїди хлоропластів об'єднуються в стопки — грани. Також в органелах знаходяться ламелли, які з'єднують між собою окремі тилакоїди і забезпечують зв'язок між ними.
органели руху
Вони характерні в основному для одноклітиннихорганізмів. До них відносяться джгутики і війки. Перші присутні у евглен, трипаносом, хламідомонад. Також джгутики присутні у сперматозоїдів тварин. Вії є у інфузорій та інших одноклітинних.
микротрубочки
Вони забезпечують транспорт речовин, а також постійну форму клітини. Деякі вчені не відносять мікротрубочки до органел.
Источник: uk.garynevillegasm.com
2.5.2.1. Мембранные органоиды (органеллы)
Эндоплазматическая сеть (эндоплазматический ретикулум, цитоплазматическая сеть) — совокупность сообщающихся между собой канальцев, вакуолей и «цистерн», стенка которых образована элементарными биологическими мембранами. Открыта К.Р. Портером в 1945 году. Открытие и описание эндоплазматической сети (ЭПС) обязано внедрению в практику цитологических исследований электронного микроскопа. Мембраны, образующие ЭПС, отличаются от плазмолеммы клетки меньшей толщиной (5-7 нм) и большей концентрацией белков, в первую очередь обладающих ферментативной активностью.
зличают две разновидности ЭПС (рис. 8): шероховатую (гранулярную) и гладкую (агранулярную). Шероховатая ЭПС представлена уплощенными цистернами, на поверхности которых расположены рибосомы и полисомы. Мембраны гранулярной ЭПС содержат белки, способствующие связыванию рибосом и уплощению цистерн. Особенно хорошо развита шероховатая ЭПС в клетках, специализирующихся на белковом синтезе. Гладкую ЭПС формируют переплетающиеся канальцы, трубочки и небольшие пузырьки. Каналы и цистерны ЭПС этих двух разновидностей не разграничены: мембраны одного типа переходят в мембраны другого типа, формируя в области перехода так называемую переходную (транзиторную) ЭПС.
Основными функциями гранулярной ЭПС являются:
1) синтез на прикреплённых рибосомах белков (секретируемых белков, белков клеточных мембран и специфических белков содержимого мембранных органоидов); 2) гидроксилирование, сульфатирование, фосфорилирование и гликозилирование белков; 3) транспорт веществ в пределах цитоплазмы; 4) накопление как синтезируемых, так и транспортируемых веществ; 5) регуляция биохимических реакций, связанная с упорядоченностью локализации в структурах ЭПС веществ, вступающих в реакции, а также их катализаторов — ферментов.
Гладкая ЭПС отличается отсутствием на мембранах белков (рибофоринов), связывающих субъединицы рибосом. Предполагается, что гладкая ЭПС образуется в результате формирования выростов шероховатой ЭПС, мембрана которых утрачивает рибосомы.
Функциями гладкой ЭПС являются: 1) синтез липидов, включая мембранные липиды; 2) синтез углеводов (гликогена и др.); 3) синтез холестерина; 4) обезвреживание токсических веществ эндогенного и экзогенного происхождения; 5) накопление ионов Са2+; 6) восстановление кариолеммы в телофазе митоза; 7) транспорт веществ; 8) накопление веществ.
Как правило, гладкая ЭПС развита в клетках слабее, чем шероховатая ЭПС, однако в клетках, вырабатывающих стероиды, триглицериды и холестерин, а также в клетках печени, осуществляющих детоксикацию различных веществ, она развита значительно лучше.
Рис. 9. Комплекс Гольджи:
1 – стопка уплощённых цистерн; 2 – пузырьки; 3 – секреторные пузырьки (вакуоли)
Переходная (транзиторная) ЭПС — это участок перехода гранулярной ЭПС в агранулярную ЭПС, который располагается у формирующейся поверхности комплекса Гольджи. Трубочки и канальцы переходной ЭПС распадаются на фрагменты, из которых образуются пузырьки, транспортирующие материал из ЭПС в комплекс Гольджи.
