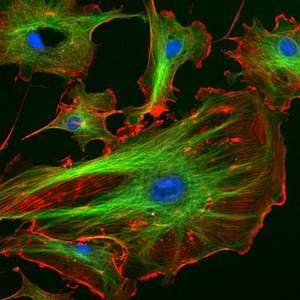
Цитоскелет состоит из
Общее понятие
Прежде чем говорить на эту тему, следует дать понятие цитоплазмы. Это внутренняя полужидкая среда клетки, которая ограничена цитоплазматической мембраной. В эту внутреннюю среду не входят ядро и вакуоли клетки.
А цитоскелет — это каркас клетки, который находится в цитоплазме клетки. Он есть в клетках эукариот (живые организмы, содержащие в клетках ядро). Является динамичной структурой, которая способна изменяться.
В некоторых источниках, рассматриваемых строение и функции цитоскелета, дается несколько иное, сформулированное другими словами определение. Он является опорно-двигательной системой клеток, которая образована белковыми нитчатыми структурами. Участвует в движении клетки.
Строение
Рассмотрим строение данной структуры, далее узнаем, какие функции выполняет цитоскелет.
Цитоскелет образовался за счет белков. В его структуре выделяется несколько систем, название которых происходит от основных структурных элементов, либо от основных белков, которые входят в состав данных систем.
Поскольку цитоскелет — это структура, то в ней выделяют три основные составляющие. Они играют важную роль в жизни и движении клеток.
Цитоскелет состоит из микротрубочек, промежуточных филаментов и микрофиламентов. Последние иначе называют актиновыми филаментами. Все они по своей природе нестабильны: постоянно собираются и разбираются. Таким образом, все компоненты имеют динамическое равновесие с белками, им соответствующими.
Микротрубочки цитоскелета, представляющие собой жесткую структуру, присутствуют в цитоплазме эукариотов, а также в ее выростах, которые называются жгутиками и ресничками. Их длина может варьироваться, некоторые достигают несколько микрометров в длину. Иногда микротрубочки объединяются с помощью ручек или мостиков.
Микрофиламенты состоят из актина — белка, похожего на тот, что входит в состав мышц. В их строении в малом количестве есть и другие белки. Главное отличие актиновых филаментов от микротрубочек состоит в том, что некоторых из них нельзя увидеть в световом микроскопе. В животных клетках они объединяются в сплетение под мембраной и, таким образом, связаны с ее белками.
Микрофиламенты животных и растительных клеток также взаимодействуют с белком миозином. При этом их система имеет способность к сокращению.
Промежуточные филаменты состоят из различных белков. Данный структурный компонент достаточно не изучен. Есть вероятность, что у растений он вообще отсутствует. Также некоторые ученые считают, что промежуточные филаменты являются дополнением к микротрубочкам. Точно доказано то, что при рзрушении системы микротрубочек филаменты перестраиваются, а при обратной процедуре влияние филаментов практически не сказывается на микротрубочках.
Функции
Говоря о строении и функциях цитоскелета, перечислим, каким именно образом он влияет на клетку.
Благодаря микрофиламентам, происходит движение белков вдоль мембраны цитоплазмы. Актин, содержащийся в них, принимает участие в мышечных сокращениях, фагоцитозе, движениях клетки, а также в процессе слияния сперматозоидов и яйцеклеток.
Микротрубочки активно участвуют в поддержании клеточной формы. Еще одна их функция — транспортная. Они переносят органеллы. Они могут выполнять механическую работу, куда входит перемещение митохондрий и ресничек. Особо важная роль принадлежит микротрубочкам в процессе клеточного деления.
Они направлены на создание или сохранение определенной клеточной асимметрии. Под определенным воздействием микротрубочки разрушаются. Это может привести к утрате данной асимметрии.
К функциям цитоскелета также относятся адаптация клетки ко внешнему воздействию, процессы эндо- и экзоцитоза.
Таким образом, мы рассмотрели, какие функции выполняет цитоскелет в живом организме.
