Взаимодействие аллельных генов полное и неполное доминирование
Полное доминирование — взаимодействие двух аллелей одного гена, когда доминантный аллель полностью исключает проявление действия второго аллеля. В фенотипе присутствует только признак, задаваемый доминантной аллелью.
Неполное доминирование — доминантный аллель в гетерозиготном состоянии не полностью подавляет действие рецессивного аллеля. Гетерозиготы имеют промежуточный характер признака.
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной.
Кодоминирование — проявление у гибридов нового признака, обусловленного взаимодействием двух разных аллелей одного гена. Фенотип гетерозигот не является чем-то промежуточным между фенотипами разных гомозигот.
7.Множественный аллелизм. Генетика групп крови.
Множественный аллелизм — различные состояния (три и более) одного и того же локуса хромосом, возникшие в результате мутаций.
Присутствие в генофонде вида одновременно различных аллелей гена называют множественным аллелизмом.
имером этому служат разные варианты окраски глаз у плодовой мухи: белая, вишневая, красная, абрикосовая, эозиновая,— обусловленные различными аллелями соответствующего гена. У человека, как и у других представителей органического мира, множественный аллелизм свойствен многим генам. Так, три аллеля гена I определяют групповую принадлежность крови по системе АВ0 (IA, IB, I0). Два аллеля имеет ген, обусловливающий резус-принадлежность. Более ста аллелей насчитывают гены α- и β-полипептидов гемоглобина.
Причиной множественного аллелизма являются случайные изменения структуры гена (мутации), сохраняемые в процессе естественного отбора в генофонде популяции. Многообразие аллелей, рекомбинирующихся при половом размножении, определяет степень генотипического разнообразия среди представителей данного вида, что имеет большое эволюционное значение, повышая жизнеспособность популяций в меняющихся условиях их существования. Кроме эволюционного и экологического значения аллельное состояние генов оказывает большое влияние на функционирование генетического материала. В диплоидных соматических клетках эукариотических организмов большинство генов представлено двумя аллелями, которые совместно влияют на формирование признаков. .
У человека серией множественных аллелейпредставлен ген, определяющий группу крови. При этом гены, обусловливающие группы крови А и В, не являются доминантными по отношению друг к другу и оба доминантны по отношению к гену, определяющему группу крови О. Следует помнить, что в генотипе диплоидных организмов могут находиться только два гена из серии аллелей. Остальные аллели данного гена в разных сочетаниях входят в генотип других особей данного вида. Таким образом, множественный аллелизм характеризует разнообразие генофонда целого вида, т. е. является видовым, а не индивидуальным признаком.
Система групп крови АВО
Система групп крови ABO — это основная система групп крови, которая используется при переливании крови у людей. Ассоциированные анти-А и анти-В-антитела (иммуноглобулины), обычно относятся к типу IgM, которые, как правило, образуются в первые годы жизни в процессе сенситизации к веществам, которые находятся вокруг, в основном таких, как продукты питания, бактерии и вирусы. Система групп крови ABO также присутствует у некоторых животных, например, у обезьян (шимпанзе, бонобо и горилл).
.
Наследование групп крови системы АВО. В системе АВО синтез агглютиногенов и агглютининов определяется аллелями гена I:I0, IA, IB. Ген I контролирует и образование антигенов, и образование антител. При этом наблюдается полное доминирование аллелей IA и Ю над аллелем I0, но совместное доминирование (кодоминирование) аллелейIА и IB. Из-за кодоминирования наследование групп крови системы АВО происходит сложным образом. Например, если мать гетерозиготна по II группе, крови (генотип IAI0), а отец гетерозиготен по Ш группе крови (генотип IBI0), то в их потомстве с равной вероятностью может родиться ребенок с любой группой крови. Если у матери I группа крови (генотип I0I0), а у отца IV группа крови (генотип IАIB), то в их потомстве с равной вероятностью может родиться ребенок или со П (генотип IAI0), или с III (генотип IBI0) группой крови (но не с I, и не с IV).