Пластинчатый комплекс (комплекс Гольджи, аппарат Гольджи) — органоид клетки, участвующий в окончательном формировании продуктов её жизнедеятельности (секретов, коллагена, гликогена, липидов и других продуктов), а также в синтезе гликопротеидов. Органоид назван по имени описавшего его в 1898 году итальянского гистолога К. Гольджи. Образован тремя составляющими (рис. 9): 1) стопкой уплощённых цистерн (мешочков); 2) пузырьками; 3) секреторными пузырьками (вакуолями). Зона скопления этих элементов получила название диктиосомы. Таких зон в клетке может быть несколько (иногда несколько десятков и даже сотен). Комплекс Гольджи располагается около ядра клетки, часто вблизи центриолей, реже рассеян по всей цитоплазме. В секреторных клетках он располагается в апикальной части клетки, через которую осуществляется выделение секрета путём экзоцитоза. От 3-х до 30-ти цистерн в виде изогнутых дисков диаметром 0,5-5 мкм образуют стопку. Смежные цистерны разделены пространствами в 15-30 нм. Отдельные группы цистерн в пределах диктиосомы отличаются особым составом ферментов, определяющих характер биохимических реакций, в частности процессинга белка и др.
Второй составляющий элемент диктиосомы — пузырьки представляют собой сферические образования диаметром 40-80 нм, умеренно плотное содержимое которых окружено мембраной. Пузырьки формируются путём отщепления от цистерн.
Третий элемент диктиосомы — секреторные пузырьки (вакуоли) представляют собой относительно крупные (0,1-1,0 мкм) сферические мембранные образования, содержащие секрет умеренной плотности, претерпевающий конденсацию и уплотнение (вакуоли конденсации).
Комплекс Гольджи отчётливо поляризован по вертикали. В нём выделяют две поверхности (два полюса):
1) цис-поверхность, или незрелую поверхность, которая имеет выпуклую форму, обращена к эндоплазматической сети (ядру) и связана с отделяющимися от неё мелкими транспортными пузырьками;
2) транс-поверхность, или поверхность, обращённую к плазмолемме вогнутой формы (рис. 8), со стороны которой от цистерн комплекса Гольджи отделяются вакуоли (секреторные гранулы).
Основными функциями комплекса Гольджи являются: 1) синтез гликопротеинов и полисахаридов; 2) модификация первичного секрета, его конденсация и упаковка в мембранные пузырьки (формирование секреторных гранул); 3) процессинг молекул (фосфорилирование, сульфатирование, ацилирование и т.п.); 4) накопление секретируемых клеткой веществ; 5) образование лизосом; 6) сортировка синтезированных клеткой белков у транс-поверхности перед их окончательным транспортом (производится посредством рецепторных белков, распознающих сигнальные участки макромолекул и направляющих их в различные пузырьки); 7) транспорт веществ: из транспортных пузырьков вещества проникают в стопку цистерн комплекса Гольджи с цис-поверхности, а выходят из неё в виде вакуолей с транс-поверхности. Механизм транспорта объясняют две модели: а) модель перемещения пузырьков, отпочковывающихся от предшествующей цистерны и сливающихся с последующей цистерной последовательно в направлении от цис-поверхности к транс-поверхности; б) модель перемещения цистерн, основанная на представлении о непрерывном новообразовании цистерн за счёт слияния пузырьков на цис-поверхности и последующем распаде на вакуоли цистерн, смещающихся к транс-поверхности.
Указанные выше основные функции позволяют констатировать, что пластинчатый комплекс — важнейший органоид клетки эукариот, обеспечивающий организацию и интеграцию внутриклеточного метаболизма. В этом органоиде протекают заключительные этапы формирования, созревания, сортировки и упаковки всех секретируемых клеткой продуктов, ферментов лизосом, а также белков и гликопротеинов поверхностного аппарата клетки и др. веществ.