Эукариоты
Между эукариотами и прокариотами существует определенная разница. Поэтому важно рассмотреть цитоскелет данных животных. Эукариоты (животные, имеющие в клетке ядро) имеют три типа филаментов.
Актиновые филаменты (иначе говоря, микрофиламенты) размещаются у мембраны клетки. Они принимают участие в межклеточном взаимодействии, а также передают сигналы.
Промежуточные филаменты — это наименее динамичная часть цитоскелета.
Микротрубочки являются полыми цилиндрами, они — очень динамичная структура.
Прокариоты
К прокариотам относятся одноклеточные организмы — бактерии и археи, которые не имеют сформированного ядра. Считалось, что прокариоты не имеют цитоскелета. Но с 2001-го года начались активные исследования их клеток. Были найдены гомологи (схожие, подобные) всех элементов эукариотного цитоскелета.
Ученые установили, что одна из белковых групп бактериального клеточного скелета не имеет аналогов среди эукариотов.
Заключение
Таким образом, мы рассмотрели строение и функции цитоскелета. Он играет исключительно важную роль в жизнедеятельности клетки, обеспечивая важнейшие ее процессы.
Все цитоскелетные компоненты взаимодействуют. Это подтверждается существованием прямых контактов микрофиламентов, промежуточных филаментов и микротрубочек.
Согласно современным представлениям, важнейшим звеном, которое объединяет различные клеточные части и осуществляет передачу данных, является именно цитоскелет.
Источник: FB.ru

-Совокупность нитевидных белковых структур – микротрубочек и микрофиламентов, составляющих опорно-двигательную систему клетки.
Цитоскелет — высокодинамичная система цитоплазмы. Многие структуры цитоскелета могут легко разрушаться и вновь возникать, меняя свое расположение или морфологию. В основе этих особенностей цитоскелета лежат реакции полимеризации-деполимеризации основных структурных цитоскелетных белков и их взаимодействие с другими белками, как структурными, так и регуляторными.
Цитоскелетом обладают только эукариотические клетки, в клетках прокариот (бактерий) его нет, что является важным различием этих двух типов клеток. Цитоскелет придаёт клетке определённую форму даже при отсутствии жёсткой клеточной стенки. Он организует движение органоидов в цитоплазме (т. н. течение протоплазмы), лежащее в основе амёбоидного движения. Цитоскелет легко перестраивается, обеспечивая в случае необходимости изменение формы клеток. Способность клеток изменять форму обусловливает перемещение клеточных пластов на ранних стадиях зародышевого развития. При делении клетки (митозе) цитоскелет «разбирается» (диссоциирует), а в дочерних клетках вновь происходит его самосборка.
Функции цитоскелета многообразны. Он способствует поддержанию формы клетки, осуществляет все типы клеточных движений . Кроме того, цитоскелет может принимать участие в регуляции метаболической активности клетки .
Цитоскелет образован белками. В цитоскелете выделяют несколько основных систем, называемых либо по основным структурным элементам, заметным при электронно-микроскопических исследованиях (Микрофиламенты, промежуточные филаменты, микротрубочки), либо по основным белкам, входящим в их состав (актин-миозиновая система, кератины, тубулин-динеиновая система).
Промежуточные филаменты являются наименее понятной структурой среди основных компонентов цитоскелета в отношении их сборки, динамики и функций. Их свойства и динамика сильно отличаются от соответствующих характеристик как микротрубочек, так и актиновых филаментов. Функции же промежуточных филаментов до сих пор остаются в области гипотез.
Цитоплазматические промежуточные филаменты обнаружены в подавляющем большинстве эукариотических клеток, как у позвоночных, так и беспозвоночных животных, у высших растений. Редкие примеры клеток животных, у которых не обнаружены промежуточные филаменты, не могут считаться окончательными, так как белки промежуточных филаментов могут образовывать необычные структуры.
