Y хроматин это
Хроматин — это вещество хромосом , представляющее собой комплекс ДНК , РНК и белков . Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот . Именно в составе хроматина происходит реализация генетической информации , а также репликация и репарация ДНК .
До 25—40% сухого веса хроматина составляют гистоновые белки. Гистоны являются компонентом нуклеосом , надмолекулярных структур, участвующих в упаковке хромосом. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из гистонов четырёх типов: H2A, H2B, H3 и H4. Эти гистоны называются кóровыми. В одну нуклеосому входят по два кóровых гистона каждого типа — всего восемь белков. Линкерный гистон H1, более крупный, чем кóровые гистоны, связывается с ДНК в месте её входа на нуклеосому.
Нить ДНК с нуклеосомами образует нерегулярную соленоид -подобную структуру толщиной около 30 нанометров , так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином , он хорошо видим под микроскопом. ДНК, находящаяся в гетерохроматине не транскрибируется , обычно это состояние характерно для незначащих или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра . Полная конденсация хромосом происходит перед делением клетки.
Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется транскрипционной активностью. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием , фосфорилированием , метилированием и другими модификациями.
Считается, что в ядре существуют так называемые функциональные домены хроматина (ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». Вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные и центромерные (отвечающие за связывание сестринских хроматид в митозе ) участки хромосом закреплены на белках ядерной ламины .
Источник: xn--b1algemdcsb.xn--p1ai
ПОЛОВОЙ ХРОМАТИН — участок ядра соматической клетки, находящейся в интерфазе, представляющий собой конденсированную половую хромосому; в результате конденсации Х-хромосомы образуется X-хроматин, а конденсации Y-хромосомы — Y-хроматин. У людей с нормальным составом хромосом (см. Кариотип) в соматических клетках женщин содержится X-хроматин, а в соматических клетках мужчин — Y-хроматин. По наличию этих образований может быть определен генетический пол индивида (см. Пол), что находит практическое применение при диагностике разнообразных клин, форм тестикулярной и овариальной дисгенезии (см. Дисгенезия гонад), ориентировочном исследовании индивидов на ложный — псевдогермафродитизм (см.) или истинный гермафродитизм (см.), в суд.-мед. практике и т. п.
X-хроматин (так наз. тельце Барра) как специфический для женского пола цитол, признак впервые был описан Барром (М. Barr) и Бертрамом (E. G. Bertram) в 1949 г.; одна из двух X-хромосом в раннем эмбриогенезе генетически инактивируется (гетерохроматинизируется) и остается конденсированной в течение всего интерфазного периода жизни соматической клетки. Деконденсация гетерохроматинизированной X-хромосомы в соматической клетке не означает ее генетической активации. Вторая Х-хромосома в женской клетке и Х-хромосома в мужской соматической клетке не образуют X-хроматина. Y-хроматин формируется за счет части длинного плеча Y-хромосомы; он состоит из гетерохроматина, конденсируемого в интерфазном ядре и способного интенсивно флюоресцировать после окраски акрихином и другими флюорохромами (см.). Тест с флюорохромами для определения половой принадлежности клеток по Y-хроматину был впервые предложен Касперссоном (Т. Caspersson) в 1970 г.
При определении генетического пола индивида его клетки исследуют как на Х-, так и на Y-хроматин. Для этого получают клеточный материал (источником его могут быть самые разнообразные ткани, хотя предпочтительнее те, для к-рых не требуется предварительного культивирования in vitro; для определения X-хроматина чаще всего используют мазки со слизистой оболочки щеки, реже слизистой оболочки влагалища, а также клетки волосяных фолликулов; для пренатальной диагностики пола плода используют амниотические клетки. Содержание Y-хрома-тина, помимо перечисленных тканей, можно определять также в сперматозоидах. Прекрасным материалом для анализа X- и Y-хроматина являются однослойные культуры клеток, обычно фибробластов. В культивируемых лимфоцитах крови хорошо выявляется Y-хроматин. Фиксацию препаратов проводят до их подсушивания на воздухе, обычно фиксируют метанолом или смесью этанола и уксусной к-ты (в соотношении 3:1) либо только этанолом. Методы окраски для выявления X- и Y-хроматина различны. X-хроматин лучше выявляется при окраске препаратов основными нефлюоресцирующими красителями: основным фуксином, тионином, ацетоорсеином, толуидиновым синим и др. Окрашенные препараты высушивают и изучают с масляной иммерсией в проходящем свете. Y-хроматин выявляют при окраске препаратов флюорохромами — производными акридинового оранжевого: акрихином, акрихин-ипритом, акрихин-пропилом. Препараты заключают в специальный буферный р-р и изучают в ультрафиолетовом свете с помощью люминесцентного микроскопа (см.). Анализ проводят на разъединенных, распластанных клетках. Срезы тканей для определения П. х. используют лишь тогда, когда невозможно получить мазки или препараты-отпечатки среза органа.
