Трофические связи это
Трофические связи возникают, когда один вид питается другим – либо живыми особями, либо их мертвыми остатками, либо продуктами жизнедеятельности. И стрекозы, ловящие на лету других насекомых, и жуки-навозники, питающиеся пометом крупных копытных, и пчелы, собирающие нектар растений, вступают в прямую трофическую связь с видами, предоставляющими им пищу. В случае конкуренции двух видов из-за объектов питания между ними возникает косвенная трофическая связь, так как деятельность одного отражается на снабжении кормом другого. Любое воздействие одного вида на поедаемость другого или доступность для него пищи следует расценивать как косвенную трофическую связь между ними. Например, гусеницы бабочек-монашенок, объедая хвою сосен, облегчают короедам доступ к ослабленным деревьям.
Трофические связи являются главными в сообществах. Именно они объединяют живущие вместе виды, поскольку каждый из них может обитать лишь там, где имеются необходимые ему пищевые ресурсы. Любой вид не только приспособлен к определенным источникам питания, но и сам служит пищевым ресурсом для других.
щевые взаимосвязи создают в природе трофическую сеть, распространяющуюся в конечном счете на все виды в биосфере. Образ этой трофической сети можно воссоздать, поставив в центр любой вид и соединив его стрелками со всеми другими, которые находятся с ним в прямых или косвенных пищевых отношениях, и затем продолжать эту процедуру для каждого вовлеченного в схему вида. В результате будет охвачена вся живая природа, от китов до бактерий. Любой биоценоз пронизан пищевыми связями и представляет собой более или менее локализованный в пространстве участок общей трофической сети, связывающей все живое на Земле.
ТРОФИЧЕСКАЯ ЦЕПЬ, пищевая цепь, цепь питания, взаимоотношения между организмами, через которые в экосистеме происходит трансформация вещества и энергии; группы особей (бактерии, грибы, растения и животные), связанные друг с другом отношением пища — потребитель.
В трофической цепи при переносе потенциальной энергии от звена к звену большая её часть (до 80-90%) теряется в виде теплоты. Поэтому число звеньев (видов) в трофической цепи обычно не превышает 4-5 и, очевидно, чем длиннее трофическая цепь, тем меньше продукция её последнего звена по отношению к продукции начального. В состав пищи каждого вида входит обычно не один, а несколько или много видов, каждый из которых в свою очередь может служить пищей нескольким видам. Поэтому трофические взаимоотношения видов в природе точнее передаются термином трофическая сеть (или паутина). Однако представление о трофической цепи сохраняет своё значение, когда оказывается возможным разнести всех членов сообщества по отдельным звеньям цепи — трофическим уровням.
Существует 2 основных типа трофических цепей — пастбищные и детритные.
В пастбищной трофической цепи (цепь выедания) основу составляют автотрофные организмы, затем идут потребляющие их растительноядные животные (например, зоопланктон, питающийся фитопланктоном), потом хищники (консументы) 1-го порядка (например, рыбы, потребляющие зоопланктон), хищники 2-го порядка (например, судак, питающийся другими рыбами). Особенно длинны трофические цепи в океане, где многие виды (например, тунцы) занимают место консументов 4-го порядка.
В детритных трофических цепях (цепи разложения), наиболее распространенных в лесах, большая часть продукции растений не потребляется непосредственно растительноядными животными, а отмирает, подвергаясь затем разложению сапротрофными организмами и минерализации. Таким образом, детритные трофические цепи начинаются от детрита, идут к микроорганизмам, которые им питаются, а затем к детритофагам и к их потребителям — хищникам. В водных экосистемах (особенно в эвтрофных водоемах и на больших глубинах океана) значительная часть продукции растений и животных также поступает в детритные трофические цепи.