Органоиды внутриклеточного переваривания. Лизосомы — это мелкие ограниченные элементарной мембраной пузырьки, содержащие гидролитические ферменты. Мембрана лизосом толщиной около 6 нм осуществляет пассивную компартментализацию, временно отделяя гидролитические ферменты (более 30 разновидностей) от гиалоплазмы. В неповреждённом состоянии мембрана устойчива к действию гидролитических ферментов и препятствует их утечке в гиалоплазму. В стабилизации мембраны важная роль принадлежит кортикостероидным гормонам. Повреждение мембран лизосом ведёт к самоперевариванию клетки гидролитическими ферментами.
Мембрана лизосом содержит АТФ-зависимый протонный насос, обеспечивающий закисление среды внутри лизосом. Последняя способствует активизации ферментов лизосом — кислых гидролаз. Наряду с этим мембрана лизосом содержит рецепторы, обусловливающие связывание лизосом с транспортными пузырьками и фагосомами. Мембрана обеспечивает также диффузию веществ из лизосом в гиалоплазму. Связывание части молекул гидролаз с мембраной лизосом ведёт к их инактивации.
Выделяют несколько разновидностей лизосом: первичные лизосомы (гидролазные пузырьки), вторичные лизосомы (фаголизосомы, или пищеварительные вакуоли), эндосомы, фагосомы, аутофаголизосомы, остаточные тельца (рис. 8).
Эндосомами называют мембранные пузырьки, переносящие макромолекулы от поверхности клетки в лизосомы путём эндоцитоза. В процессе переноса содержимое эндосом может не изменяться или претерпевать частичное расщепление. В последнем случае в эндосомы проникают гидролазы или эндосомы непосредственно сливаются с гидролазными пузырьками, вследствие чего среда постепенно закисляется. Эндосомы разделяют на две группы: ранние (периферические) и поздние (перинуклеарные) эндосомы.
Ранние (периферические) эндосомы формируются на ранних этапах эндоцитоза после отделения пузырьков с захваченным содержимым от плазмолеммы. Они располагаются в периферических слоях цитоплазмы и характеризуются нейтральной или слабощелочной средой. В них происходит отщепление лигандов от рецепторов, сортировка лигандов и, возможно, возвращение рецепторов в специальных пузырьках в плазмолемму. Наряду с этим в ранних эндосомах может происходить расщепление ком-
Рис. 10 (А). Схема образования лизосом и их участия во внутриклеточном пищеварении. (Б) Электронная микрофотография среза вторичных лизосом (обозначены стрелками):
1 – образование из гранулярной эндоплазматической сети мелких пузырьков с ферментами; 2 – перенос ферментов в аппарат Гольджи; 3 – образование первичных лизосом; 4 – выделение и использование (5) гидролаз при внеклеточном ращеплении; 6 — фагосомы; 7 – слияние первичных лизосом с фагосомами; 8, 9 – образование вторичных лизосом (фаголизосом); 10 – экскреция остаточных телец; 11 – слияние первичных лизосом с разрушающимися структурами клетки; 12 – аутофаголизосома
плексов «рецептор-гормон», «антиген-антитело», ограниченное расщепление антигенов, инактивация отдельных молекул. В условиях закисления (рН=6,0) среды в ранних эндосомах может происходить частичное расщепление макромолекул. Постепенно, перемещаясь вглубь цитоплазмы, ранние эндосомы превращаются в поздние (перинуклеарные) эндосомы, располагающиеся в глубоких слоях цитоплазмы, окружающих ядро. Они достигают 0,6-0,8 мкм в диаметре и отличаются от ранних эндосом более кислым (рН=5,5) содержимым и более высоким уровнем ферментативного переваривания содержимого.
