Вторичная клеточная стенка
Клеточная оболочка способна к утолщению и видоизменению. В результате этого образуется ве вторичная структура. Утолщение оболочки происходит путем наложения новых слоев на нервпчпуго оболочку. Ввиду того что наложепие идет уже иа твердую оболочку, фибриллы целлюлозы в каждом слое лежат параллельно, а в соседних слоях — под углом друг к другу. Этим достигается значительная прочность и твердость вторичной оболочки. По мере того как число слоев фибрилл целлюлозы становится больше и толщина стенки увеличивается, она теряет эластичность и способность к росту. Во вторичной клеточпой стенке содержание целлюлозы значительно возрастает, в некоторых случаях до 60% и более. По мере дальнейшего старения клеток матрикс оболочки может заполняться различными веществами — лигнином, суберином (одревеснение или опробковение оболочки). Лигнин образуется из гемицеллюлозы н пектиновых веществ.[ …]
Клеточная оболочка древесного волокна имеет несколько слоев: первичный, который называется наружной оболочкой волокна, и вторичный (стенка, состоящая, в свою очередь, из трех слоев: наружного, среднего и внутреннего). Между первичными стенками клеток находится слой межклеточного вещества, при помощи которого волокна соединяются друг с другом. Вторичная стенка относительно толстая и представляет собой главную массу объема клетки.[ …]
Во вторичных слоях клеточных стенок древесины сосны накапливались в больших количествах маннаны (22%) и уроновый ангидрид (25%).[ …]
Фаза утолщения клеточной стенки. Как происходит утолщение. В период разрастания протопласт окружен только первичной стенкой. Когда же древесная клетка достигает своего наибольшего размера по поверхности или вскоре после этого, стенка клетки утолщается. Это вызвано наслаиванием вторичной стенки на первичную, причем этот новый слой возникает в результате дальнейшей деятельности протопласта внутри полости клетки. Естественно, что клетки, в которых протопласт исчез, не могут продолжать утолщать свои стенки. Образование вторичной стенки является признаком необратимого изменения в клетке, дальнейшее разрастание которой уже исключено, но не обязательно исключается дальнейшее деление при условии, что получаемые таким образом дочерние клетки занимают такой же объем, как и первоначальная клетка.[ …]
Фаза утолщения клеточной стенки. Как происходит утолщение. В период разрастания протопласт окружен только первичной стенкой. Когда же древесная клетка достигает своего наибольшего размера по поверхности или вскоре после этого, стенка клетки утолщается.
о вызвано наслаиванием вторичной стенки на первичную, причем этот новый слой возникает в результате дальнейшей деятельности протопласта внутри полости клетки. Естественно, что клетки, в которых протопласт исчез, не могут продолжать утолщать свои стенки. Образование вторичной стенки является признаком необратимого изменения в клетке, дальнейшее разрастание которой уже исключено, но не обязательно исключается дальнейшее деление при условии, что получаемые таким образом дочерние клетки занимают такой же объем, как и первоначальная клетка.[ …]
М.1ип — ковер, покрывало). Он состоит из таблитчатых тонкостенных клеток с густой цитоплазмой. Обычно он однорядный, но иногда бывает дву рядным или многорядным. Клетки «ноту ма сначала одноядерные, по позднее они часто становятся двухъядерными или даже многоядерными. Тапетум представляет собой физиологически чрезвычайно активную ткань: его клетки содержат ферменты, гормоны, и питательный материал, используемый в процессе микроспорогепеза. Имеются некоторые основания считать секреторный тип в эволюционном отношении первичным, а амебоидный — вторичным.[ …]
Необходимо, однако, отметить, что эти данные следует рассматривать как приближенные, так как исходные препараты не были тщательно очищены.[ …]
Трудно определить расположение в клеточной стенке полиуронидных гемицеллюлоз, потому что реагенты, используемые для их выявления, оказывают воздействие и на лигнин [54].
которые исследователи предполагают, что цементирующим веществом между фибриллами и различными слоями клеточной стенки являются гемицеллюлозы. Коэн [55] считает даже, что лигнин вторичной стенки имеет одинаковую природу с гемицеллюлозами. Основанием для такого предположения служит, по-видимому, тот факт, что некоторые углеводы при обработке сильными кислотами могут давать нерастворимые остатки определенного рисунка. Следует подчеркнуть, однако, что участки, как тщательно обработанные реагентами, растворяющими гемицеллюлозы, так и не обработанные ими, дают при воздействии 72%-ной серной кислоты остатки очень похожей структуры [56].[ …]
Для выяснения состава отдельных слоев клеточных стенок была сделана попытка количественного определения ксилоуронидов в разных слоях трахеид и либриформа [49]. Измерения производились на волокнах из красной японской сосны, европейской пихты, бука и березы. Для этого волокна осторожно нитровали в среде уксусного ангидрида и четыреххлористого углерода. Затем наружный нитрованный слой удаляли растворением в ацетоне, после чего контролировали содержание пентозанов в остатке по фурфуролу. Было установлено, что пентозаны в древесных волокнах по слоям разделены неравномерно. Наибольшее количество пентозанов найдено в наружных слоях волокон и концентрация их падает от периферии к центру. Так, наружные слои волокон хвойной древесины содержат 50—80% пентозанов, а у лиственных почти 100%. Во вторичных слоях клеточных стенок у хвойных содержание пентозанов оказалось не более 2—4%, а у лиственных 8—10%. Таким образом, химический метод подтвердил результаты, полученные ранее методом сорбции ультрафиолетового света.[ …]
Модулярное строение растений, наличие у них клеточных стенок и отсутствие настоящей системы циркуляции (типа кровеносной или лимфатической), по-видимому, делают любую форму иммунного ответа неэффективной защитой. Здесь нет мигрирующих популяций фагоцитов, которые могут быть мобилизованы на борьбу с чужеродными объектами. Однако многие растения отвечают уже на самые ранние стадии заражения реакцией сверхчувствительности. Гибнущие посаженные клетки вместе с непосредственно окружающими их выделяют «фитоалексины»— низкомолекулярные, обладающие антимикробным действием вторичные метаболиты, накапливающиеся в зоне появления патогена до уровня, подавляющего его развитие (Bailey, Mansfield, 1982). Фитоалексины могут предотвратить распространение паразитов от участка первичного заражения, но эта реакция совершенно не похожа на иммунный ответ: она строго локализована, довольно неспецифична и вызывается различными стимулами.[ …]
Различают лигнин первичный, находящийся в одревесненных клеточных стенках (природный лигнин) и вторичный — изолированный лигнин. Последний является в значительной степени веществом, измененным в процессе изолирования и загрязненным примесями посторонних веществ. Изменение лигнина выражается в отщеплении метоксильных групп, внутримолекулярной конденсации и в других признаках.[ …]
Многие различия между типами тканей обусловлены строением клеточной стенки, особенно вторичной. Как мы уже говорили, образование первичной клеточной стенки происходит в процессе растяжения клетки, и, следовательно, она должна обладать свойством растяжимости, тогда как вторичная стенка формируется уже после того, как удлинение прекратилось.[ …]
Престон [12, 13, 14, 15), изучавший при помощи рентгеновских лучей клеточные стенки нескольких пород деревьев, поставил под сомнение утверждение Бейли и Керра о различном расположении цепных молекул в трехслойной стенке трахеиды. Он предположил, что целлюлозные цепи по всем трем слоям клеточной стенки имеют однородное направление и образуют довольно крутую спираль. Дальнейшее исследование этого вопроса, проведенное Бейли и Беркли [16], в том числе методом отложения микрокристаллов иода в удлиненных промежутках целлюлозной матрицы, привели к подтверждению этого предположения. Бейли и Беркли установили, что ориентация фибрилл соответственно во внутреннем и внешнем слоях колеблется от 45 до 90° по отношению к оси волокон, тогда как в центральном слое угол находится между 0 и 45°. Уордроп и Престон [17], изучая двойное лучепреломление в образцах древесины, срезанных под различными углами, нашли, что вторичная стенка действительно состоит из трех слоев и направления целлюлозных цепей мало отличаются от направлений, установленных Бейли и Беркли.[ …]
Одновременно с этими внутренними изменениями наружная твердая стенка ооспоры расщепляется на ее вершине на пять зубцов, давая выход проростку, возникающему из центральной клетки (рис.