Топические связиТопические связи характеризуют любое, физическое или химическое, изменение условий обитания одного вида в результате жизнедеятельности другого. Эти связи крайне разнообразны. Они заключаются в создании одним видом среды для другого (например, внутренний паразитизм или норовый комменсализм), в формировании субстрата, на котором поселяются или, наоборот, избегают селиться представители других видов, во влиянии на движение воды, воздуха, изменение температуры, освещенности окружающего пространства, в насыщении среды продуктами выделения и т. п. Морские желуди, поселяющиеся на коже китов, личинки мух, обитающие в лепешках коровьего навоза,
На основе топических связей в биоценозе формируются консорции (сочетания) – группы разнородных организмов, поселяющихся на теле или в теле особи какого-либо определенного вида – центрального члена консорции. В большинстве случаев члены одной консорции связаны также разнообразными трофическими отношениями. Консорции формируются фактически вокруг представителей любого вида, обладающего средообразующим воздействием на других. По свидетельству В. Н. Беклемишева, «сосна с ее микоризными грибками, с эпифитными мхами и лишайниками на стволе и ветвях, со всем множеством населяющих ее членистоногих – это сложнейший консорций, это – целый мир. Полевка с ее эктопаразитами, с гельминтами, с простейшими, бактериями, населяющими ее внутренние органы, опять есть целый консорций».
Отдельные консорции могут быть разной степени сложности. Наибольшим числом консортивных связей отличаются те растения, которым принадлежит основная роль в создании внутренней среды биоценоза. Так как каждый член крупной консорции может быть, в свою очередь, центром более мелкого объединения, можно выделить консорции первого, второго и даже третьего порядка. Таким образом, биоценоз – это система связанных между собой консорций, возникающих на основе теснейших топических и трофических отношений между видами. Консортивные связи, в основе которых лежат топические отношения, формируют своего рода блочную структуру биоценоза.
Топические и трофические связи имеют наибольшее значение в биоценозе, составляют основу его существования. Именно эти типы отношений удерживают друг возле друга организмы разных видов, объединяя их в достаточно стабильные сообщества разных масштабов.
Форические связиФорические связи – это участие одного вида в распространении другого. В роли транспортировщиков выступают животные. Перенос животными семян, спор, пыльцы растений называют зоохорией, перенос других, более мелких животных – форезией (от лат. форас– наружу, вон). Перенос осуществляется обычно с помощью специальных и разнообразных приспособлений. Животные могут захватывать семена растений двумя способами: пассивным и активным. Пассивный захват происходит при случайном соприкосновении тела животного с растением, семена или соплодия которого обладают специальными зацепками, крючками, выростами (череда, лопух). Распространителями их обычно служат млекопитающие, которые на шерсти переносят такие плоды иногда на довольно значительные расстояния. Активный способ захвата – поедание плодов и ягод. Не поддающиеся перевариванию семена животные выделяют вместе с пометом. В переносе грибных спор большую роль играют насекомые. По-видимому, плодовые тела грибов возникли как образования, привлекающие насекомых-расселителей.
Форезия животных распространена преимущественно среди мелких членистоногих, особенно у разнообразных групп клещей (рис. 89). Она представляет собой один из способов пассивного расселения и свойственна видам, для которых перенос из одного биотопа в другой жизненно необходим для сохранения или процветания. Например, многие летающие насекомые – посетители скоплений быстро разлагающихся растительных остатков (трупов животных, помета копытных, куч гниющих растений и т. п.) несут на себе гамазовых, уроподовых или тироглифоидных клещей, переселяющихся таким образом от одного скопления пищевых материалов к другому. Собственные расселительные возможности не позволяют этим видам преодолевать значительные для них расстояния. Жуки-навозники иногда ползают с поднятыми надкрыльями, которые не в состоянии сложить из-за густо усеявших тело клещей. Посредством форезии на насекомых распространяются некоторые виды нематод. Ноги навозных мух часто имеют вид ламповых щеток из-за обилия прикрепившихся к ним нематодрабдитид. Среди крупных животных форезия почти не встречается.
Фабрические связиФабрические связи – это такой тип биоценотических отношений, в которые вступает вид, использующий для своих сооружений (фабрикаций) продукты выделения, либо мертвые остатки, либо даже живых особей другого вида. Так, птицы употребляют для постройки гнезд ветви деревьев, шерсть млекопитающих, траву, листья, пух и перья других видов птиц и т. п. Личинки ручейников строят домики из кусочков веток, коры или листьев растений, из раковин мелких видов катушек, захватывая даже раковинки с живыми моллюсками. Пчела-мегахила помещает яйца и запасы в стаканчики, сооружаемые из мягких листьев различных кустарников (шиповника, сирени, акации и т. п.).