Фагосомы (гетерофагосомы) — мембранные пузырьки, которые содержат захваченный клеткой извне материал, подлежащий внутриклеточному перевариванию.
Первичные лизосомы (гидролазные пузырьки) — пузырьки диаметром 0,2-0,5 мкм, содержащие неактивные ферменты (рис.10). Их перемещение в цитоплазме контролируется микротрубочками. Гидролазные пузырьки осуществляют транспорт гидролитических ферментов из пластинчатого комплекса к органоидам эндоцитозного пути (фагосомам, эндосомам и т.п.).
Вторичные лизосомы (фаголизосомы, пищеварительные вакуоли) — пузырьки, в которых активно осуществляется внутриклеточное переваривание посредством гидролаз при рН≤5. Их диаметр достигает 0,5-2 мкм. Вторичные лизосомы (фаголизосомы и аутофаголизосомы) формируются путём слияния фагосомы с эндосомой или первичной лизосомой (фаголизосомы) либо путём слияния аутофагосомы (мембранного пузырька, содержащего собственные компоненты клетки) с первичной лизосомой (рис. 10) или поздней эндосомой (аутофаголизосомы). Аутофагия обеспечивает переваривание участков цитоплазмы, митохондрий, рибосом, фрагментов мембран и т.п. Убыль последних в клетке компенсируется их новообразованием, что ведёт к обновлению («омоложению») клеточных структур. Так, в нервных клетках человека, функционирующих многие десятилетия, большинство органоидов обновляется в течение 1 месяца.
Разновидность лизосом, содержащих непереваренные вещества (структуры), названа остаточными тельцами. Последние могут длительно находиться в цитоплазме или выделять своё содержимое путём экзоцитоза за пределы клетки (рис. 10). Распространённым видом остаточных телец в организме животных являются липофусциновые гранулы, представляющие собой мембранные пузырьки (0,3-3 мкм), содержащие труднорастворимый коричневый пигмент липофусцин.
Пероксисомы представляют собой мембранные пузырьки диаметром до 1,5 мкм, матрикс которых содержит около 15 ферментов (рис. 8). Среди последних наиболее важны каталаза, на которую приходится до 40% общего белка органоида, а также пероксидаза, оксидаза аминокислот и др. Пероксисомы образуются в эндоплазматическом ретикулуме и обновляются каждые 5-6 дней. Наряду с митохондриями, пероксисомы являются важным центром утилизации кислорода в клетке. В частности, под воздействием каталазы распадается перекись водорода (Н2О2), образующаяся в ходе окисления аминокислот, углеводов и др. веществ клетки. Таким образом, пероксисомы защищают клетку от повреждающего эффекта перекиси водорода.
Органоиды энергетического обмена. Митохондрии описаны впервые Р. Келликером в 1850 году в мышцах насекомых под названием саркосом. Позднее они изучались и описывались Р. Альтманом в 1894 году как «биопласты», а в 1897 году К. Бенда назвал их митохондриями. Митохондрии представляют собой мембранные органоиды, обеспечивающие клетку (организм) энергией. Источником запасаемой в виде фосфатных связей АТФ энергии являются процессы окисления. Наряду с этим митохондрии участвуют в биосинтезе стероидов и нуклеиновых кислот, а также в окислении жирных кислот.
М
Рис. 11. Схема строения митохондрии:
1 – наружная мембрана; 2 – внутренняя мембрана; 3 – кристы; 4 – матрикс
итохондрии имеют эллиптическую, сферическую, палочковидную, нитевидную и др. формы, которые могут изменяться в течение определенного времени. Их размеры составляют 0,2-2 мкм в ширину и 2-10 мкм в длину. Количество митохондрий в различных клетках варьирует в широких пределах, достигая в наиболее активных 500-1000. В клетках печени (гепатоцитах) их число составляет около 800, а занимаемый ими объем равен примерно 20% объема цитоплазмы. В цитоплазме митохондрии могут располагаться диффузно, однакообычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл), органелл движения (аксонема спермия). Митохондрии состоят из наружной и внутренней мембран, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны — кристы (рис. 11, 12).