Источник: StudFiles.net
Взаимодействие аллельных генов. Множественный аллелизм
Взаимодействие аллельных генов. Различают несколько типов взаимодействия аллельных генов.
Полное доминирование — взаимодействие, при котором доминантный аллель полностью подавляет проявление рецессивного аллеля. В этом случае как у гомозигот АД, так и у гетерозигот Аа проявляется доминантный признак. По такому типу наследуются многие признаки организмов. Например, у гороха желтый цвет семян полностью доминирует над зеленым, а гладкая поверхность семян — над морщинистой. У человека карий цвет глаз полностью доминирует над голубым, резус-поло-жительность — над резус-отрицательностью, праворукость — над леворукостью, шестипалость доминантна по отношению к нормальному строению кисти и т. д.
Некоторые аллельные гены взаимодействуют по типу неполного доминирования. В этом случае доминантный признак проявляется только при наличии двух доминантных аллелей в генотипе (АА). Если же организм гетерозиготен (Аа), т. е. имеет лишь один доминантный аллель, в фенотипе проявляется признак, промежуточный между доминантным и рецессивным.
Примером неполного доминирования является наследование окраски цветков у растений ночной красавицы. Как видно из рисунка 92, гомозиготные растения имеют либо красные (АА), либо белые (аа) цветки. При скрещивании растений с красными и белыми цветками в первом поколении у всех растений цветки будут розовыми (Ля), т. е. наблюдается промежуточный характер наследования.
При неполном доминировании изменяется характер расщепления во втором поколении, так как фенотип гетерозигот отличается от фенотипа гомозигот. В этом случае расщепление по фенотипу и по генотипу совпадает: 1 часть растений имеет белую окраску цветков (яя), 2 части — розовую (Ля) и 1 часть — красную (АА), или 1:2:1.

Неполное доминирование широко распространено. По такому типу наследуется курчавость волос у человека (курчавые волосы не полностью доминируют над прямыми, промежуточный признак — волнистые волосы), масть крупного рогатого скота, окраска оперения у кур и плодов земляники, другие признаки растений, животных и человека.
При кодоминировании у гетерозиготных особей полностью проявляются оба аллельных гена. Классическим примером ко-доминирования является взаимодействие генов у человека с четвертой группой крови (по системе АВО).
Группы крови О, А, В и АВ определяются геном /. Известны три разновидности этого гена: /А, Р, Аллельные гены /А и /вполностью доминируют над но кодоминантны по отношению друг к другу.
Ген /А обусловливает наличие на мембранах эритроцитов антигена А, ген Р определяет наличие другого антигена — В. Следовательно, у людей с генотипами /А/А и /А/’ эритроциты несут только антиген А — это вторая (А) группа крови. У людей с генотипами РР и РР красные кровяные клетки несут только антиген В — это кровь третьей (В) группы. Человек с генотипом /’/’ имеет первую (0) группу крови — на поверхности его эритроцитов отсутствуют антигены А и В.
У гетерозигот с генотипом 1АР гены /А и Р не подавляют проявление друг друга, а кодоминируют. Каждый из них в полной мере выполняет свою функцию, поэтому эритроциты содержат оба антигена (А и В). В результате формируется новый признак — четвертая (АВ) группа крови.
Множественный аллелизм. Окраска семян гороха, наследование которой изучал Мендель, определяется двумя аллелями — доминантным и рецессивным. Но многие гены представлены не двумя, а большим числом аллелей. Вы уже знаете, что ген, определяющий у человека группы крови (по системе АВО), существует в трех формах — /А, /в, Для гена, контролирующего синтез гемоглобина, известно несколько десятков разновидностей. При этом у каждой конкретной особи имеется не более двух аллелей каждого гена, поскольку соматические клетки содержат диплоидный набор хромосом.