Из-за сложности межвидовых отношений каждый конкретный вид может преуспевать далеко не везде, где складываются подходящие для него условия физической среды. Различают физиологический и синэкологический оптимумы вида. Физиологический оптимум – это благоприятное для вида сочетание всех абиотических факторов, при котором возможны наиболее быстрые темпы роста и размножения. Синэкологический оптимум – это такое биотическое окружение, при котором вид испытывает наименьшее давление со стороны врагов и конкурентов, что позволяет ему успешно размножаться. Синэкологический и физиологический оптимумы далеко не всегда совпадают. Если в подходящем биотопе экологическая ниша занята более сильным конкурентом или чересчур велико влияние хищников и паразитов, вид в нем не приживается.
имер несовпадения физиологического и синэкологического оптимумов – массовое размножение вредителя зерновых культур, гессенского комарика, после особенно суровых зим, которые, казалось бы, должны неблагоприятно сказываться на численности этого насекомого. В нормальные по условиям годы гессенского комарика сильно истребляют несколько видов его естественных врагов – паразитических перепончатокрылых наездников. В суровые зимы из-за очень слабой морозоустойчивости враги гессенского комарика вымерзают почти полностью. Это дает возможность вредителю быстро восстановить собственную численность, сокращенную морозами, и беспрепятственно размножаться в угрожающем для урожая количестве.
Экологическая нишаПоложение вида, которое он занимает в общей системе биоценоза, комплекс его биоценотических связей и требований к абиотическим факторам среды называют экологической нишей вида.
Концепция экологической ниши оказалась очень плодотворной для понимания законов совместной жизни видов. Над ее развитием работали многие экологи: Дж. Гриннелл, Ч. Элтон, Г. Хатчинсон, Ю. Одум и др.
Понятие «экологическая ниша» следует отличать от понятия «местообитание». В последнем случае подразумевается та часть пространства, которая заселена видом и которая обладает необходимыми абиотическими условиями для его существования. Экологическая ниша вида зависит не только от абиотических условий среды, но и в не меньшей мере от его биоценотического окружения.
рактер занимаемой экологической ниши определяется как экологическими возможностями вида, так и тем, насколько эти возможности могут быть реализованы в конкретных биоценозах. Это характеристика того образа жизни, который вид может вести в данном сообществе. Термин экологическая ниша предложен Джозефом Гриннеллом в 1917 году и изначально был близок к понятию биотопа. Позже Ч. Элтон определил экологическую нишу как положение вида в сообществе. Было отмечено, что экологически близкие и занимающие сходное положение в сообществе виды не могут устойчиво существовать на одной территории. По образному выражению Одума, биотоп — это адрес организма, а ниша — это его профессия. Разные виды могут занимать один биотоп, но различные экологические ниши. И наоборот, виды могут занимать сходные экологические ниши в разных биотопах.
Современное понятие экологической ниши дано в 1957, 1965 годах Джорджем Хатчинсоном. Экологическая ниша по Хатчинсону представляет многомерное пространство, отдельные измерения которого соответствуют факторам, необходимым для нормального существования вида. Таких факторов может быть очень много. Изучение экологических ниш показало, что в большинстве случаев достаточно рассмотрения трех из них: пространственного, пищевого и временного. Сейчас разработаны методы оценки ширины экологической ниши и степени перекрывания экологических ниш разных видов. Для исследований перекрывания ниш часто используют матричный анализ.
Даже у одного вида могут быть различные экологические ниши в разных районах обитания, что зависит от доступности пищи и числа конкурентов и других факторов. Некоторые организмы, онтогенез которых разделен на ряд разных этапов, занимают последовательно различные экологические ниши. Так, головастик травяной лягушки — консумент первого порядка, обитающий в водной экосистеме, а взрослая лягушка – консумент второго порядка или хищник первого, обитает большую часть сезона на суше. Такую же смену экологических ниш мы можем наблюдать у беспозвоночных животных: водные личинки и сухопутные взрослые особи многих насекомых.