9, 3). Первое деление центральной клетки происходит поперечной перегородкой, перпендикулярной к ее длинной оси, и приводит к образованию двух функционально различных клеток. Из одной, более крупной клетки в дальнейшем образуется стеблевой побег, который на начальной стадии развития называют предростком, из другой, меньшей клетки — первый ризоид. Оба они растут путем поперечных клеточных делений. Предросток растет вверх и довольно быстро зеленеет, заполняясь хлоропластами, первый ризоид направляется вниз и остается бесцветным (рис. 269, 4). После ряда клеточных делений, сообщающих им строение однорядных нитей, происходит их дифференцировка на узлы и междоузлия, и дальнейший их верхушечный рост протекает уже так, как было описано выше для стебля. Из узлов предростка возникают вторичные предростки, мутовки листьев и боковые ветви стебля, из узлов первого ризоида — вторичные ризоиды и их мутовчатые волоски. Таким путем и формируется таллом, состоящий из нескольких стеблевых побегов в верхней части и нескольких сложных ризоидов в нижней части (рис. 2G9, 5).[ …]
Надмолекулярная структура. На рис 6.10 приведена модель структуры клеточной стенки. Она включает 2 основных слоя: первичную стенку Р и вторичную Последняя подразделяется на 3 слоя: 5], 5 , Слой М, срединная пластинка, является межклеточным веществом, соединяющим клетки между собой.[ …]
В последующих разделах (ем. часть II) будет исчерпывающе рассмотрена химия клеточных стенок, относительные количества лигнина в них и другие родственные темы. Однако заканчивая рассмотрение четвертой и конечной фазы онтогенеза древесной клетки, следует упомянуть о некоторых явлениях, которые тем или иным путем связаны с лигннфикацией, как ее пошшают ботаники. Подобно образованию и разрастанию клеток, а также утолщению клеточных стенок, лигнификация может происходить лишь при жизни клеточного протопласта, так как отмершие клетки не могут лигнифи-цировать свои стенки. Процесс лигнификации может быть закончен в слое межклеточного вещества и в первичной стенке, но может продолжаться во вторичной стенке, даже если этот названный последним слой еще центростремительно увеличивается в толщину. В древесине деревьев лигнификация часто очень скоро заканчивается в слое, примыкающем к внутренней стороне камбия, обычно почти одновременно с тем, когда новые клетки достигли своего наибольшего размера, а вторичные стенки — своей конечной толщины. Это объясняет, почему заболонь при одинаковом содержании влаги так же или почти так же крепка, как ядровая древесина.[ …]
Детальное исследование распределения лигнина и полисахаридов в одревесневших клеточных стенках древесины ели и березы измерением интенсивности абсорбции тонкого пучка ультрафиолетовых лучей при прохождении их через прозрачный срез подтвердило преимущественное расположение лигнина в срединной пластинке и первичной стенке, а также частично в наружных слоях вторичной стенки [42, 43].
срединной пластинке еловой древесины содержание лигнина достигает 73%, а во вторичной стенке — не более 16%. Отсюда следует, что полисахариды сосредоточены в основном во вторичном слое. Была сделана попытка измерить этим методом взаимное расположение целлюлозы и гемицеллюлоз. Для этого полисахариды вначале были превращены в окрашенные соединения, абсорбирующие свет.[ …]
В большинстве клеток ясно различаются чередующиеся зоны большего или меньшего отложения лигнина, которые создают видимость концентрических колец. При противоположном процессе, когда клеточная стенка обрабатывается делигнифицирующими. реагентами, рисунок целлюлозы остается прежним. Это свидетельствует о том, что существуют, по-видимому, две взаимопроникающие системы, состоящие одна из целлюлозы и других полисахаридов, а другая из лигнина. Бейли и Керр [4] показали, что размеры частиц доходят до 0,1 ¡х и меньше. Промежутки или полосы объясняют существование относительно больших «фибрилл», замеченных некоторыми исследователями [49]. Кроме преобладающих концентрических рисунков, в волокнах некоторых видов древесины проявляется расположение радиальных линий или комбинация обоих типов. Клетки сжатой древесины часто имеют жесткие, почти твердые полосы лигнина рядом с полостью клетки и радиально-расположенные пластинки его, отделенные зонами полисахаридного вещества, в средней части стенки клетки.[ …]
В состав лишайников входят многие элементы и вещества. Все их можно разделить на две большие группы — первичные и вторичные. К первичным относятся те вещества, которые непосредственно принимают участие в клеточном обмене веществ; из них построено тело лишайников. К вторичным относятся конечные продукты обмена веществ, располагающиеся обычно на стенках гиф. Многие из этих вторичных лишайниковых веществ (в более старой литературе их называли лишайниковыми кислотами) специфичны для лишайников и не встречаются в организмах из других систематических групп.[ …]
Риттер [49], Людтке [50] и др. сообщили, что при обработке древесных волокон различными реагентами, вызывающими набухание, вторичная стенка (а также, вероятно, и первичная) распадается на нитеобразные фрагменты или фибриллы. Риттер [51 ] разделил эти фибриллы на веретенообразные тела, а их в свою очередь, на сферические единицы [52]. Значение таких относительно крупных структурных единиц (длина веретенообразных тел примерно 4[х) неясно, ввиду описанной выше тонкопористой структуры вторичной стенки. Ни в остатках лигнина после растворения целлюлозы, ни в остатках целлюлозы после растворения лигнина не обнаруживается заметных промежутков, указывающих на границы названных единиц клеточных стенок. Кроме того, недавно проведенными исследованиями с помощью электронного микроскопа в структуре клеточных стенок не было установлено присутствия подобных сравнительно крупных единиц.[ …]
При оценке действия различных дереворазрушающих грибов на растительную ткань необходимо учитывать, что отдельные гифы их. движутся в толще клеточных стенок избирательно. Так, грибы белой гнили предпочитают срединную пластинку и первичную оболочку, где сосредоточен главным образом лигнин. Грибы красной или бурой гнили, наоборот, предпочитают проходить по вторичной оболочке, наиболее богатой углеводами. Соответственно различается и окраска поврежденной ими древесины. Более подробно эти вопросы будут рассмотрены в дальнейшем.[ …]
Исследования трахеид и либриформа с помощью поляризационного и электронного микроскопа, а также рентгенографии позволили установить существование в клеточных стенках пяти концентрических слоев [39]: наружной, или первичной, стенки и вторичной стенки. Вторичная стенка в свою очередь разделяется на три слоя, обычно обозначаемых 81, вг и Бз. Кроме того, между первичными стенками соседних клеток располагается склеивающая их срединная пластинка (рис. 35).[ …]
Повышение выходов при использовании водяного пара объясняется тем, что ускоряется вынос ценных продуктов из реакционного пространства и задерживается развитие реакций вторичного распада. Кроме того, при соприкосновении водяного пара с капиллярной системой древесины на поверхностных слоях ее возможна конденсация пара, что создает условия для термического разложения в кислой водной среде. При этом реакции разложения происходят в первую очередь в слоях клеточной стенки, которые расположены с внутренних сторон клеточных полостей и состоят преимущественно из нетермостойких гемицеллюлоз, легко отщепляющих ацетильные группы и часть связанных с ними метоксилов, образуя соответственно уксусную кислоту и метиловый спирт.[ …]
Вряд ли правильно называть клетками сегменты, слагающие нити сфероплеи, и не только потому, что они обладают множеством ядер и хлоропластов (и, следовательно, являются явно вторичными образованиями), но и потому, что отделяющие их поперечные перегородки не похожи на клеточные стенки других многоклеточных зеленых водорослей. Они сильно варьируют по форме, а также по способу и месту образования (рис. 226, 4—6). Часто поперечные перегородки имеют вид кольцевых внутренних утолщений на стенках клетки, которые не смыкаются в центре, так что остается отверстие, через которое проходит цитоплазматический тяж (рис. 226, 4). В других случаях вместо перегородок образуются особые пробки. И, наконец, в любом месте нити могут возникать группы радиально сходящихся тяжей, напоминающих скелетные тяжи каулерпы и играющих механическую роль.[ …]