Явление существования гена более чем в двух аллельных формах называется множественным аллелизмом. Множественные аллели нередко образуют серии, в которых каждый предыдущий аллель доминантен по отношению к последующим. Например, у кролика известна серия аллелей: С > cch > ch > са, где аллель С, определяющий черную окраску меха, доминирует над тремя другими: над аллелем, определяющим шиншилловую (cch), гималайскую (ch) и белую (са)окраску. Аллель шиншилловой окраски, в свою очередь, доминирует над аллелем гималайской окраски, а последний доминирует над аллелем, определяющим белую окраску (рис. 93). Известны серии множественных аллелей, определяющих цвет глаз у дрозофилы, рисунок на листьях белого клевера, и др.
Анализирующее скрещивание. При полном доминировании одного аллеля над другим организмы, имеющие доминантный фенотип, могут иметь разные генотипы — АА или Аа. Как определить генотип особи, обладающей доминантным признаком? Это можно сделать по результатам скрещивания исследуемой особи с рецессивной гомозиготой (аа). Такое скрещивание называется анализирующим.
Например, у томатов ген, контролирующий появление круглых плодов, полностью доминирует над геном, определяющим грушевидную форму. Нужно установить генотип растения с круглыми плодами. Введем обозначения соответствующих генов: А — круглые плоды, а. — грушевидные. Исследуемое растение может иметь генотип АА или Аа. В этом случае записывают фенотип и чес кий радикал — ту часть генотипа, которая определяет фенотип организма. В нашем случае фенотипический радикал следует записать в виде Л-.
Скрестим исследуемую особь с рецессивной гомозиготой, т. е. с растением, имеющим грушевидные плоды.
Р: А- х яя
круглые грушевидные
Рассмотрим два варианта анализирующего скрещивания (рис. 91), используя два возможных генотипа исследуемого растения (АА и Ля). Потомство, полученное при анализирующем скрещивании, принято обозначать как Fa. Обратите внимание, что в первом случае в потомстве наблюдается единообразие гибридов, а во втором — расщепление 1 = 1.
Итак, если потомство, полученное при анализирующем скрещивании, является единообразным по доминантному признаку, то анализируемая особь является гомозиготной (АА). Если же в потомстве наблюдается расщепление, исследуемый организм является гетерозиготным (Ля). Значит, анализирующее скрещивание — это скрещивание особи, имеющей доминантный фенотип, с гомозиготной рецессивной особью для выявления генотипа анализируемой особи.
Источник: www.qazbrand.info
Доминирование генов
Вы когда-нибудь задумывались, почему у вас есть такой особый цвет глаз или тип волос? Все это связано с передачей генов. Как обнаружил Грегор Мендель, черты наследуются при помощи передачи генов от родителей к их потомкам. Гены представляют собой участки ДНК, расположенные на наших хромосомах. Они передаются от одного поколения к другому через половое размножение. Ген для конкретного признака может существовать в более чем одной форме или аллеле. Для каждой характеристики или признака клетки животных обычно наследуют два аллеля. Парные аллели могут быть гомозиготными (с идентичными аллелями) или гетерозиготными (с разными аллелями) для данного признака.
Когда пары аллелей одинаковы, генотип этого признака идентичен, а фенотип или характеристика, которая наблюдается, определяется гомозиготными аллелями. Когда парные аллели для признака различные или гетерозиготные, возможно несколько вариантов. Гетерозиготные аллели, которые обычно наблюдаются в клетках животных, включают полное доминирование, неполное доминирование и кодоминирование.
Полное доминирование
» data-layzr=»https://natworld.info/wp-content/uploads/2017/07/горох.jpg» alt=»» width=»500″ height=»333″ data-layzr-srcset=»https://natworld.info/wp-content/uploads/2017/07/горох.jpg 500w, https://natworld.info/wp-content/uploads/2017/07/горох-300×200.jpg 300w» sizes=»(max-width: 500px) 100vw, 500px» />
При этом виде доминирования один аллель является доминирующим, а другой рецессивным. Доминантный аллель полностью маскирует рецессивный. Фенотип определяется доминирующим аллелем. Например, гены для формы семени в растениях гороха существуют в двух формах: гладкая (R) и морщинистая (r). В растениях гороха, которые являются гетерозиготными по форме семени, гладкий горох является доминирующим по отношению морщинистому семени, а генотип является (Rr).