Понятие экологической ниши Одумом (1959) было расширено на поведенческую составляющую. Он подчеркивал, что экологическая ниша организма зависит не только от того, где организм живет, но и от того, что он делает. Говоря о поведенческой составляющей экологической ниши, можно привести следующие наглядные примеры. В пустынях Средней Азии живут представители двух семейств ящериц: агамы и ящурки. Эти семейства отличаются по способу охоты и использования территории. Агамы чаще сидят и караулят свою добычу (как кошки), а ящурки активно ищут ее по всей территории (как волки). В результате они легко уживаются на одной и той же территории. Разделению экологической ниши внутри вида способствует сложное территориальное поведение, которым обладают многие наземные позвоночные.
Г. Хатчинсон выдвинул понятия фундаментальной и реализованной экологической ниши.
д фундаментальной понимается весь набор условий, при которых вид может успешно существовать и размножаться. В природных биоценозах, однако, виды осваивают далеко не все пригодные для них ресурсы вследствие, прежде всего, конкурентных отношений. Реализованная экологическая ниша – это положение вида в конкретном сообществе, где его ограничивают сложные биоценотические отношения. Иными словами, фундаментальная экологическая ниша характеризует потенциальные возможности вида, а реализованная – ту их часть, которая может осуществиться в данных условиях, при данной доступности ресурса. Таким образом, реализованная ниша всегда меньше, чем фундаментальная.
В экологии широко обсуждается вопрос о том, сколько экологических ниш может вместить биоценоз и сколько видов какой-либо конкретной группы, имеющих близкие требования к среде, могут ужиться вместе.
Специализация вида по питанию, использованию пространства, времени активности и другим условиям характеризуется как сужение его экологической ниши, обратные процессы – как ее расширение. На расширение или сужение экологической ниши вида в сообществе большое влияние оказывают конкуренты. Правило конкурентного исключения, сформулированное Г. Ф. Гаузе для близких по экологии видов, может быть выражено таким образом, что два вида не уживаются в одной экологической нише.
Эксперименты и наблюдения в природе показывают, что во всех случаях, когда виды не могут избежать конкуренции за основные ресурсы, более слабые конкуренты постепенно вытесняются из сообщества. Однако в биоценозах возникает много возможностей хотя бы частичного разграничения экологических ниш близких по экологии видов.
Выход из конкуренции достигается благодаря расхождению требований к среде, изменению образа жизни, что, другими словами, является разграничением экологических ниш видов. В этом случае они приобретают способность сосуществовать в одном биоценозе. Каждый из живущих вместе видов в отсутствие конкурента способен на более полное использование ресурсов. Это явление легко наблюдать в природе. Так, травянистые растения ельника способны довольствоваться небольшим количеством почвенного азота, которое остается от перехвата его корнями деревьев. Однако если на ограниченной площадке обрубить корни этих елей, условия азотного питания трав улучшаются и они бурно идут в рост, принимая густо-зеленую окраску. Улучшение условий жизни и увеличение численности какого-либо вида в результате удаления из биоценоза другого, близкого по экологическим требованиям, называется конкурентным высвобождением.
Разделение совместно живущими видами экологических ниш с частичным их перекрыванием – один из механизмов устойчивости природных биоценозов. Если какой-либо из видов резко снижает свою численность или выпадает из состава сообщества, его роль берут на себя другие. Чем больше видов в составе биоценоза, тем ниже численность каждого из них, тем сильнее выражена их экологическая специализация. В этом случае говорят о «более плотной упаковке экологических ниш в биоценозе».
Экологические ниши видов изменчивы в пространстве и во времени. Они могут быть резко разграничены в индивидуальном развитии в зависимости от стадии онтогенеза, как, например, у гусениц и имаго чешуекрылых, личинок и жуков майского хруща, головастиков и взрослых лягушек. В этом случае меняется и среда обитания, и все биоценотическое окружение. У других видов экологические ниши, занимаемые молодыми и взрослыми формами, более близки, но тем не менее между ними всегда имеются различия. Ослабление межвидовой конкуренции приводит к расширению экологической ниши вида. На океанических островах с бедной фауной ряд птиц по сравнению с их сородичами на материке заселяет более разнообразные местообитания и расширяет спектр кормов, так как не сталкивается при этом с конкурирующими видами. У островных обитателей отмечается даже повышенная изменчивость формы клюва как показатель расширения характера кормовых связей.