Морфологические микротрубочки представляют собой полые цилиндры диаметром около 25 нм с толщиной стенки около 5 нм. Стенка цилиндра состоит их протофиламентов — линейных полимеров тубулина с продольно ориентированными гетеродимерами. В составе микротрубочек протофиламенты идут вдоль их длинной оси с небольшим сдвигом друг относительно друга, так что субъединицы тубулина образуют трехстартовую спираль. В состав микротрубочек большинства животных входит 13 протофиламентов
Актиновые филаменты играют ключевую роль в сократительном аппарате мышечных и немышечных клеток , а также принимают участие во многих других клеточных процессах, таких как подвижность , поддержание формы клетки , цитокинез
Актиновые филаменты или фибриллярный актин ( F-актин ) представляют собой тонкие фибриллы диаметром 6-8 нм. Они являются результатом полимеризации глобулярного актина — G-актина . В клетке актиновые филаменты с помощью других белков могут образовывать множество разнообразных структур.
Источник: biology623.blogspot.com
Общий план строения филаментов цитоскелета
Элементы цитоскелета являются полимерами, мономерами которых выступают определенные белковые субъединицы. В отличие от других биополимеров, таких как сами белки или нуклеиновые кислоты, структурные единицы цитоскелета соединены друг с другом слабыми нековалентными связями. Полимерная строение выгодна из-за того, что позволяет клетке быстро перегруппировывать цитоскелет: белковые мономеры маленькие, и они могут быстро диссоциировать в цитоплазме, в отличие от длинных филаментов.
Промежуточные филаменты состоят из субъединиц, которые сами являются удлиненными фибриллярного белка, в то время как мономерами микрофиламентов и микротрубочек является глобулярные белки актин и тубулин соответственно. Белки цитоскелета могут самоорганизовываться в длинные филаменты, образуя различные типы латеральных контактов и контактов типа «хвост-голова». В живой клетке этот процесс регулируется огромным количеством вспомогательных белков.
Элементы цитоскелета могут быть одновременно динамичными и очень прочными за того, что они состоят из нескольких протофиламентив — длинных линейных нитей, построенных с мономеров, размещенных в один ряд. Обычно протофиламенты спирально закручиваются друг вокруг друга. Микротрубочки состоят из тринадцати протофиламентив размещенных по кругу, микрофиламенты — из двух спирально закрученных, а промежуточные филаменты — с восьми. Вследствие такого строения диссоциация мономера с конца фибриллы происходит значительно легче, чем разрыв посередине, так как для диссоциации необходимо разрушения только одного продольного связи и одного-двух латеральных, а для разрыва — большого количества продольных связей. Поэтому перестройка элементов цитоскелета происходит относительно легко, и в то же время они могут легко противостоять тепловым повреждением и выдерживать различные механические воздействия.
Элементы цитоскелета эукариот
Основными функциями цитоскелета является поддержание формы клетки и обеспечения перемещения как клетки в целом, так и внутриклеточных компонентов внутри клетки. Цитоскелет состоит из трех основных компонентов: микрофиламентов, микротрубочек и промежуточных филаментов. Это супрамолекулярные, протяжные полимерные структуры, состоящие из белков одного типа.
| Сравнительная характеристика основных элементов цитоскелета | |||
|---|---|---|---|
| Микротрубочки | Актиновые филаменты | Промежуточные филаменты | |
| Фотография | |||
| Схема строения | |||
| Струкутра | Трубка из 13 протофиламентив белка тубулина | Два закрученных одна вокруг одного протофиламенты актина | Несколько протофиламентив, состоящие из фибриллярных белков объединены в канатоподибну структуру |
| Диаметр | 25Нм с просветом в 15 нм | 7 нм | 8-12нм |
| Белковые субъединицы | Тубулин — димер, состоящий из α- и β-тубулина | Актин | Различные белки в зависимости от типа клеток и функции (например кератин, белки ламины, виментину т.д.) |
| Нуклеотиды нужны для полимеризации | ГТФ | АТФ | Не нужны |
| Основные функции |
|
|
|
Динамика элементов цитоскелета
Элементы цитоскелета являются динамическими структурами: их можно сравнить с цепочкой муравьев, которые идут к месту сбора пищи. Хотя сам цепочка может существовать часами, каждый муравей в нем находится в постоянном движении. Так же и элементы цитоскелета постоянно обмениваются субъединицами с цитоплазмой, где мономеры находятся в растворимой форме. Относительной стабильностью характеризуются только промежуточные филаменты, поэтому информация о динамике касается в большей степени микротрубочек и актиновых филаментов.