Неполное доминирование
» data-layzr=»https://natworld.info/wp-content/uploads/2017/07/волнистые-волосы.jpg» alt=»» width=»500″ height=»332″ data-layzr-srcset=»https://natworld.info/wp-content/uploads/2017/07/волнистые-волосы.jpg 500w, https://natworld.info/wp-content/uploads/2017/07/волнистые-волосы-300×199.jpg 300w» sizes=»(max-width: 500px) 100vw, 500px» />
При неполном доминировании один аллель для конкретного признака не является полностью доминирующим над другим аллелем. Это приводит к третьему фенотипу, в котором наблюдаемые характеристики представляют собой смесь доминирующих и рецессивных фенотипов. Пример неполного доминирования проявляется в наследовании типа волос. Кудрявый тип волос (CC) является доминирующим для прямого типа волос (сс). Человек, гетерозиготный по этому признаку, будет иметь волнистые волосы (Cc).
Доминирующая фигурная характеристика не полностью выражена по прямой характеристике, создавая промежуточную характеристику волнистых волос. При неполном доминировании одна характеристика может быть несколько более заметной, чем другая для данного признака. Например, у человека с волнистыми волосами может быть больше или меньше волн, чем у другого с волнистыми волосами. Это указывает на то, что аллель для одного фенотипа выражается немного больше, чем аллель для другого фенотипа.
Кодоминирование
При совместном доминировании ни один из аллелей не является доминирующим, но оба аллеля для конкретного признака полностью выражены. Это приводит к третьему фенотипу, в котором наблюдается более одного фенотипа. Пример кодоминирования наблюдается у индивидов с чертой серповидной клетки.
Наличие серповидной клетки связано с развитием эритроцитов с аномальной формой. Нормальные эритроциты имеют двояковогнутую, дискообразную форму и содержат огромное количество белка, называемого гемоглобином. Гемоглобин помогает красным клеткам крови связываться и переносить кислород в клетки и ткани организма. Серповидная клетка является результатом мутации гена гемоглобина. Этот гемоглобин считается ненормальным и заставляет клетки крови принимать серповидную форму.
Серповидные клетки часто застревают в кровеносных сосудах и блокируют нормальный кровоток. Те, которые несут черту серповидной клетки, гетерозиготные для гена гемоглобина и наследуют один нормальный ген гемоглобина и один серповидный ген гемоглобина. У них нет болезни, потому что аллель гемоглобина серпа и нормальный гемоглобиновый аллель являются кодоминантными по отношению к форме клеток. Это означает, что у носителей серповидных клеток образуются как нормальные эритроциты, так и серповидные.
Неполное доминирование и кодоминирование
Люди часто путают неполное доминирование и кодоминирование. Хотя они и являются примерами наследования, но отличаются выражением генов. Ниже перечислены некоторые различия между ними:
Экспресивность аллеля
- Неполное доминирование: один аллель для определенного признака не полностью выражен над его парным аллелем. Например, цвет тюльпана, аллель для красного цвета (R) не полностью маскирует аллель для белого цвета (r).
- Кодоминирование: оба аллеля для конкретного признака полностью выражены. Аллель для красного цвета (R) и аллель для белого цвета (r) выражены и видны в гибриде.
Зависимость аллелей
- Неполное доминирование: влияние одного аллеля зависит от его парного аллеля для данного признака.
- Кодоминирование: влияние одного аллеля не зависит от его парного аллеля для данного признака.
Фенотип
- Неполное доминирование: Гибридный фенотип представляет собой смесь обеих аллелей, что приводит к третьему промежуточному фенотипу. Пример: Красный цветок (RR) X Белый цветок (rr) = Розовый цветок (Rr)
- Кодоминирование: гибридный фенотип представляет собой комбинацию выраженных аллелей, что приводит к третьему фенотипу, который включает оба фенотипа. Пример: Красный цветок (RR) X Белый цветок (rr) = Красно-белый цветок (Rr)
Наблюдаемые характеристики
- Неполное доминирование: фенотип может быть выражен в разной степени в гибриде. (Пример: розовый цветок может иметь более легкую или более темную окраску в зависимости от количественной выраженности одного аллеля по сравнению с другим).