Снаружи от плазматической мембраны их клеток нет дополнительной плотной клеточной стенки или она состоит из хитина, редко из целлюлозы. Запасные углеводы обычно в форме гликогена (животного крахмала).[ …]
Маркс-Фигипи и Пепцел [12] изучали изменение СП хлопковой целлюлозы на различных стадиях созревания хлопка. Они показали, что вязкость растворов хлопковой целлюлозы снижается через несколько часов после открытия коробочки. Целлюлоза вторичной клеточной стенки в волокнах нераскрывшихся коробочек хлопка при небольшой зрелости (выход целлюлозы—18%) имеет единственный максимум на кривой распределения при СП 14 000. Около 10% материала имеет более низкий молекулярный вес (СП 1500—2500), эта целлюлоза содержится в первичной клеточной стенке.[ …]
Положение мест образования микрофибрилл но отношению к поверхности мембраны цитоплазмы может быть различно. Так, у бактерий этот процесс протекает в среде, значительно удаленной от поверхности клетки [28, 29] и, следовательно, от мембраны. По-видимому, аналогичным образом синтез протекает и в утолщенных первичных стенках клеток эпидермиса колеоптилей овса, поскольку синтез целлюлозы в этом случае осуществляется равномерно но толщине клеточной стенки [30]. В оболочках асцидий отложение целлюлозы происходит, по-видимому, также в местах, удаленных от поверхности секреторных клеток [24], хотя достаточно убедительных доказательств этого предположения нет. Напротив, микрофибриллы вторичных стенок клеток растений, возможно, образуются на внутренней поверхности стенки, в непосредственной близости от мембраны цитоплазмы [4, 27]. Поскольку целлюлозы во вторичных стенках значительно больше, чем в первичных, можно сделать вывод, что большинство целлюлозных микрофибрилл образуется вблизи мембраны цитоплазмы [4]. Однако это не является обязательным.[ …]
Одним из методов, основанных на этом принципе, является метод определения реакционной способности целлюлозы по картине набухания ксантогенатов в изо-пропилОвом спирте. Процесс набухания при взаимодействии волокна с растворителем схематически можно представить следующим образом: жидкость проникает внутрь волокна, вследствие чего объем волокна увеличивается. Затем происходит разрыв слабого эластичного наружного слоя вторичной клеточной стенки волокна и в местах разрыва образуются вздутия («бусы»). Остатки этого слоя образуют на набухшем волокне перетяжки и манжеты. Затем наружный слой отделяется и волокно равномерно набухает, на нем образуются поперечные полосы и волокно делится на пакеты дисков и отдельные диски, которые в дальнейшем растворяются.[ …]
Зависимость прочности древесины от содержания влаги. Так как прочность и жесткость древесины частично определяются силами сцепления, связывающими молекулы, то любой агент, уменьшающий эти силы, меняет ее прочность в целом. Одним из таких агентов является вода, поэтому прочность древесины увеличивается по мере уменьшения содержания влаги не только в результате повышенной плотности, происшедшей от усушки, но также из-за присутствия вторичных валентных сил сцепления1. Так как присутствие воды в количестве, превышающем точку насыщения волокна, не изменяет характера клеточной стенки, то потеря или приобретение капиллярной (свободной) воды практически не влияет на показатели прочности древесины.[ …]
Структуры, содержащие много лигнина, окрашиваются в темно-коричневый цвет до черного, тогда как слабо лигнифицированные зоны окрашиваются в светло-желтый цвет до янтарного. Результаты этой цветной реакции полностью подтверждают предшествующие работы по исследованию химии клеточной стенки. Вторичные стенки волокнистых элементов у древесины лиственных пород, растущих в умеренном климате, более светлые, следовательно, они менее лигнифицированы, чем вторичные стенки хвойных пород. Стенки сосудов у лиственных пород окрашены в более темный цвет, чем окружающие волокнистые элементы, следовательно, они содержат больше лигнина; мембраны пор также сильно лигнифицированы [84].[ …]
Эта операция осуществлялась на одревесневших срезах, предварительно освобожденных от лигнина с помощью хлорита натрия в уксуснокислой среде. Затем срезы были обработаны п-фенилаз-; бензоилхлоридом с целью этерификации полисахаридов. Ярко окрашенные в оранжево-красный цвет срезы после набухания в пиридине фотометрировались. Подвергая такой обработке срезы, со стоящие из холоцеллюлозы, до и после удаления гемицеллюлоз, удалось установить, что основная масса гемицеллюлоз в древесине ели и березы сосредоточена в наружных слоях вторичной стенки. Так, при экстракции среза еловой холоцеллюлозы 16%-ньш едким натром было установлено, что из наружных слоев клетки извлекается до 60—80%, из средины клеточной стенки около 50% и из слоя Бз только 16% растворимых в щелочи гемицеллюлоз от общего количества полисахаридов. Аналогичная картина наблюдалась и для поперечных срезов либриформа из древесины березы.[ …]
Опыты Риттера [26], а позднее Бейли [27] и др. показали, что независимо от возможного присутствия пектиновых полиуронидов в срединной пластинке,она состоит главным образом из лигнина, как его понимают химики (нерастворим в холодной 72%-ной серной кислоте, растворим после хлорирования и обработки слабыми основаниями или основными солями). Кроме того, Риттер доказал, что большая часть лигнина находится именно в этом слое. Это утверждение противоречило преобладавшему в то время мнению о присутствии большей части лигнина в других слоях, особенно во вторичной стенке. Позднее было доказано [30], что в таких случаях кажущаяся широкой и объемистой вторичная стенка в действительности подобна паутине, которая после высыхания съеживается и превращается в разрозненные кусочки. Исли первичные стенки включены в сложную срединную пластинку, то весьма вероятно, что здесь находится и большая часть лигнина.[ …]
Кальциевые каналы обнаружены и в мембранах растительных клеток. Показана регуляция входа 45Са2+ микросомы, выделенные из колеоп-тилей кукурузы и гипокотилей тыквы, светом, ПУК и зависимость этой реакции от кальмодулина. Для функционирования потенциалзависимых Са2+-каналов (харовая водоросль Ыие11ор,ш) необходимо наличие М§2+. Состояние этих потенциалзависимых каналов контролируется системой ферментов, рейдирующих уровень цАМФ в клетке. Были также получены данные, свидетельствующие о прямом действии экзогенного цАМФ на поглощение 45Са2+ в клетках СМатуёотопт гетскагсШ (мутант без клеточной стенки). Данные, приведенные на рис. 4.1, свидетельствуют о регуляторном действии цАМФ на поглощение Са2+ клетками. Это указывает на возможность взаиморегуляции двух систем вторичных посредников — цАМФ и Са2+. В опытах с животными клетками усиление поглощения Са2+ под действием цАМФ объясняется фосфорилированием белков потенциалзависимых Са2+-каналов и вследствие этого увеличением пребывания их в открытом состоянии.[ …]
Изучению действия ультразвука на целлюлозные волокна посвящено много исследований [447—459]. Некоторые исследователи сопоставляли или сочетали влияние ультразвука с различными механическими воздействиями. Так, Яйме, Кронерт и Нейхауз [447] изучали действие ультразвука на целлюлозные волокна по сравнению с высокочастотными механическими колебаниями и показали, что ультразвук с частотой 20—3000 кгц разрыхляет структуру волокна, увеличивает степень его набухания и обезвоживания. Механическая прочность бумаги, изготовленной из таких целлюлоз, повышается, особенно прочность к раздиранию. Аналогично действуют и высокочастотные механические колебания. Ивасаки, Линдберг и Мейер [448] считают, что общая картина изменений структуры волокна под действием ультразвука в водной среде сходна с изменениями структуры волокон при механическом размоле. При этом происходят глубокие изменения морфологической структуры волокон, приводящие к сдвигам во вторичной клеточной стенке, отрыву крупных кусков от первичной стенки, затем к набуханию вторичной стенки и ее дефибриллированию. В работе Сафоновой и Клен-ковой [449, 450] при изучении микрофотографий волокон, подвергнутых ультразвуковому воздействию в воде, показано, что имеются и другие, более глубокие нарушения в структуре волокна, которое становится пронизанным целой сетью многочисленных поперечных каналов. Отмечается [457], что волокна ранней древесины и волокна, не подвергавшиеся высушиванию, более восприимчивы к действию ультразвука.[ …]
Источник: ru-ecology.info
Общие черты строения растительной клетки.