Н
Рис. 12. Электронная фотография митохондрии (поперечный разрез)
аружная мембрана митохондрий сходна с плазмолеммой. Она отличается высокой проницаемостью, обеспечивая проникновение молекул с массой менее 10 килодальтон из цитозоля в межмембранное пространство митохондрий. Наружная мембрана содержит порин и другие транспортные белки, а также рецепторы, распознающие переносимые белки в зонах слипания наружной и внутренней мембран.
Межмембранное пространство митохондрий шириной 10-20 нм содержит небольшое количество ферментов. Его ограничивает изнутри внутренняя мембрана митохондрий, содержащая транспортные белки, ферменты дыхательной цепи и сукцинатдегидрогеназу, а также комплекс АТФ-синтетазы. Внутренняя мембрана характеризуется низкой проницаемостью для мелких ионов. Она формирует складки толщиной 20 нм, которые располагаются чаще всего перпендикулярно продольной оси митохондрий, а в некоторых случаях (мышечные и др. клетки) — продольно. С повышением активности митохондрий количество складок (их общая площадь) возрастает. На кристах находятся оксисомы — грибовидные образования, состоящие из округлой головки диаметром 9 нм и ножки толщиной 3 нм. В области головки происходит синтез АТФ. Процессы окисления и синтеза АТФ в митохондриях разобщены, из-за чего не вся энергия накапливается в АТФ, рассеиваясь частично в виде тепла. Такое разобщение наиболее выражено, например, в бурой жировой ткани, используемой для весеннего «разогрева» находившихся в состоянии «зимней спячки» животных.
Внутренняя камера митохондрии (область между внутренней мембраной и кристами) заполнена матриксом (рис. 11, 12), содержащим ферменты цикла Кребса, ферменты белкового синтеза, ферменты окисления жирных кислот, митохондриальную ДНК, рибосомы и митохондриальные гранулы.
Митохондриальная ДНК представляет собственный генетический аппарат митохондрий. Она имеет вид кольцевой двухцепочечной молекулы, в которой содержится около 37 генов. Митохондриальная ДНК отличается от ядерной ДНК низким содержанием некодирующих последовательностей и отсутствием связей с гистонами. Митохондриальная ДНК кодирует иРНК, тРНК и рРНК, однако обеспечивает синтез только 5-6% митохондриальных белков (ферментов системы транспорта ионов и некоторых ферментов синтеза АТФ). Синтез всех других белков, а также удвоение митохондрий контролируются ядерной ДНК. Большая часть рибосомальных белков митохондрий синтезируется в цитоплазме, а затем транспортируется в митохондрии. Наследование митохондриальной ДНК у многих видов эукариот, включая человека, происходит только по материнской линии: митохондриальная ДНК отца исчезает при гаметогенезе и оплодотворении.
Митохондрии имеют относительно короткий жизненный цикл (около 10 суток). Разрушение их происходит путём аутофагии, а новообразование — путём деления (перешнуровки) предшествующих митохондрий. Последнему предшествует репликация митохондриальной ДНК, которая происходит независимо от репликации ядерной ДНК в любые фазы клеточного цикла.
У прокариот митохондрии отсутствуют, и их функции выполняет клеточная мембрана. Согласно одной из гипотез, митохондрии произошли из аэробных бактерий в результате симбиогенеза. Существует предположение об участии митохондрий в передаче наследственной информации.