- Кодоминирование: оба фенотипа полностью выражены в гибридном генотипе.
Краткий вывод
При неполном доминировании один аллель для конкретного признака не является полностью доминирующим над другим аллелем. Это приводит к третьему фенотипу, в котором наблюдаемые характеристики представляют собой смесь доминирующих и рецессивных фенотипов.
При кодоминировании ни один аллель не является доминирующим, но оба аллеля для конкретного признака полностью выражены. Это приводит к третьему фенотипу, в котором наблюдается более одного фенотипа.
Источник: natworld.info
Свойства генов
♦ Свойства генов и особенности их проявления в признаках:
■ ген дискретен в своем действии, т.е. обособлен в своей активности от других генов;
■ один ген отвечает за проявление одного строго определенного признака или нескольких признаков (плейотропия);
■ один признак может быть результатом действия нескольких генов (аллельных или неаллельных);
■ ген может усиливать степень проявления признака при увеличении числа его доминантных аллелей;
■ ген может взаимодействовать с другими генами; это приводит к появлению новых признаков;
■ изменение положения гена в хромосоме или влияние факторов внешней среды могут модифицировать его проявление в признаках;
■ ген обладает способностью к мутациям.
Взаимодействие аллельных генов
Взаимодействие генов — явление, когда за один признак отвечает несколько генов (или аллелей).
Аллельное взаимодействие — это взаимодействие аллелей одного и того же гена (за признак отвечает несколько аллелей одного и того же гена).
❖ Типы аллельных взаимодействий:
■ доминирование,
■ неполное доминирование,
■ сверхдоминирование,
■ кодоминирование.
Доминирование — тип взаимодействия двух аллелей одного гена, когда один (доминантный) из них полностью исключает действие другого (рецессивного). Примеры: доминирование у человека темных волос над светлыми, карих глаз над голубыми.
Неполное доминирование — степень активности доминантного аллеля недостаточна для того, чтобы полностью подавить действие рецессивного аллеля и обеспечить полное проявление доминантного признака.
■ В этом случае у гетерозигот формируется промежуточный (по отношению к родительским признакам) признак — имеет место промежуточный характер наследования. Этот признак будет наблюдаться у гибридов первого поколения и гетерозигот второго поколения. Во втором поколении расщепление по фенотипу и генотипу оказывается одинаковым 1:2:1 (одну часть составляет доминантная гомозигота АА с выраженным доминантным признаком, две части составляет гетерозигота Аа с промежуточным признаком и одну часть составляет гомозигота аа с рецессивным признаком).
■ Примеры неполного доминирования: наследование формы (курчавости) волос у человека, масти крупного рогатого скота, окраски цветков у растения ночная красавица (см. таблицу).
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа).
Кодоминирование — оба аллеля равноценны, не подавляют друг друга и участвуют в определении признака у гетерозиготной особи. Пример: наследование IV группы крови у человека, которая детерминируется одновременным присутствием в генотипе двух кодоминантных генов IА и Iв. Первый из этих генов детерминирует синтез в эритроцитах белка-антигена А, второй — синтез белка-антигена В; наличие обоих этих генов в генотипе приводит к тому, что у людей с IV группой крови эритроциты содержат как белок-антиген А, так и белок-антиген В.
Взаимодействие неаллельных генов
Неаллельное (или межаллельное) взаимодействие — это взаимодействие аллелей разных генов, т.е. генов, располагающихся в негомологичных хромосомах или разных локусах гомологичных хромосом.
■ Неаллельное взаимодействие генов приводит к модификации менделевского расщепления по фенотипу 9 : 3 : 1, т.е. к появлению в потомстве гетерозиготы иных расщеплений, например 9 : 3 : 4; 9 : 6 : 1; 12 : 3 : 1 и др.
❖ Основные типы межаллельных взаимодействий:
■ комплементарность;
■ эпистаз;
■ полимерия.
Замечание: комплементарное и эпистатическое взаимодействия возникают в тех случаях, когда признак контролируется одной парой неаллельных генов.