Если межвидовая конкуренция сужает экологическую нишу вида, не давая проявиться всем его потенциям, то внутривидовая конкуренция, наоборот, способствует расширению экологических ниш. При возросшей численности вида начинается использование дополнительных кормов, освоение новых местообитаний, появление новых биоценотических связей.
Экологические ниши растений, на первый взгляд, менее разнообразны, чем животных. Они четко очерчены у видов, различающихся по питанию: сапрофитов (подъельник, гнездовка), паразитов (заразиха, повилика, раффлезия), полупаразитов (погремок, марьянник, омела), насекомоядных растений (росянка, пузырчатка, непентес). Но и зеленые фотоавтотрофные растения занимают разные экологические ниши в фитоценозах. Множественность экологических ниш определяется разным использованием растениями среды, размещением органов в воздухе и почве, ритмами сезонного развития, длительностью периода вегетации, сроками цветения и особенностями плодоношения, взаимосвязями с элементами абиотической среды и всеми компонентами биоценоза.
В водоемах растения, полностью погруженные в воду (элодея, роголистник, уруть), оказываются в иных условиях температуры, освещенности, газового режима, чем плавающие на поверхности (телорез, водокрас, ряска) или укореняющиеся на дне и выносящие листья на поверхность (кувшинка, кубышка, виктория). Различаются они и взаимосвязями со средой. Эпифиты тропических лесов занимают сходные, но все же не идентичные ниши, так как относятся к разным экологическим группам по отношению к свету и воде (гелиофиты и сциофиты, гигрофиты, мезофиты и ксерофиты). Разные эпифитные орхидеи имеют узкоспециализированных опылителей.
В соответствии с представлениями Л. Г. Раменского об экологической индивидуальности видов и с учетом того, что виды растений в сообществе по-разному участвуют в освоении и преобразовании среды и трансформации энергии, можно считать, что в сложившихся фитоценозах каждый вид растения имеет свою экологическую нишу.
В онтогенезе растения, как и многие животные, меняют экологическую нишу. С возрастом они более интенсивно используют и преобразуют среду. Переход растения в генеративный период заметно расширяет круг консортов, меняет размер и напряженность фитогенного поля. Средообразующая роль стареющих, сенильных растений убывает. Они теряют многих консортов, но увеличивается роль связанных с ними деструкторов. Продукционные процессы ослабляются.
У растений имеет место перекрывание экологических ниш. Оно усиливается в отдельные периоды при ограничении ресурсов среды, но поскольку виды используют ресурсы индивидуально, избирательно и с разной интенсивностью, конкуренция в устойчивых фитоценозах ослабляется.
На богатство экологических ниш в биоценозе оказывают влияние две группы причин. Первая – условия среды, предоставляемые биотопом. Чем мозаичнее и разнообразнее биотоп, тем больше видов могут размежевать в нем свои экологические ниши. Другой источник разнообразия ниш – сами виды, являющиеся ресурсом и создающие среду для других. Американский исследователь Р. Макартур, изучая ярусное разнообразие растительности по горному склону и число видов гнездящихся птиц, продемонстрировал прямую зависимость между этими показателями, положив начало многим исследованиям, подтверждающим связь видовой емкости сообществ с их внутренней структурой. Любой новый вид, внедряющийся в сообщество, увеличивает число имеющихся в нем экологических ниш не только за счет своего собственного положения среди других, но и предоставляя ресурсы для паразитов и хищников, норовых и гнездовых сожителей и т. п.
Ценотические стратегии видовВ фитоценологии разработаны классификации растений по их способности к совместному произрастанию и ценотической значимости. Общие положения этих классификаций могут быть применены и к животным, так как они характеризуют своего рода стратегии видов, определяющие их место в биоценозах. Наиболее часто используется система Леонтия Григорьевича Раменского и Д. Грайма.
Группы растений, которые занимают сходное положение в фитоценозах, называют фитоценотипами. Л. Г. Раменский предложил различать среди совместно живущих растений три типа – виоленты, патиенты и эксплеренты. Он популярно характеризовал их как силовиков, выносливцев и выполняющих (т. е. заполняющих свободное пространство), уподобляя львам, верблюдам и шакалам. Виоленты обладают высокой конкурентной способностью в данных условиях: «энергично развиваясь, они захватывают территорию и удерживают ее за собой, подавляя, заглушая соперников энергией жизнедеятельности и полнотой использования ресурсов среды». Патиенты «в борьбе за существование… берут не энергией жизнедеятельности и роста, а своей выносливостью к крайне суровым условиям, постоянным или временным». Они довольствуются теми ресурсами, которые остаются от виолентов. Эксплеренты «имеют очень низкую конкурентную мощность, но зато они способны очень быстро захватывать освобождающиеся территории, выполняя промежутки между сильными растениями, так же легко они вытесняются последними».