Примером динамичности и гибкости цитоскелета клетки может быть перегруппировки микротрубочек, которые в интерфазе образуют структуру похожую на звезду, лучи которой отходят от центра клетки, а перед разделением способны быстро создать веретено деления.
то же время некоторые структуры, построенные из элементов цитоскелета могут существовать очень долгое время: например на поверхности волосковых клеток внутреннего уха является вырасти — стереоцили, поддерживаемых пучками микрофиламентов. Эти пучки существуют на протяжении всей жизни животного, хотя их субъединицы постоянно обновляются
Скорость присоединения и диссоциации субъединиц описывается константами k on (измеряется в М -1 × с -1) и k off (измеряется в с -1) соответственно. Причем скорость присоединения зависит не только от k on, но и от концентрации свободных мономеров в цитоплазме, а скорость диссоциации является постоянной. Когда филамент растет, то количество свободных мономеров в цитоплазме падает, пока не достигнет определенного уровня — критической концентрации (C C), при которой скорость присоединения будет равна скорости диссоциации: C C × k on = k off, откуда:
Нуклеация
Мономеры элементов цитоскелета могут спонтанно образовывать комплексы в растворе. Однако, такие олигомеры обычно нестабильны, потому что каждая субъединица в них образует связи только с небольшим количеством других. Этих взаимодействий часто недостаточно, чтобы удержать комплекс, и он в основном быстро распадается. Для образования длинных филаментов необходимо наличие первоначального агрегата с такого количества мономеров, которой будет достаточно для стабилизации, такой агрегат называется ядром, а процесс его образования — нуклеации. Для актиновых филаментов, ядро должно состоять минимум из трех субъединиц, тогда как образование микротрубочек начинается с сложного комплекса (предположительно, из 13 молекул тубулина, образующих кольцо).
Нуклеация обычно является лимитирующим этапом в образовании длинных филаментов в растворе свободных мономеров. После инициации полимеризации в таком растворе наблюдается лаг-фаза, во время которой не наблюдается образование филаментов. Ее существование объясняется тем, что нестабильность небольших олигомеров создает кинетический барьер в полимеризации, и длится она до тех пор, пока не произойдет процесс нуклеации. Если к раствору мономеров добавить готовые комплексы субъединиц (например, состоящие из соединенных ковалентно мономеров), тогда лаг-фазы наблюдаться не будет.
Потребность в нуклеации используется клеткой для регулирования образования новых элементов цитоскелета. Существуют специальные белки, которые могут катализировать нуклеации в специфическом месте, где необходимо образование микротрубочек или актиновых филаментов.
Полярность микротрубочек и микрофиламентов
В отличие от мономеров промежуточных филаментов, актин и тубулин имеют два структурно и функционально разные концы. В составе микрофиламентов и микротрубочек все субъединицы возвращены в одну сторону, таким образом данные элементы цитоскелета обладают полярностью. Два конца этих филаментов отличаются по динамике полимеризации и деполимеризации:
- конец, на котором полимеризация и деполимеризация происходят быстрее называется плюс концов;
- конец, на котором полимеризация и деполимеризация происходят медленнее называется минус концов.
В микротрубочках α-субъединицы тубулина возвращены в минус-конца, а β — до плюс. В Микрофиламентов мономеры актина размещены таким образом, что их АТФ-связывающая щель указывает в сторону минус конца.