Источник: StudFiles.net
Плазмолема
Плазмолема відокремлює клітину від навколишнього середовища й у той же час забезпечує обмін речовин між середовищем і клітиною. Вона — напівпроникна для іонізованих часток, тому на її поверхні створюється різниця потенціалів, що сприяє проходженню прямого і зворотного піноцитозу. Складається вона з надмембранного, мембранного та субмембранного комплексів. Мембранний комплекс представлений типовою елементарною мембраною, яка складається з двох мономолекулярних білкових шарів, розділених бімолекулярним ліпідним прошарком. Надмембранний комплекс утворений гліколіпідами та глікопротеїнами. Вони формують на поверхні плазмолеми глікокалікс. Субмембранний комплекс — цитоскелет утворюють білки тубуліни.
Лізосоми
Пухирці, оточені мембраною, містять до 60 різних гідролізованих ферментів, що діють у кислому середовищі. Вони здатні розщеплювати білки, вуглеводи і нуклеїнові кислоти.
Утворюються лізосоми в пластинчастому комплексі. В міру їхнього старіння розрізняють чотири типи лізосом: 1) первинні лізосоми, чи гранули, що запасають, які ще не беруть участі у розщепленні; 2) вторинні лізосоми, чи травні вакуолі, утворені злиттям первинної лізосоми з фагосомою чи з піноцитозним пухирцем; 3) залишкові тільця; 4) цитолізосоми, тобто лізосоми-санітари, які автолізують клітину, що гине, чи звільняють клітину від структур, що відмирають.
Мітохондрії
Форма цієї органели є дуже різноманітною не тільки у різних клітин, але й у межах однієї клітини, що характеризує фізіологічний стан останньої. Мітохондрії надзвичайно поширені і є силовою станцією клітини, що дає необхідну для життя енергію. Особливо їх багато в м’язовій і нервовій тканинах, які відрізняються високою активністю обміну речовин і багатим кровопостачанням. Мітохондрії мають форму ниток, паличок чи дрібних зерен. Електронна мікроскопія показує, що вони складаються з двох мітохондріальних мембран, вставлених одна в одну, а проміжок між ними заповнений рідким вмістом — мітохондріальним матриксом. Внутрішня мітохондріальна мембрана утворює численні гребені (кристи), чи трубчасті вирости, що вдавлюються в порожнину мітохондрій.
Основна функція мітохондрій — це клітинне дихання, тобто окислення вуглеводів чи жирів, пов’язаних з фосфорилюванням, у результаті якого створюються запаси АТФ макроергічної речовини, що є джерелом енергії, необхідної для клітини. Близько 90% енергії, яку споживає клітина, утворюють мітохондрії, а також вони беруть участь у переносі іонів і води.
Клітинний центр чи центросома
Ця органела притаманна тваринним і багатьом рослинним клітинам. Основна і постійна її частина — центріолі, які побудовані з дев’яти триплетів мікротрубочок та функціонально зв’язані з рухом клітини, що особливо видно на прикладі розподілу клітини. Навколо центріолей розташована центросфера — скупчення безструктурної цитоплазми, часто оточене цистернами пластинчастого комплексу. Іноді навколо центріолей утворюються трубчасті нитки, що розходяться радіально в усі сторони — промениста сфера (стан клітини при мітотичному розподілі).
Цитоплазматична (ендоплазматична) сітка
Представлена ця органела трубками і пухирцями, не зв’язаними між собою. Вона дуже мінлива, що пов’язано з регулюванням площі активних поверхонь, на яких відбуваються важливі біологічні процеси. Цитоплазматична сітка — транспортна система клітини, на мембранах якої адсорбуються різні речовини, що вступають у хімічну взаємодію.
Розрізняють два типи цитоплазматичної сітки: гранулярну й агранулярну. Перша на своїй поверхні містить особливі гранули РНК — рибосоми, а інша їх не має. Гранулярна сітка бере участь у синтезі білків, агранулярна — у синтезі вуглеводів.
Рибосоми — складова частина гранулярної цитоплазматичної сітки, мають неправильно округлу форму і складаються з двох нерівних частин — субодиниць. Попарне з’єднання їх залежить від наявності в цитоплазмі достатньої кількості іонів магнію. Рибосоми звичайно розташовуються групами — полісомами чи полірибосомами. Їхню функцію пов’язують із синтезом білків клітини.