1. 1. Клеточная стенка, которой окружен каждый протопласт – живое содержимое клетки — служит скелетом растения. Протопласт заключен в ней как в жестком футляре, а это делает возможным увеличение тургорного давления.
Клеточная стенка, которая образуется во время деления клетки, называется первичной клеточной стенкой. Она состоит из целлюлозных микрофибрилл (волокон), погруженных в матрикс. Матрикс на 80-60% состоит из целлюлозных волокон и на 20-40 % из пектиновых веществ.. (рис. 6).
Целлюлоза – полимер, мономером которого является глюкоза. Одна молекула целлюлозы состоит из 10000 остатков глюкозы, соединенных в единую цепочку. Целлюлозные цепочки соединяются друг с другом и образуют пучки или волокна, которые называются микрофибриллами. Соединение целлюлозных молекул в микрофибриллы происходит за счет образования водородных связей между гидроксильными группами соседних цепей.
Микрофибриллы располагаютсмя слоями в матриксе и образуют каркас клеточной стенки. В первичной клеточной стенке целлюлозных волокон мало.
Матрикс – мягкая пластичная масса, состоящая из 60-70% из воды и 40-30 % гемицеллюлоз и пектиновых веществ.
Гемицеллюлозы – полимеры, мономерами которых являются пентозы и гексозы. Гемицеллюлозные цепочки короче целлюлозных цепочек и более разветвлены.
Пектиновые вещества – полимеры, мономерами которых являются сахароподобные единицы. Они образуют линейные или разветвленные цепи.
В матриксе клеточной стенки также могу откладываться минеральные вещества (оксид кремния, карбонат кальция), которые также придают прочность клеточной стенке.
У многих растений, когда клетка заканчивает свой рост, на внутренней поверхности первичной клеточной стенки отлагаются дополнительные слои целлюлозных волокон и образуется вторичная клеточная стенка. Целлюлозные волокна в ней располагаются под прямым углом друг к другу, что придает вторичной клеточной стенке большую жесткость и прочность.
В некоторых клетках после прекращения роста утолщение вторичных клеточных стенок приводит к уменьшению объема протопласта. А в волокнах и трахеидах (механическая и проводящая ткань) клеточная стенка может заполнять почти всю клетку. Протопласт отмирает и остаются только клеточные стенки в виде полых цилиндров, которые выполняют функцию опоры или функцию проведения веществ.
В таких клетках на поверхности первичных клеточных стенок и в промежутках между целлюлозными волокнами откладывается лигнин, который пропитывает и первичную, и вторичную клеточные стенки. Лигнин – сложное полимерное вещество, которое скрепляет целлюлозные волокна и удерживает их на месте. Лигнин обеспечивает клеткам дополнительную защиту от неблагоприятных физических и химических воздействий среды.
Клеточные стенки тканей, образующих кору, обычно пропитываются суберином. Это водонепроницаемое вещество, которое приводит к опробковению тканей.
Клеточные стенки эпидермиса листа пропитываются воскообразным веществом кутином. Он предохраняет ткани листа от излишней потери воды.
Образование клеточной стенки происходит следующим образом. После окончания деления клетки откладывается первый слой клеточной стенки – срединная пластинка. Она состоит сначала из пектиновых веществ, позднее в ней появляются целлюлозные волокна, и она пропитывается суберином придающим клеточной стенке особую прочность.
По мере того, как клетка растет и ее объем увеличивается, откладывается следующий слой – первичная клеточная стенка. Рост первичной клеточной стенки происходит за счет образования новых молекул целлюлозы. Первичная клеточная стенка формируется по обе стороны от срединной пластинки, образуя два слоя.
Позднее, за счет утолщения первичной клеточной стенки образуется вторичная клеточная стенка. Вторичная клеточная стенка придает растительной клетке окончательную форму.
В образовании клеточной стенки принимают участие аппарат Гольджи и плазмалемма. Вещества матрикса гемицеллюлозы и пектиновые вещества синтезируются в аппарате Гольджи. и выделяются через плазмалемму путем экзоцитоза по всей поверхности клетки. Молекулы целлюлозы синтезируются специальными ферментами, встроенными в плазмалемму.
2. 2. Вакуоль. Это мембранный мешочек, заполненный водным раствором органических веществ и минеральных солей. Молодые клетки содержат несколько мелких вакуолей, которые в зрелых клетках сливаются в одну крупную центральную вакуоль. Центральная вакуоль в клетке может занимать до 80-90 % ее объема. От цитоплазмы центральная вакуоль отделена мембраной, которая называется тонопласт, обладающий избирательной проницаемостью.
Жидкость, заполняющая центральную вакуоль, называется клеточным соком. Это коллоидный раствор, который содержит минеральные соли, сахара, органические кислоты, кислород, углекислый газ. В клеточном соке содержатся пигменты антоцианы, которые придают цветкам, листьям, плодам синюю, голубую, фиолетовую окраску. Центральная вакуоль служит местом хранения некоторых продуктов обмена веществ клетки, например, оксалата кальция, токсичного для клеток. В клеточном соке содержатся биологически активные вещества растения (алкалоиды, фенолы, дубильные вещества, таннины). Они определяют фармакологическую ценность растений, а также служат защитой от поедания животными. У некоторых растений в клеточном соке содержится млечный сок — латекс (одуванчик, чистотел, гевея).
Центральная вакуоль выполняет несколько функций:
1. 1. Центральная вакуоль поддерживает тургорное состояние клетки. Вода поступает в концентрированный клеточный сок путем осмоса через тонопласт. В клетке развивается тургорное давление и цитоплазма прижимается к клеточной стенке, что придает форму растительной клетке.
2. 2. Через тонопласт, обладающий избирательной проницаемостью, осуществляется активный транспорт веществ. Поэтому в вакуолях накапливаются различные вещества:
а) в вакуоли накапливаются отходы жизнедеятельности клетки (оксалат кальция);
б) в вакуоли хранятся запасные вещества (сахара, белки в алейроновых зернах семян)
в) вещества, выделяемые из клетки, также находятся в вакуоли (таннины, дубильные вещества алкалоиды, латекс).
3. 3. Пластиды.
Как уже упоминалось ранее к пластидам относятся хлоропласты, хромопласты и лейкопласты.
Источник: www.bsu.ru
Клеточная стенка (клеточная оболочка) – характерный признак растительной клетки, отличающий ее от клетки животной. Клеточная стенка придает клетке определенную форму. Культивируемые на специальных питательных средах клетки растений, у которых ферментативным путем удаляется стенка, всегда принимают сферическую форму. Клеточная стенка придает клетке прочность и защищает протопласт, она уравновешивает тургорное давление и препятствует, таким образом, разрыву плазмалеммы. Совокупность клеточных стенок образует внутренний скелет, поддерживающий тело растения и придающий ему механическую прочность.
Клеточная стенка бесцветна и прозрачна, легко пропускает солнечный свет. Обычно стенки пропитаны водой. По системе клеточных стенок осуществляется транспорт воды и растворенных в ней низкомолекулярных соединений (транспорт по апопласту).
Клеточная стенка состоит в основном из полисахаридов, которые можно подразделить на скелетные вещества и вещества матрикса.
Скелетным веществом клеточной стенки растений является целлюлоза (клетчатка), представляющая собой бета-1,4-D-глюкан. Это самое распространенное органическое вещество биосферы. Молекулы целлюлозы представляют собой очень длинные неразветвленные цепи, они располагаются параллельно друг другу группами по нескольку десятков и скреплены многочисленными водородными связями. В результате образуются микрофибриллы, которые создают структурный каркас стенки и обусловливают ее прочность. Микрофибриллы целлюлозы видны только в электронный микроскоп, их диаметр равен 10-30 нм, длина достигает нескольких мкм.