Несмотря на то, что абсолютные занчення k on и k off могут сильно отличаться для плюс и минус конца, их соотношение является постоянной величиной. Поскольку изменение свободной энергии ΔG вследствие диссоциации или присоединения новой субъединицы одинакова, не в зависимости от того, на каком конце филамента произошли изменения. Поэтому, когда концентрация свободных мономеров C <C C, оба конца укорачиваются, а когда C> C C, оба конца растут. Это подтверждается только при отсутствии гидролиза нуклеозидтрифосфатов (АТФ или ГТФ).
Гидролиз нуклеотидтрифосфатив
Актин и тубулин — это не просто мономеры элементов цитоскелета, они также являются ферментами, которые могут осуществлять гидролиз АТФ и ГТФ соответственно. Одна молекула актина связывает одну молекулу АТФ, тогда как димер тубулина — две молекулы ГТФ (по одной на каждую субъединицу), то ГТФ, что находится в α-субъединицы никогда не гидролизуетья и не обменивается, тогда как ГТФ β-субъединицы может превращаться на ГДФ.
В свободных мономерах актина и тубулина гидролиз нуклеотидов происходит очень медленно, для ускорения этого процесса необходимо действие определенного фактора — ГТФаза- или АТФаза-активирующих белков. Причем для тубулина и актина такими факторами являются другие молекулы тубулина или актина соответственно, поэтому гидролиз нуклеотидтрифосфату значительно ускоряется после инкорпорации мономера в филамент цитоскелета, где он взаимодействует с другими идентичинмы молекулами. Микротрубочки и микрофиламенты могут существовать в двух формах «Т-форме» (мономеры связаны с ГТФ или АТФ) и «Д-форме» (мономеры связаны с ГДФ или АДФ).
После гидролиза нуклеотидтрифосфату большая часть энергии, высвобождаемой «хранится» в структуре филаментов. Поэтому изменение свободной энергии для диссоциации мономера с Д-формы становится негативный, чем для диссоциации с Т-формы, а следовательно и соотношение k off / k on, равное значению критической концентрации, будет больше для Д-формы, чем для Т. Иными словами, Д-форма более «склонна» к диссоциации. При определенном значении концентрации свободных субъединиц C, когда C C (T) <C <C C (D), Т-форма филаментов будет расти, а Д-форма — вкорочуватись.
Тредмилинг
Вероятность того, что определенная субъединица филаментов цитоскелета гидролизует связан нуклеотидтрифосфат и перейдет в Д-форму, тем больше, чем дольше эта субъединица находится в составе полимера. Поэтому посередине филаментов, где все мономеры уже «древние», они имеют в своем составе нуклеотиддифосфаты. К концам присоединяются преимущественно новые молекулы в Т-форме (поскольку концентрация АТФ или ГТФ в цитоплазме в десятки раз превышает концентрацию АДФ и ГДФ соответственно). На минус-конце полимеризация происходит медленно, поэтому гидролиз «успевает» за ней, и не происходит накопления субъединиц в Т-форме. Зато на плюс-конце, где полимеризация значительно быстрее, образуется «кэп» из нескольких субъединиц, содержащих негидролизовани нуклеотидтрифосфаты. Таким образом один конец (+) филамента находится в Т-форме, а другой (-) — в Д-форме и при концентрации свободных филаментов C, где C C (T) <C <C C (D), плюс-конец расти , а минус- — вкорочуватиметься, а общая длина не будет меняться. Этот процесс называется тредмилинг, он наблюдается как в микротрубочек, так и в актиновых филаментов, но характерно для последних. Тредмилинг всегда нуждается в энергии, выделяемой за счет гидролиза АТФ.
Динамическая нестабильность
В зависимости от скорости полимеризации и гидролиза нуклеотидтрифосфатив конце филаментов цитоскелета могут изменять свое состояние: переходить с Т-формы в Д-форму и наоборот. Если концентрация свободных филаментов при этом меньше C C (T) и больше C C (D), то такой переход будет иметь важные последствия: филамент будет переходить от роста к укорочению (это событие называется катастрофа) либо наоборот (восстановления). Способность элементов цитоскелета быстрой смены «режимов» полимеризация / деполимеризация при постоянной концентрации свободных субъединиц называется динамической нестабильностью.