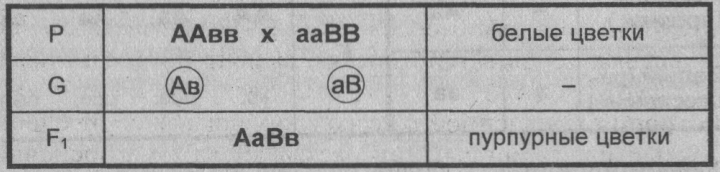
Комплементарное, или дополнительное, взаимодействие — такой тип межаллельного взаимодействия генов, при котором одновременное присутствие в генотипе гибрида доминантных генов разных аллельных пар приводит к появлению нового признака, отсутствующего у обоих родителей.
Пример: наследование окраски цветков душистого горошка (родительские растения с генотипами А-вв, ааВ- имеют белые цветки, гибриды с генотипом А-В- пурпурные; см. таблицу).
Замечание: знак «-» в формуле генотипа означает, что это место может занимать как доминантный, так и рецессивный аллель.
Объяснение: пурпурный пигмент образуется с помощью специального фермента, который синтезируется только при наличии обоих доминантных генов: как А, так и В. Цветки родительских особей имеют белый цвет, так как в генотипе каждого из них присутствует только один из этих генов.
Во втором поколении при самоопылении, обеспечивающем равновероятное (случайное) образование гамет и зигот разного типа, наблюдается расщепление по фенотипу в соотношении пурпурной и белой окраски цветков как 9 : 7 (9 пурпурных: A-В- и 7 белых: ЗА-Bb, ЗааВ-. 1aabb).
Эпистаз — такой тип межаллельного взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена, и подавляемый признак не проявляется.
Супрессор (или ген-ингибитор) — ген, подавляющий действие других неаллельных генов. Супрессором может быть как доминантный, так и рецессивный ген.
Доминантный эпистаз — эпистаз, в котором супрессором является доминантный ген. При доминантном эпистазе во втором поколении наблюдается расщепление фенотипа 12:3:1 или 13:3.
Рецессивный эпистаз (криптомерия) — эпистаз, в котором супрессором является рецессивный ген. При рецессивном эпистазе во втором поколении наблюдается расщепление по фенотипу 9:3:4.
Пример эпистаза: наследование окраски шерсти у домашних кроликов. Синтез черного пигмента детерминирует рецессивный ген с, доминантная аллель I другого гена является супрессором, подавляя действие гена с. Тогда кролики с генотипами C-I-, ccl-будут белыми, кролики с генотипами C-ii — серыми, а с генотипом ссii — черными.
Многие признаки контролируются двумя и более парами неаллельных генов (называемых в этом случае полимерными).
Полимерия — взаимодействие нескольких неаллельных полимерных генов. При полимерии степень выраженности фенотипического признака часто зависит от числа полимерных генов, ответственных за его проявление. При кумулятивной полимерии действие генов суммируется; примеры: масса тела, молочность крупного рогатого скота, яйценоскость кур, некоторые параметры умственных способностей человека и др. При некумулятивной полимерии степень проявления признака от числа доминантных генов в генотипе не зависит (пример: оперенность ног у кур).
Плейотропия — зависимость нескольких признаков от одного гена. Каждый плейотропный ген оказывает какое-то основное действие, но модифицирует проявление других генов.
Сцепление генов. Опыты Моргана
Сцепленные гены — любые гены, расположенные в одной хромосоме.
Группа сцепления — все гены, расположенные в одной хромосоме.
■ Количество групп сцепления равно числу пар хромосом (т.е. гаплоидному числу хромосом). У человека 46 хромосом, т.е. 23 группы сцепления.
■ Наследование признаков, за которые отвечают гены из одной группы сцепления, не подчиняется законам Менделя.
Опыты Т. Моргана (1911-1912 гг.): анализ наследования двух пар альтернативных признаков у мух-дрозофил — серого (В) и черного (b) цвета тела и нормальной (V) или укороченной (v) длины крыльев.