Пластинчастий комплекс
Внутрішньоклітинний сітчастий апарат, особливо виражений у залозистих клітинах і взагалі в клітинах з інтенсивним синтезом речовин (нервові, яйцеві клітини). Розташований він навколо ядра чи між ядром і полюсом клітини, що виділяє секрет.
Пластинчастий комплекс складається з цистерн, мікропухирців і вакуолей. За допомогою мікропухирців, вкритих мембраною, здійснюється зв’язок цієї органели з цитоплазматичною сіткою. Пластинчастий комплекс — активна органела, тісно зв’язана із синтезом речовин, особливо в тих випадках, коли вони синтезуються у великих кількостях.
Препарат 5. Пластинчастий комплекс в нервових клітинах спинального ганглія (рис. 7).
У підручниках існує декілька назв цієї органели: зона Гольджі, апарат Гольджі, сітчастий апарат та ін. Препарат являє собою гістологічний зріз через спинальний ганглій, оброблений складним методом імпрегнації азотнокислим сріблом.
При малому збільшенні потрібно відшукати нервові клітини між нервовими волокнами. Клітини найчастіше круглої, овальної або ж багатогранної форми, досить великих розмірів.
Потрібно в центрі поля зору поставити декілька клітин.
Вивчаючи різні клітини при великому збільшенні, можна впевнитись у наявності в цитоплазмі дуже ніжної сітки навколо ядра. Вона має вигляд малих паличок, ком та ін. Це і є пластинчастий комплекс. У різних клітинах вигляд цієї органели не зовсім однаковий, що залежить від функціонального стану клітини.
Замалювати при великому збільшенні — об. 40, ок. 15 — декілька нервових клітин з пластинчастим комплексом. У деяких клітинах каріоплазма може бути відсутньою тому, що зріз пройшов нижче або вище ядра (рис. 7).
Цитоплазматичний ретикулум (ендоплазматична сітка)
Препарат 6. Тигроїдна речовина в нервових клітинах спинального ганглія (рис. 8).
Препарат являє собою зріз спинального ганглія, зафарбований толуїдиновою синькою.
При малому збільшенні добре спостерігаються нервові клітини, які мають різні розміри та круглу, овальну або ж полігональну форми. Каріоплазма клітини має вигляд світлого пухирця, а цитоплазма зафарбована у світлий колір.
При великому збільшенні в ядрі видно кругле темне ядерце, оточене світлою каріоплазмою. Цитоплазма клітин заповнена невеличкими брилками, зафарбованими в синій колір. Ці брилки і є тигроїдною речовиною або субстанцією (речовиною) Ніссля. Як виявилось при субмікроскопічному дослідженні, тигроїдна речовина — це скупчення рибосом на мембранах ендоплазматичної сітки (тобто гранулярний ендоплазматичний ретикулум). Звичайно, при світловій мікроскопії цієї сітки не видно.
Замалювати при великому збільшенні — об. 40, ок. 15 — декілька нервових клітин з тигроїдною речовиною в цитоплазмі (рис. 8).
Рис. 7.
Пластинчастий комплекс в нервових клітинах спинального ганглія
.
Позначення: 1. Оболонка клітини. 2. Цитоплазма. 3. Пластинчастий комплекс. 4. Каріоплазма
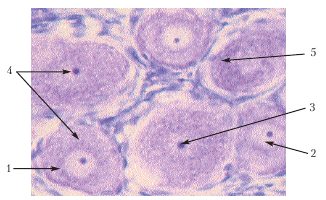
Рис. 8.
Цитоплазматичний ретикулум у нервових клітинах спинального ганглія
.
Позначення: 1. Нервова клітина. 2. Каріоплазма. 3. Ядерце. 4.Цитоплазма. 5. Тигроїдна речовина
Источник: med-books.info