Явление динамической нестабильности особенно характерно для микротрубочек. В Т-форме их протофиламенты прямые, а при переходе к Д-формы они искривляются. Когда микротрубочки имеет ГТФ-кэп, он стабилизирует всю структуру, однако после ее потери (из-за замедления полимеризации или ускорения гидролиза) протофиламенты в Д-форме начинают очень быстро «розлуплюватись».
Также наблюдаются некоторые флуктуации длины актиновых филаментов, однако они в десяток раз меньше, чем в микротрубочек.
Яды, влияющие на цитоскелет эукариот
Поскольку нормальное функционирование системы микротрубочек и промежуточных филаментов необходимо для выживания и разделения килтины, эти клеточные компоненты часто являются мишенями действия природных токсинов. Некоторые из этих ядов связываются со свободными мономерами актина или тубулина и препятствуют им полимеризоваться, другие же наоборот — взаимодействуют с полимерными формами и не допускают диссоциации мономеров. Например вещество таксол с тихоокеанского тиса (Taxus brevifolia) стабилизирует микротрубочки в полимерных форме, в то время как колхицин с безвременника осеннего (Colchicum autumnale) и винбластин с катарантуса (Catharanthus) наоборот не дают мономерам тубулина объединяться. Существуют вещества, аналогичным образом действуют и на актиновые филаменты: фалоидин с бледной поганки (Amanita phalloides) способствует филаментозний форме актина, а латрункулин с морской губки Latrunculia magnifica — наоборот, растворимой мономерной.
Кроме того, что подобные вещества широко используются для изучения свойств цитоскелета, некоторые из них также есть и терапевтическими препаратами. Таксол и винбластин благодаря своей способности изменять характер полимеризации микротрубочек способны достаточно эффективно убивать клетки, которые быстро делятся, при этом проявляя небольшое влияние на другие клетки. Поэтому их используют для лечения раковых заболеваний. Особенно популярен таксол для терапии рака молочной железы и рака легких, он часто бывает эффективным даже в тех случаях, когда другие методы химиотерапии не действуют.
Цитоскелет прокариот
До недавнего времени считалось, что цитоскелет имеют только эукариоты. Но последние исследования показывают, что для всех составных частей эукариотического цитоскелета можно найти гомологи у прокариот. Хотя сходство в аминокислотной последовательности белков небольшая, восстановления трехмерной структуры белковых молекул позволяет говорить о значительной структурное сходство и гомологичнисть этих структур.
- Гомологи тубулина: с гомологов тубулина распространенным среди прокариот является белок FtsZ, что был первым найденным компонентом прокариотической цитоскелета. Подобно тубулина, FtsZ формирует филаменты тратя ГТФ, но эти филаменты НЕ группируются в трубочки. В течение деления клетки, FtsZ — первый белок, перемещается на место разделения, формируя «кольцо разделения» или Z-кольцо, которое обеспечивает прохождение цитокинеза, также FtsZ важен для привлечения ферментов, которые синтезируют новую клеточную стенку между дочерними клетками.
- Гомологи актина: MreB и ParM — это актино-образные белки прокариот. MreB нужен для придания формы клетке, в частности отвечает за различие между пиличкоподибнимы и спиральными бактериями. Все несферических бактерии имеют гены MreB или его близких гомологов. Продукты этих генов формируют спиральную сеть под клеточной мембраной, которая служит для удержания ферментов, задействованных в биосинтезе клеточной стенки. Белок ParM кодируется плазмидной ДНК многих бактерий и нужен для сегрегации копий плазмиды при делении.
- Гомолог белков промежуточных филаментов кресцентин: бактерия Caulobacter crescentus содержит третий белок, кресцентин, гомологический промежуточных филаментов эукариот. Кресцентин также используется для поддержания формы клетки.