Целлюлоза нерастворима и не набухает в воде. Она очень инертна в химическом отношении, не растворяется в органических растворителях, концентрированных щелочах и разведенных кислотах. Микрофибриллы целлюлозы эластичны и очень прочны на разрыв (сходны со сталью). Эти свойства определяют широкое применение целлюлозы и ее продуктов. Мировая продукция хлопкового волокна, состоящего почти целиком из целлюлозы, составляет 1,5•107 тонн в год. Из целлюлозы получают бездымный порох, ацетатный шелк и вискозу, целлофан, бумагу. Качественную реакцию на целлюлозу проводят с реактивом хлор-цинк-йод, целлюлозная клеточная стенка окрашивается в сине-фиолетовый цвет.
У грибов скелетным веществом клеточной стенки является хитин – полисахарид, построенный из остатков глюкозамина. Хитин еще более прочен, чем целлюлоза.
Микрофибриллы погружены в аморфный матрикс, обычно представляющий собой насыщенный водой пластичный гель. Матрикс является сложной смесью полисахаридов, молекулы которых состоят из остатков нескольких различных сахаров и представляют собой более короткие, чем у целлюлозы, и разветвленные цепи. Матричные полисахариды определяют такие свойства клеточной стенки, как сильная набухаемость, высокая проницаемость для воды и растворенных в ней низкомолекулярных соединений, катионообменные свойства. Полисахариды матрикса делят на две группы – пектиновые вещества и гемицеллюлозы.
Пектиновые вещества сильно набухают или растворяются в воде. Они легко разрушаются под действием щелочей и кислот. Простейшими представителями пектиновых веществ являются растворимые в воде пектовые кислоты – продукты полимеризации альфа-D-галактуроновой кислоты (до 100 единиц), связанных 1,4-связями в линейные цепи (альфа-1,4-D-галактуронан). Пектиновые кислоты (пектины) – это более высокомолекулярные (100-200 единиц) полимерные соединения альфа-D-галактуроновой кислоты, в которых карбоксильные группы частично метилированы. Пектаты и пектинаты – кальциевые и магниевые соли пектовых и пектиновых кислот. Пектиновые кислоты, пектаты и пектинаты растворимы в воде в присутствии сахаров и органических кислот с образованием плотных гелей.
В клеточных стенках растений в основном присутствуют протопектины – высокомолекулярные полимеры метоксилированной полигалактуроновой кислоты с арабинанами и галактанами, у двудольных растений в состав цепей галактуронана входит небольшое количество рамнозы. Протопектины нерастворимы в воде.
Гемицеллюлозы представляют собой разветвленные цепи, построенные из остатков нейтральных сахаров, чаще встречаются глюкоза, галактоза, манноза, ксилоза; степень полимеризации 50-300. Гемицеллюлозы химически более устойчивы, чем пектиновые вещества, они труднее гидролизуются и слабее набухают в воде. Гемицеллюлозы могут откладываться в стенках клеток семян в качестве запасных веществ (финиковая пальма, хурма). Пектиновые вещества и гемицеллюлозы связаны взаимными переходами. Помимо полисахаридов, в матриксе клеточных стенок присутствует особый структурный белок. Он связан с остатками сахара арабинозы и поэтому является гликопротеидом.
Матричные полисахариды не просто заполняют промежутки между целлюлозными микрофибриллами. Их цепи располагаются упорядоченно и образуют многочисленные связи как друг с другом, так и с микрофибриллами, что значительно повышает прочность клеточной стенки.
Клеточные стенки растений часто подвергаются химическим видоизменениям. Одревеснение, или лигнификация происходит в том случае, если в матриксе откладывается лигнин – полимерное соединение фенольной природы, нерастворимое в воде. Одревесневшая клеточная стенка теряет эластичность, резко повышается ее твердость и прочность на сжатие, снижается проницаемость для воды. Реактивами на лигнин являются: 1) флороглюцин и концентрированная хлористоводородная или серная кислота (одревесневшие стенки приобретают вишнево-красную окраску) и 2) сульфат анилина, под действием которого одревесневшие стенки становятся лимонно-желтыми. Лигнификация характерна для стенок клеток проводящей ткани ксилемы (древесины) и механической ткани склеренхимы.
Опробковение, или суберинизация происходит в результате отложения с внутренней стороны клеточной стенки гидрофобных полимеров – суберина и воска. Суберин представляет собой смесь эфиров полимерных жирных кислот. Мономерами воска являются жирные спирты и восковые эфиры. Воск легко извлекается органическими растворителями и быстро плавится, образует кристаллы. Суберин – аморфное соединение, не плавится и не растворяется в органических растворителях. Суберин и воск, образуя чередующиеся параллельные слои, выстилают всю полость клетки с внутренней стороны в виде пленки. Субериновая пленка практически непроницаема для воды и для газов, поэтому после ее образования клетка обычно отмирает. Опробковение характерно для стенок клеток покровной ткани пробки. Реактивом на опробковевшую клеточную стенку является судан III, окраска оранжево-красная.
Кутинизации подвергаются наружные стенки клеток покровной ткани эпидермы. Кутин и воск откладываются чередующимися слоями на наружной поверхности клеточной стенки в виде пленки – кутикулы. Кутин представляет собой жироподобное полимерное соединение, близкое по химической природе и свойствам суберину. Кутикула предохраняет растение от излишнего испарения воды с поверхности растения. Окрасить ее можно реактивом судан III в оранжево-красный цвет.
Минерализация клеточной стенки происходит вследствие отложения в матриксе большого количества минеральных веществ, чаще всего кремнезема (оксида кремния), реже оксалата и карбоната кальция. Минеральные вещества придают стенке твердость и хрупкость. Отложение кремнезема характерно для клеток эпидермы хвощей, осок и злаков. Приобретенная в результате окремнения жесткость стеблей и листьев служит защитным средством против улиток, а также значительно снижает поедаемость и кормовую ценность растений.
У некоторых специализированных клеток наблюдается ослизнение клеточной стенки. При этом вместо целлюлозной вторичной стенки происходит отложение аморфных, сильно гидратированных кислых полисахаридов в виде слизей и камедей, близких по химической природе к пектиновым веществам. Слизи хорошо растворяются в воде с образованием слизистых растворов. Камеди клейкие, вытягиваются в нити. В сухом виде они имеют роговую консистенцию. При отложении слизи протопласт постепенно оттесняется к центру клетки, его объем и объем вакуоли постепенно уменьшаются. В конце концов, полость клетки может целиком заполниться слизью, и клетка отмирает. В некоторых случаях слизь может проходить через первичную клеточную стенку на поверхность. В синтезе и секреции слизи основное участие принимает аппарат Гольджи.
Выделяемая растительными клетками слизь выполняет различные функции. Так, слизь корневого чехлика служит в качестве смазки, облегчающей рост кончика корня в почве. Слизевые железки насекомоядных растений (росянка) выделяют ловчую слизь, к которой приклеиваются насекомые. Слизь, выделяемая наружными клетками семенной кожуры (лен, айва, подорожники), закрепляет семя на поверхности почвы и защищает проросток от высыхания. Слизь окрашивается реактивом метиленовый синий в голубой цвет.
Выделение камедей обычно происходит при поранении растений. Например, камедетечение из пораненных участков стволов и ветвей часто наблюдается у вишни и сливы. Вишневый клей представляет собой застывшую камедь. Камедь выполняет защитную функцию, закрывая рану с поверхности. Образуются камеди в основном у древесных растений из семейств бобовых (акации, трагакантовые астрагалы) и розоцветных подсемейства сливовых (вишня, слива, абрикос). Камеди и слизи используются в медицине.
Клеточная стенка является продуктом жизнедеятельности протопласта. Полисахариды матрикса, гликопротеид стенки, лигнин и слизи образуются в аппарате Гольджи. Синтез целлюлозы, образование и ориентация микрофибрилл осуществляются плазмалеммой. Большая роль в ориентации микрофибрилл принадлежит микротрубочкам, которые располагаются параллельно откладывающимся микрофибриллам вблизи плазмалеммы. Если микротрубочки разрушить, образуются только изодиаметрические клетки.