Первая серия опытов: скрещивание гомозиготной доминантной (BBVV) особи (с серым цветом тела и нормальной длиной крыльев) с гомозиготной рецессивной (bbvv) особью черного цвета с короткими крыльями. Все потомки F1, в соответствии с первым законом Менделя, являются доминантными гетерозиготными (BbVv) особями серого цвета с нормальными крыльями.
Вторая серия опытов: анализирующее скрещивание гибридов первого поколения — гомозиготной рецессивной (черной короткокрылой) самки (bbvv) с дигетерозиготным (серым с нормальными крыльями) самцом (BbVv). Если предположить, что два гена, относящиеся к разным аллельным парам, локализованы в разных хромосомах, то у дигетерозиготы следует ожидать образования (в равных количествах) четырех типов гамет: BV, bV, Bv и bv. Тогда, согласно третьему закону Менделя, в потомстве должны присутствовать четыре разных фенотипа в равном количестве (по 25%). В действительности присутствовали только два фенотипа (в соотношении 1 : 1).
■ Это означает, что доминантные гены В и V, относящиеся к разным аллельным парам, локализованы в одной хромосоме (из пары гомологичных хромосом) и попадают в одну гамету, а оба рецессивных гена в и v локализованы в другой хромосоме и вместе попадают в другую гамету. Поэтому у дигетерозиготного самца мухи-дрозофилы образуется не четыре типа гамет (когда гены расположены в разных хромосомах), а только два: BV (50%) и bv (50%), и, следовательно, потомки F2 будут иметь два сочетания признаков.
Третья серия опытов: проверка предположения о полном сцеплении генов путем анализирующего скрещивания дигетерозиготной (серой с нормальными крыльями) самки (BbVv) из поколения F1 с гомозиготным рецессивным (черным короткокрылым) самцом (bbvv) из родительского поколения. В результате были получены потомки четырех фенотипов в следующем соотношении: по 41,5% особей с серым телом и нормальными крыльями (генотип BbVv) и особей с черным телом и короткими крыльями (генотип bbvv), и по 8,5% серых короткокрылых особей (генотип Bbvv) и черных особей с нормальными крыльями (генотип bbVv).
Отсюда следует, что сцепленные гены, т.е. гены, локализованные в одной хромосоме, не всегда передаются вместе, т.е. сцепление может быть неполным. Это связано с явлением кроссинговера. вероятность которого в данном случае составляет 17%.
Кроссинговер
Кроссинговер — явление обмена участками гомологичных хрома-тид в процессе их конъюгации в профазе мейоза I.
■ У гетерозиготных организмов кроссинговер приводит к пере-комбинации генетического материала.
■ Кроссинговер происходит не всегда; его частота зависит от расстояния между генами (подробнее о расстоянии между генами см. ниже).
■ Кроссинговер имеет место у всех растений и животных, за исключением самца мухи-дрозофилы и самки тутового шелкопряда.
■ Значение кроссинговера: он позволяет создавать новые комбинации генов и тем самым повысить наследственную изменчивость, необходимую для расширения возможностей адаптации организмов к изменившимся условиям среды.
Рекомбинация — появление новых сочетаний генов в результате кроссинговера, свободного комбинирования хромосом при образовании гамет или при их слиянии в ходе оплодотворения.
Кроссоверные (или рекомбинантные) особи — особи, образующиеся из гамет с новым, полученным путем кроссинговера, сочетанием аллелей.
Закон Моргана (закон сцепленного наследования): гены, расположенные в одной хромосоме (т.е. входящие в одну группу сцепления), наследуются преимущественно, т.е. с наибольшей вероятностью, вместе (сцепленно).
Хромосомная теория наследственности
Хромосомная теории наследственности экспериментально обоснована Т. Морганом с сотрудниками в 1911-1926 гг.