Более подробные классификации выделяют и другие, промежуточные типы. В частности, можно различать еще группу пионерных видов, которые быстро занимают вновь возникающие территории, на которых еще не было никакой растительности. Пионерные виды частично обладают свойствами эксплерентов – низкой конкурентной способностью, но, как и патиенты, обладают высокой выносливостью к физическим условиям среды.
В 70-е годы прошлого века, спустя 40 лет после Л. Г. Раменского, выделение тех же трех фитоценотипов повторил незнакомый с его классификацией ботаник Д. Грайм, обозначив их другими терминами: конкуренты, толеранты и рудералы.
Практически в любой группе организмов выделяются аналогичные по способности к совместному существованию виды, поэтому классификацию ценотических стратегий Раменского-Грайма можно относить к общеэкологической.
Источник: studopedia.su
Трофические связи возникают, когда один вид питается другим – либо живыми особями, либо их мертвыми остатками, либо продуктами жизнедеятельности. И стрекозы, ловящие на лету других насекомых, и жуки-навозники, питающиеся пометом крупных копытных, и пчелы, собирающие нектар растений, вступают в прямую трофическую связь с видами, предоставляющими им пищу. В случае конкуренции двух видов из-за объектов питания между ними возникает косвенная трофическая связь, так как деятельность одного отражается на снабжении кормом другого. Любое воздействие одного вида на поедаемость другого или доступность для него пищи следует расценивать как косвенную трофическую связь между ними. Например, гусеницы бабочек-монашенок, объедая хвою сосен, облегчают короедам доступ к ослабленным деревьям.
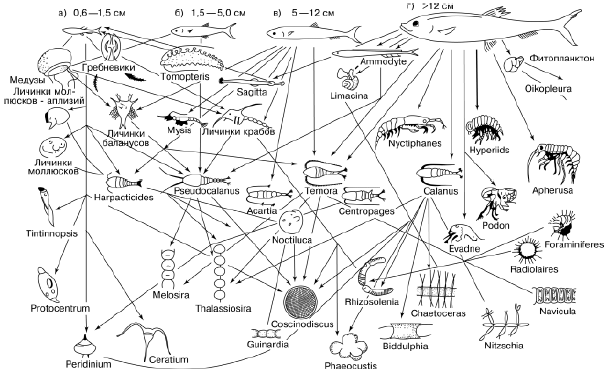
Трофические связи являются главными в сообществах. Именно они объединяют живущие вместе виды, поскольку каждый из них может обитать лишь там, где имеются необходимые ему пищевые ресурсы. Любой вид не только приспособлен к определенным источникам питания, но и сам служит пищевым ресурсом для других. Пищевые взаимосвязи создают в природе трофическую сеть, распространяющуюся в конечном счете на все виды в биосфере. Образ этой трофической сети можно воссоздать, поставив в центр любой вид и соединив его стрелками со всеми другими, которые находятся с ним в прямых или косвенных пищевых отношениях (рис. 88), и затем продолжать эту процедуру для каждого вовлеченного в схему вида. В результате будет охвачена вся живая природа, от китов до бактерий. Как показали исследования академика А. М. Уголева, существует «чрезвычайное единообразие свойств ассимиляционных систем на молекулярном и надмолекулярном уровне у всех организмов биосферы», позволяющее им получать энергетические ресурсы друг от друга. Он утверждает, что за бесконечным разнообразием типов питания стоят общие фундаментальные процессы образующих единую систему трофических взаимодействий планетарного масштаба.

Рис. 88. Пищевые связи сельди – часть трофической сети океана
Любой биоценоз пронизан пищевыми связями и представляет собой более или менее локализованный в пространстве участок общей трофической сети, связывающей все живое на Земле.
Источник: studopedia.ru
Источник: www.chem21.info