Образование клеточной стенки начинается во время деления клетки. В плоскости деления образуется клеточная пластинка, единый слой, общий для двух дочерних клеток. Она состоит из пектиновых веществ, имеющих полужидкую консистенцию; целлюлоза отсутствует. Во взрослой клетке клеточная пластинка сохраняется, но претерпевает изменения, поэтому ее называют срединной, или межклеточной пластинкой (межклеточным веществом) (рис. 2.16 ). Срединная пластинка обычно очень тонка и почти неразличима.
Сразу после образования клеточной пластинки протопласты дочерних клеток начинают откладывать собственную клеточную стенку. Она откладывается изнутри как на поверхности клеточной пластинки, так и на поверхности других клеточных стенок, принадлежавших ранее материнской клетке. После деления клетка вступает в фазу роста растяжением, который обусловлен интенсивным осмотическим поглощением клеткой воды, связанным с образованием и ростом центральной вакуоли. Тургорное давление начинает растягивать стенку, но она не рвется благодаря тому, что в нее постоянно откладываются новые порции микрофибрилл и веществ матрикса. Отложение новых порций материала происходит равномерно по всей поверхности протопласта, поэтому толщина клеточной стенки не уменьшается.
Стенки делящихся и растущих клеток называют первичными. Они содержат много (60-90%) воды. В сухом веществе преобладают матричные полисахариды (60-70%), содержание целлюлозы не превышает 30%, лигнин отсутствует. Толщина первичной стенки очень невелика (0,1-0,5 мкм).
Для многих клеток отложение клеточной стенки прекращается одновременно с прекращением роста клетки. Такие клетки окружены тонкой первичной стенкой до конца жизни ( рис. 2.16).

Рис. 2.16. Паренхимная клетка с первичной стенкой.
У других клеток отложение стенки продолжается и по достижении клеткой окончательного размера. При этом толщина стенки увеличивается, а объем, занимаемый полостью клетки, сокращается. Такой процесс носит название вторичного утолщения стенки, а саму стенку называют вторичной (рис. 2.17 ).
Вторичная стенка может рассматриваться как дополнительная, выполняющая главным образом механическую, опорную функцию. Именно вторичная стенка ответственна за свойства древесины, текстильного волокна, бумаги. Вторичная стенка содержит значительно меньше воды, чем первичная; в ней преобладают микрофибриллы целлюлозы (40-50% от массы сухого вещества), которые располагаются параллельно друг другу. Из полисахаридов матрикса характерны гемицеллюлозы (20-30%), пектиновых веществ очень мало. Вторичные клеточные стенки, как правило, подвергаются одревеснению. В неодревесневших вторичных стенках (лубяные волокна льна, волоски хлопчатника) содержание целлюлозы может достигать 95%. Большое содержание и строго упорядоченная ориентация микрофибрилл определяют высокие механические свойства вторичных стенок. Часто клетки, имеющие вторичную одревесневшую клеточную стенку, после завершения вторичного утолщения отмирают.
Срединная пластинка склеивает соседние клетки. Если ее растворить, стенки клеток теряют связь друг с другом и разъединяются. Этот процесс называется мацерация. Довольно обычна естественная мацерация, при которой пектиновые вещества срединной пластинки переводятся в растворимое состояние с помощью фермента пектиназы и затем вымываются водой (перезрелые плоды груши, дыни, персика, банана). Часто наблюдается частичная мацерация, при которой срединная пластинка растворяется не по всей поверхности, а лишь в углах клеток. Вследствие тургорного давления соседние клетки в этих местах округляются, в результате чего образуются межклетники (рис. 2.16 ). Межклетники образуют единую разветвленную сеть, которая заполняется парами воды и газами. Таким образом, межклетники улучшают газообмен клеток.
Характерная особенность вторичной стенки – ее неравномерное отложение поверх первичной стенки, в результате чего во вторичной стенке остаются неутолщенные участки – поры. Если вторичная стенка не достигает большой толщины, поры выглядят как мелкие углубления. У клеток с мощной вторичной стенкой поры в разрезе имеют вид радиальных каналов, идущих от полости клетки до первичной стенки. По форме порового канала различают поры двух типов – простые и окаймленные (рис. 2.17 ).
Рис. 2.17. Типы пор : А – клетки с вторичными стенками и многочисленными простыми порами; Б – пара простых пор; В – пара окаймленных пор.
У простых пор диаметр порового канала по всей длине одинаковый и имеет форму узкого цилиндра. Простые поры характерны для паренхимных клеток, лубяных и древесинных волокон.
Поры в двух смежных клетках, как правило, возникают друг против друга. Эти общие поры имеют вид одного канала, разделенного тонкой перегородкой из срединной пластинки и первичной стенки. Такая совокупность двух пор смежных стенок соседних клеток носит название пары пор и функционирует как одно целое. Разделяющий их канал участок стенки называется замыкающей пленкой поры, или поровой мембраной. В живых клетках замыкающая пленка поры пронизана многочисленными плазмодесмами (рис. 2.18 ).
Плазмодесмы присущи только растительным клеткам. Они представляют собой тяжи цитоплазмы, пересекающие стенку смежных клеток. Число плазмодесм в одной клетке очень велико – от нескольких сотен до десятков тысяч, обычно плазмодесмы собраны в группы. Диаметр плазмодесменного канала составляет 30-60 нм. Его стенки выстланы плазмалеммой, непрерывной с плазмалеммой смежных клеток. В центре плазмодесмы проходит мембранный цилиндр – центральный стержень плазмодесмы, непрерывный с мембранами элементов эндоплазматической сети обеих клеток. Между центральным стержнем и плазмалеммой в канале находится гиалоплазма, непрерывная с гиалоплазмой смежных клеток.
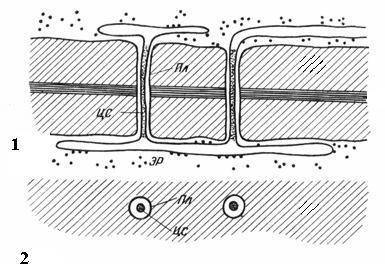
Рис. 2.18. Плазмодесмы под электронным микроскопом (схема ): 1 – на продольном срезе; 2 – на поперечном срезе; Пл – плазмалемма; ЦС – центральный стержень плазмодесмы; ЭР – элемент эндоплазматического ретикулума.
Таким образом, протопласты клеток не полностью изолированы друг от друга, а сообщаются по каналам плазмодесм. По ним происходит межклеточный транспорт ионов и мелких молекул, а также передаются гормональные стимулы. Посредством плазмодесм протопласты клеток в растительном организме образуют единое целое, называемое симпластом, а транспорт веществ через плазмодесмы получил название симпластического в отличие от апопластического транспорта по клеточным стенкам и межклетникам.
У окаймленных пор (рис. 2.17 ) канал резко суживается в процессе отложения клеточной стенки, поэтому внутреннее отверстие поры, выходящее в полость клетки, гораздо уже, чем наружное, упирающееся в первичную стенку. Окаймленные поры характерны для рано отмирающих клеток водопроводящих элементов древесины. У них поровый канал по направлению к замыкающей пленке воронковидно расширяется, а вторичная стенка нависает в виде валика над расширенной частью канала, образуя камеру поры. Название окаймленной поры происходит оттого, что при рассмотрении с поверхности внутреннее отверстие имеет вид маленького круга или узкой щели, тогда как наружное отверстие как бы окаймляет внутреннее в виде круга большего диаметра или более широкой щели.
Поры облегчают транспорт воды и растворенных веществ от клетки к клетке, в то же время не снижая прочности клеточной стенки.
Источник: StudFiles.net
Клеточные стенки высших растений
Растительные клеточные стенки выполняют целый ряд функций: они обеспечивают жесткость клетки для структурной и механической поддержки, придают форму клетке, направление ее роста и в конечном счете морфологию всему растению. Клеточная стенка также противодействует тургора, то есть осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают против патогенов, проникающих из окружающей среды, и запасают углеводы для растения. Растительные клеточные стенки строятся прежде всего с углеводного полимера целлюлозы.