- Элементы цитоскелета прокариот, не имеющих гомологов у эукариот: у прокариот также имеющийся класс элементов цитоскелета, принадлежащих к семье WACA (англ. Walker A cytoskeletal ATPase) и не имеют гомологов в еукароит. К этому классу относится белок MinD, который является составной частью системы MinCDE, что обеспечивает определение места прохождения цитокинеза, а также белки, необходимые для различия копий плазмид, такие как ParA, Soj и другие.
Эволюция цитоскелета
Родственны между собой элементы цитоскелета были найдены у подавляющего большинства представителей всех трех доменов живых организмов: эукариот, бактерий и архей. Это свидетельствует о том, что белки цитоскелета возникли еще до выделения этих трех ветвей, каким бы путем оно не происходило.
Белок FtsZ, с которого позже возник тубулин, вероятно, эволюционно очень древним. Он содержит очень мало аминокислот аргинина, лизина, фенилаланина, тирозина и гистидина и практически не содержит триптофана. Поскольку считается, что кодоны этих аминокислот были добавлены в генетический код последними, вполне вероятно, что какая-то форма FtsZ возникла еще до окончательного установления генетического кода и уже тогда служила для осуществления цитокинеза. Белки гомологи тубулина образуют отдельную семью ГТФаз, и не имеют никаких близких родственников. Зато MreB более «молодой» с эволюционной точки зрения белок, он, вместе с другими актиноподибнимы белками и актиний, принадлежит к семье АТФаз, которая также включает ферменты гексокиназы и шаперон hsp70. Причем первыми из этой семьи, больше всего, возникли гексокиназы.
Сравнение последовательностей аминокислот в белках FtsZ различных видов бактерий и архей между собой и с эукариотическими тубулина, а также MreB между собой и с эукариотическими актина выявило интересную закономерность:
- Белки FtsZ очень далеких друг от друга видов прокариот, таких как бактерии Escherichia coli, Bacillus subtilis, Mycoplasma pulmonis и Архебактерии рода Halobacterium имели высокую степень идентичности в аминокислотной последовательности (от 46 до 53%); аналогичное справедливо и для белка MreB.
- Эукариотические тубулина и актин даже еще более консервативные (напирклад между тубулина человека и дрожжей существует 75% идентичности, в то время как актин любых видов эукариот, обычно отличаются не более чем на 10%);
- Несмотря на большую консервативность белков цитоскелета в пределах групп эукариот и прокариот, при сравнении этих белков между группами, оказывается, что идентичность настолько мала, что ее почти невозможно обнаружить обычными методами (менее 15%). Причем гомология наиболее выражена в ГТФ- и АТФ-связывающих доменах.
Для объяснения этой «загадки» была выдвинута гипотеза о том, что такая резкая дивергенция эукариотических белков цитоскелета от прокариотических состоялась вследствие изменения их роли в клетке. FtsZ перестал обеспечивать прохождение цитокинеза и стал механической опорой клетки, а позже взял на себя и другие функции, в то время как MreB, взял на себя роль осуществления деления клетки и фагоцитоза.
Чрезвычайно высокий уровень косервативности актина и тубулина в клетках эукариот объясняется тем, что эти белки взаимодействуют с огромным количеством других: регуляторных, вспомогательных, моторных и др. Именно актин является «чемпионом» среди эукариотических белков по количеству белков-партнеров, поэтому замена любой аминокислоты может привести к нарушению этих взаимодействий и иметь катастрофические последствия.
Третий тип элементов цитоскелета — промежуточные филаменты, эволюционировали другим путем. Они имеющиеся фактически только у эукариот, и хотя их гомолог кресцентин и был обнаружен у одного вида бактерий, скорее всего, эти бактерии получили его в результате горизонтального переноса генов от эукариот. Белки промежуточных филаментов, в отличие от актина и тубулина, не отличаются особой консервативностью.
Источник: info-farm.ru