♦ Основные положения хромосомной теории наследственности:
■ гены, ответственные за хранение и передачу наследственной информации, локализованы в определенных участках (локусах) хромосом; различные хромосомы имеют разное число генов;
■ аллельные гены занимают одинаковые локусы в гомологичных хромосомах;
■ в хромосомах гены располагаются в линейной последовательности (друг за другом) и не перекрываются;
■ гены гомологичных хромосом образуют группу сцепления и наследуются преимущественно вместе; число групп сцепления равно гаплоидному набору хромосом (т.е. числу пар гомологичных хромосом);
■ между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер);
■ вероятность кроссинговера пропорциональна расстоянию между генами в хромосомах.
Генетические карты
Генетическая карта хромосом — схема, на которой отображается взаимное расположение генов, находящихся в одной группе сцепления, с учетом их порядка следования и относительных расстояний между ними.
■ Возможность картирования хромосом основана на постоянстве процента кроссинговера между определенными генами.
Расстояние между генами выражается в морганидах.
■ Одна морганида — такое расстояние между генами, при котором вероятность кроссинговера равна 1 %.
♦ Значение генетических карт:
■ в селекции они позволяют прогнозировать возможность получения организмов с определенными сочетаниями признаков;
■ являются основой для генной инженерии;
■ в медицине используются для диагностики ряда тяжелых наследственных заболеваний человека;
■ сравнение генетических карт разных видов живых организмов помогает установить особенности эволюционного процесса.
Цитоплазматическая наследственность
Цитоплазматическая наследственность — наследственность, связанная с действием генов, находящихся в органоидах цитоплазмы, содержащих ДНК (митохондриях и пластидах).
■ Такие гены способны к автономной репликации и равномерному распределению между дочерними клетками.
■ В передаче признаков цитоплазматическая наследственность имеет второстепенное значение.
■ Цитоплазматическое наследование осуществляется только через материнский организм (в мужских половых клетках цитоплазмы мало и в ней митохондрии и пластиды отсутствуют). Примеры: с мутациями генов, локализованных в митохондриях, связано наследование нарушений в действии дыхательных ферментов у дрожжей; с мутациями генов, локализованных в пластидах, связано, в частности, наследование пестролистности у ряда растений (ночной красавицы, львиного зева и др.).
Метки: генетика
Источник: esculappro.ru
Взаимодействие аллельных генов
Различают три типа взаимодействия аллельных генов: полное доминирование, неполное доминирование, кодоминирование.
- Полное доминирование — явление, когда доминантный ген полностью подавляет работу рецессивного гена, в результате чего развивается доминантный признак.
- Неполное доминирование — явление, когда доминантный ген не полностью подавляет работу рецессивного гена, в результате чего развивается промежуточный признак.
- Кодоминирование (независимое проявление) — явление, когда в формировании признака у гетерозиготного организма участвуют оба аллеля. У человека с серией множественных аллелей представлен ген, определяющий группу крови. При этом гены, обусловливающие группы крови A и B, являются кодоминантными по отношению друг к другу и оба доминантны по отношению к гену, определяющему группу крови 0.
Взаимодействие неаллельных генов
Различают четыре типа взаимодействия неалльлельных генов: кооперацию, комплементарность, эпистаз и полимерию.
Кооперация — явление, когда при взаимном действии двух доминантных неаллельных генов, каждый из которых имеет свое собственное фенотипическое проявление, происходит формирование нового признака.
Комплементарность — явление, когда признак развивается только при взаимном действии двух доминантных неаллельных генов, каждый из которых в отдельности не вызывает развитие признака.
Эпистаз — явление, когда один ген (как доминантный, так и рецессивный) подавляет действие другого (неаллельного) гена (как доминантного, так и рецессивного). Ген-подавитель (супрессор) может быть доминантным (доминантный эпистаз) или рецессивным (рецессивный эпистаз).
Полимерия — явление, когда несколько неаллельных доминантных генов отвечают за сходное воздействие на развитие одного и того же признака. Чем больше таких генов присутствуют в генотипе, тем ярче проявляется признак. Явление полимерии наблюдается при наследовании количественных признаков (цвет кожи, масса тела, удой коров).
В противоположность полимерии наблюдается такое явление, как плейотропия — множественное действие гена, когда один ген отвечает за развитие нескольких признаков.
Источник: jbio.ru