В течение цитокинеза сначала с клеточной пластинки формируется промежуточная ламела, и первичная клеточная стенка расширяется внутри промежуточной ламели. Фактическая структура клеточной стенки не всегда четко определена, существуют несколько моделей ее структуры: модель перекрестных ковелентних связей, модель привязи, модель диффузного слоя и модель стратифицированного слоя. Однако, первичная клеточная стенка может быть определена как сеть волокон целлюлозы направленных в случайных направлениях. Волокна воздержатся вместе водородными связями, обеспечивает достаточно высокую прочность. Клетки держатся вместе и разделяют желатиновую мембрану, называет его промежуточная ламела, которая содержит пектаты (соли пектиновой кислоты) магния и кальция. Клетки взаимодействуют через плазмодесмы, то есть цитоплазмени канальцы, соединяющие цитоплазмы клеток через клеточные стенки.
В некоторых типах клеток некоторых растений, после того как достигается максимальный размер или определенная точка развития, между растительным клеткой и первичной стенкой формируется вторичная стенка. В отличие от первичной стенки, волокна выравниваются в основном в одном направлении, и с каждым дополнительным слоем их ориентация слегка меняется. Клетки с вторичными клеточными стенками очень жесткие. Межклеточное взаимодействие все еще возможна через отверстия во вторичной клеточной стенке, позволяющие плазмодкрми проникать через обе клеточные стенки.
Основные углеводы, составляющих первичную клеточную стенку, это целлюлоза, гемицеллюлоза и пектин. Целлюлозные микроволокна связываются через хемицелюлозни мостики, формируя целлюлозно-хемицелюлозну сеть, которая окружена матрицей пектина. Самый общий тип хемицелюлозы в первичной клеточной стенке — ксилоглюкан.
Растительные клеточные стенки также содержат ряд белков, распространенные включают богатые гидропролин гликопротеины (HRGP), также известные как екстенсины, арабиногалактан протеины (AGP), богатые глицин протеины (GRP) и богатые пролин протеины (PRP). За исключением богатых глицин протеинов, все эти белки гликозилированные и содержат гидроксипролин (Hyp). Каждый класс гликопротеинов определяется характерной, часто повторяющейся последовательности аминокислот. Причудливые протеины содержат два или более областей, каждая из последовательностью от другого класса гликопротеинов. Большинство белков клеточной стенки перекрестных связаны с углеводами клеточной стенки и, возможно, имеют структурные функции.
Вторичные клеточные стенки могут содержать лигнин и суберин, делая стенки твердыми. Относительный состав углеводов, дополнительных компонентов и белков зависит от вида растения, типа и возраста клеток.
Клеточные стенки водорослей
Подобно высших растений, большинство водорослей, независимо от происхождения, имеют клеточные стенки. Клеточные стенки водорослей содержат целлюлозу и многие другие гликопротеинов. Дополнительные полисахариды в клеточных стенках водорослей раньше часто использовали как один из признаков для установления их таксономии, но некоторые несвязанные группы имеют те же полисахариды.
- Манозилови микроволокна зучтричаються в клеточных стенках многих зеленых водорослей (примитивних растений), в том числе родов Codium, Dasycladus и Acetabularia, и некоторых красных водорослей например, родов Porphyra и Bangia.
- Ксилан
- Альгиновая кислота — обычный полисахарид в клеточных стенках бурых водорослей
- Сульфоновани ролисахариды встречаются клеточных стенках большинства водорослей, например, в красных водорослях находим агарозу, каррагинан, порфиран, фурселеран и фуноран.
Другие вещества, которые могут быть найдены в водорослевых клеточных стенках, включают спорополенин и кальций.
Группа диатомовых водорослей (диатомей) синтезируют клеточные стенки (также известно как фрустулы или клапаны) с Кремнев кислоты (особенно ортокремневои кислоты, H 4 SiO 4). Кислота полимеризуется внутриклеточно, затем стенка вытесняется наружу для защиты клетки. Такие стенки требуют меньше энергии на свой синтез, позволяя сохранить много энергии, что, возможно, частично отвечает за быстрый рост этих организмов.
Клеточные стенки грибов
Не все виды грибов имеют клеточные стенки, но если стенка присутствует, она состоит из глюкозамина и хитина, того же углерода, что придает прочность экзоскелета насекомых. Они служат подобной цели, что и клеточные стенки растений, предоставляя грибным клеткам твердость и поддерживать свою форму и предотвращения осмотическому лизису. Они также ограничивают вход молекул, потенциально ядовитых для гриба, например некоторым фунгициды растительного и искусственного происхождения. Состав, свойства и форма грибной клеточной стенки меняются на протяжении жизненного цикла и зависят от условий произрастания.
Другие эукариоты
Оомицеты, патогены растений похожи на грибы, также имеют целлюлозные клеточные стенки. До недавнего времени за этот признак их относили к грибам, структурные и молекулярные свидетельства заставили отнести их к гетероконтив, как и бурые водоросли.
Клеточные стенки бактерий
Как в других организмах, бактериальные клеточные стенки обеспечивают структурную целостность клеткам. У прокариот, первичная функция клеточной стенки — защитить ячейку от внутреннего тургора, вызванным высокой концентрацией белков и инших молекулы внутри клетки по сравнению с окружающей средой. Бактериальная клеточная стенка отличается от других тем, что ее основным компонентом является пептидогликан, слой которого размещается немедленно вне цитоплазматической мембраны. Пептидогликан отвечает за твердость стенки и придания формы клетке. Он относительно пористый и не мешает потока растворимых молекул сквозь него. Существует два основных типа бактериальных клеточных стенок, по этому признаку бактерии делятся на грам-отрицательные и грам-положительные.
Грам-положительные бактерии
Грам-положительные бактерии характеризуется присутствием очень толстого слоя пептидогликана, который отвечает за содержание красителя кристал-виолет течение процедуры окрашивания по Граму. Такая стенка находится исключительно в организмах, принадлежащих к типам Actinobacteria (или грамм-положительные организмы с высоким содержанием G + C) и Firmicutes (или грамм-положительные организмы с низким содержанием G + C). Бактерии в пределах типа Deinococcus-Thermus также могут положительно краситься по Граму, но содержат некоторые структуры клеточной стенки, типичные от грамотрицательных организмов. В клеточную стенку грамположительных бактерий встроенные полимерного спирты, тейхоевая кислота, некоторые из которых связываются с липидами, формируя липотейхоеву кислоту. Ци вещества отвечают за соединение пептидогликана с цитоплазматической мембраной. Тейхоевая кислота придает клетке отрицательный электрический заряд благодаря наличию фосфодиестерних связей между мономерами тейхоевая кислоты.
Грамотрицательные бактерии
В отличие от грамположительных бактерий, грамотрицательные бактерии содержат очень тонкий слой пептидогликана, отвечающий за неспособность клеточных стенок содержать краситель кристал-виолет течение процедуры окрашивания по Граму. В дополнение к слою пептидогликанов, грамотрицательные бактерии имеют вторую, так называемую внешнюю мембрану, находится кнаружи от клеточной стенки и компонует фосфолипиды и липополисахариды на своей внешней стороне. Отрицательно заряженные липополисахариды также предоставляют клетке отрицательный электрический заряд. Химическая структура липополисахаридив внешней мембраны часто уникальная для отдельных штаммов бактерий и часто отвечает за реакцию антигенов с представителями этих штаммов.
Как любой двойной слой фосфолипидов, внешняя мембрана достаточно непроницаема для всех заряженных молекул. Однако, белковые каналы (погрузится) присутствуют во внешней мембране, позволяют пассивный транспорт многих ионов, сахара и аминокислот через внешнюю мембрану. Таким образом, эти молекулы присутствуют в периплазматическое, слое между внешней и цитоплазматической мембранами. Периплазматическое содержит слой пептидогликана и много белков, шо отвечают за гидролиз и прием внеклеточных сигналов. Читается, что перивлазма гелеобразная, а не жидкая, из-за высокого содержания белка и пептидогликана. Сигналы и живильни вещества с периплазматическое попадают в цитоплазму клетки используя транспортные белки в цитоплизматичний мембране.
Клеточные стенки архей
Хотя они и не уникальны, клеточные стенки архей несколько отличаются от бактериальных. Например, клеточные стенки большинства архей образованные внешними слоями белков или S-слоем. S-слоя распространенные в бактериях, где они служат единственным компонентом клеточной стенки в некоторых организмах (например в Planctomyces) или внешним слоем во многих организмах с пептидогликана. За исключением одной группы метаногенов, археи не имеют пептидогликаннои стенки. Даже в данном случае, пептидогликаны очень отличается от типа, найденного в бактериях.
Источник: info-farm.ru
«Ботаника есть естественная наука, которая учит познанию растений». Такое определение ботаники — необходимое и достаточное — дано выдающимся шведским ученым Карлом Линнеем (1707-1778 гг.). В сферу ботаники входят изучение строения и функций растений, их происхождения, эволюции, классификации, взаимоотношений друг с другом и средой обитания, представления об образуемых растениями сообществах, расселении на Земном шаре, использовании и охране.
Конечно, уже первобытный человек обладал первоначальными знаниями о растениях, необходимых для его существования. Это понятно, поскольку его жизнь зависела от знаний о съедобных, ядовитых, целебных растениях и полезных для скота. Обширнейшими сведениями о растениях, особенно сельскохозяйственных и лекарственных, располагали культуры Индии, Финикии, страны древнего Египта и Месопотамии. Не случайно первый «травник на камне» был создан в знаменитом храме в Карнаке фараоном новой египетской династии Тутмосом III.
Но основы ботаники (от греч. botanicos — относящийся к растениям, botane — трава, растение) как научной дисциплины были заложены в античное время Теофрастом (371-286 гг. до н.э.) — любимым и выдающимся учеником великого древнегреческого мыслителя Аристотеля (384-322 гг. до н. э.). Титул «отца ботаники» Теофраст заслужил потому, что его интересовали не только применение растений в хозяйстве и медицине, он исследовал строение и физиологические отправления растений, их распространение, влияние на них почвы и климата. Теофрасту принадлежит и первая классификация растений, хотя и весьма наивная с позиций XX века.
В процессе исторического развития в ботанике появились разные методы изучения растений. Чем более расширялись представления о растениях, тем более дифференцировались научные дисциплины, составляющие ботанику как одну из самых разветвленных естественных наук: морфология в широком понимании, палеоботаника, физиология, биохимия растений, систематика, география, экология растений, геоботаника, палиноморфология, изучающая структуру пыльцевых зерен, и т.д. Особое место среди этих дисциплин занимала и занимает морфология (от греч. morphe — форма и logos — учение).
«Органическая форма — это видимое проявление внутренних связей, характеризующих жизнь на каждом уровне. Она может быть проще всего определена как биологическая организация и представляет собой наиболее важную проблему, с которой сталкиваются изучающие науку о жизни. Форму можно назвать не только душой естественной истории, так как она служит мерой эволюционного родства, но и душой всей биологии, так как она является очевидным и легко доступным изучению проявлением основных черт жизни».
По морфологическим признакам судят о разнообразии растений, они составляют основу их классификации; без знания структуры невозможно изучать жизненные отправления растений, в том числе их способность благодаря фотосинтезу создавать органические вещества и увеличивать содержание в атмосфере кислорода. Поэтому изучение структурных особенностей растений необходимо для развития других ботанических дисциплин.
Дифференциация методов исследования строения растений привела к разделению морфологии на многочисленные специальные дисциплины: морфологию в узком смысле слова (макроморфологию), изучающую внешнее строение растений; эмбриологию, изучающую начальные этапы развития семенных растений от заложения репродуктивных структур, осуществляющих размножение, до образования семени; анатомию, изучающую строение растений на клеточном и тканевом уровнях. Учение о клетке в настоящее время составляет содержание самостоятельной биологической дисциплины — цитологии.
Разнообразие методов, используемых в морфологии растений, позволяет решать следующие проблемы, нередко имеющие общебиологическое значение.
1. Изучение топографических закономерностей в строении растений. Главным методом исследования служит описательный, созданный К. Линнеем. Сейчас этот метод обычно называют сравнительно-морфологическим.
2. Изучение закономерностей формообразования (морфогенеза) в процессе индивидуального развития растения — его онтогенеза. Это требует изучения структурных преобразований растения на всех этапах его развития — от зиготы до естественной смерти. При этом важное значение имеет анализ всех проявлений морфогенеза: особенностей роста, морфологической и анатомической дифференциации тела растения, возникающих в процессе его развития, полярности, симметрии, корреляции. Естественно, глубина изучения этих вопросов зависит от тесных контактов морфологии с другими ботаническими дисциплинами: физиологией, генетикой, биохимией, биологией развития.
С этой проблемой связано и развитие репродуктивной биологии, основу которой составляет изучение всех структур и процессов, приводящих к размножению растений — одному из главных свойств всех живых организмов, обеспечивающему не только увеличение числа особей, но и их расселение. Большой интерес в настоящее время вызывает раздел репродуктивной биологии, непосредственно связанный с накоплением биомассы, — биотехнологией: культурой изолированных клеток и тканей как способа быстрого размножения растений.
3. Изучение морфогенетических трансформаций в течение длительного процесса эволюции. Развитие этого направления — эволюционной морфологии — основано на синтезе данных онтогенетической морфологии и сравнительной морфологии ныне живущих и вымерших растений. Задача эволюционной морфологии — изучение общих закономерностей преобразования структуры растений в процессе эволюции, без знания которых невозможно решение вопросов, связанных с филогенией растений, отражающей не только родственные отношения между разными таксонами, но и основные направления их эволюции. Таксонами (лат. taxon, во множественном числе taxa) называют любые конкретные систематические группы определенного ранга. Так, таксоном в ранге семейства будет семейство Ranunculaceae (лютиковые), в ранге рода — Ranunculus L. (лютик), а в ранге вида, например, Ranunculus repens L. (лютик ползучий).
О родственных связях прежде всего судят по сходству морфологических признаков. Однако нередко оно может быть не результатом родства, а либо параллельного развития нескольких групп растений от каких-то общих предков, либо следствием конвергенции — появлением сходных особенностей строения под влиянием одинаковых условий существования. Только разностороннее изучение растений и сопоставление данных онтогенетического, сравнительно-морфологического и палеоботанического исследований может восстановить реальный ход их исторического развития, что способствует выявлению родственных связей между таксонами и разработке эволюционной системы растений.
4. Изучение связи между структурой и функцией, между растением и условиями внешней среды.
Взаимодействие структуры и функции составляет основу жизнедеятельности любого организма. Функции без структуры не бывает, структура без функции бессмысленна. Ведь «изучать органы независимо от их отправлений, организмы независимо от их жизни почти так же невозможно, как изучать машину и ее части, не интересуясь их действием». Только соединение морфологического и физиологического методов исследования дает представление о растении как целостной структурно-функциональной и весьма динамичной системе, приспособленной к жизни в определенной экологической обстановке и чутко реагирующей на любые изменения внешних условий.
Реакции растений на неблагоприятные факторы среды их обитания проявляются сначала в биохимических и физиологических нарушениях, затем они затрагивают внутриклеточные структуры и, наконец, возникают изменения морфологического характера, заметные невооруженному глазу. Сначала они проявляются у отдельных растений, а впоследствии распространяются на все сообщество. Оценка уровня деградации растений под действием антропогенных факторов, прогнозирование возможных изменений растений под влиянием неблагоприятных условий составляют сущность ботанического мониторинга (от лат. и англ. monitor — предостерегающий). Его задача — вовремя сигнализировать обо всех случаях превышения отрицательных нагрузок, вызванных деятельностью человека, и принимать действенные меры для изменения режима эксплуатации растительных ресурсов и охраны растительного покрова как части глобальной проблемы сохранения генофонда и охраны окружающей среды.
Само собой очевидно, что морфология растений как фундаментальная ботаническая дисциплина абсолютно необходима для решения разнообразных практических задач: медицинских, лесохозяйственных, природоохранных и многих других. Перечислить все области применения морфологии растений вряд ли возможно.
Предлагаемый учебник посвящен морфологии высших растений. Прежде, чем перейти к анализу закономерностей их строения и демонстрации присущего им морфологического разнообразия, следует определить, что представляет собой растение как объект изучения, каковы его связи с другими живыми организмами, населяющими нашу планету, и, наконец, какое место в мире растений занимают высшие растения.
Источник: dist-tutor.info