Тельце Барра в нормальной диплоидной клетке имеет форму треугольника, круглого или даже палочковидного образования, средний линейный размер к-рого 1 мкм. Очень часто тельце Барра находится на периферии ядра и нередко соприкасается с ядерной оболочкой. Размер, форма, положение в ядре и плотная окраска позволяют отличать X-хроматин от глыбок конденсированного хроматина других хромосом. Частота, с к-рой X-хроматин встречается в клетках, зависит от состояния организма (гормональный статус, физическая нагрузка и др.)* Она низка у новорожденных девочек в первые 2—3 дня жизни. У половозрелой женщины частота встречаемости X-хроматина колеблется в разные периоды менструального цикла, различна в разных тканях и никогда не достигает 100% . В клетках слизистой оболочки щеки X-хроматин обнаруживается в 25—60% клеток, в культивируемых in vitro фибробластах — в 40—80% клеток, в некультивируемых амниотических клетках примерно в 5% клеток. В гранулоцитах X-хроматин имеет вид барабанной палочки; низкая частота выявления (1,5—5%) и трудность дифференцировки этого образования с другими ядерными структурами ограничили практическое распространение этого теста. В полиплоидных клетках число телец Барра кратно числу диплоидных наборов хромосом. У мужчин цитологически сходные с X-хроматином глыбки хроматина обнаруживаются примерно в 1% клеток.
Размеры Y-хроматина в нормальных диплоидных клетках у разных индивидов сильно различаются, что связано с большими колебаниями длины самой Y-хромосомы. При диагностике пола по Y-хроматину следует помнить, что встречаются индивиды или с незначительным количеством гетерохроматина в Y-xpo-мосоме или вовсе лишенные гетерохроматина, и необнаружение Y-хроматина у таких лиц не является фактом, отрицающим их принадлежность к мужскому полу. У большинства мужчин Y-хроматин выглядит сравнительно крупным (0,3—1 мкм), ярко светящимся образованием (тельцем) обычно округлой формы; в отдельных клетках он может иметь двойную структуру или быть несколько диффузным. На периферии клеточного ядра Y-тельце располагается реже, чем тельце Барра. В ядре присутствуют ярко светящиеся глыбки хроматина других хромосом, отличить которые от Y-тель-ца обычно нетрудно. Частота встречаемости Y-хроматина существенно различается в разных тканях. В клетках слизистой оболочки щеки он обнаруживается в 20—80% клеток, в клетках волосяного фолликула в 70—90%, в лимфоцитах периферической крови в 60—87% клеток. Сходные с Y-тельцем флюоресцирующие тельца в клетках женщины (аутосомный гетерохроматин) встречаются примерно в 5% клеток. У женщин, беременных плодом мужского пола, клетки последнего могут проникать в кровяное русло матери. Они обнаруживаются по наличию в них Y-хроматина. Для определения генетического пола по Y-хроматину рекомендуется просмотреть не менее 50 клеток.
Практическое применение теста на П. х. двояко. Этот тест используется для определения пола индивида по его клеткам, когда либо сам индивид недоступен для исследования (пренатальная диагностика пола плода, суд.-мед. экспертиза и т. п.), либо в тех случаях, когда проводится массовая проверка соответствия фенотипического (паспортного) пола генетическому (напр., при обследовании женщин на спортивных соревнованиях). Определение пола плода внутриутробно проводят при подозрении на наследственную болезнь, сцепленную с полом (гемофилия, некоторые формы мышечной дистрофии и др.), с целью предотвращения рождения неизлечимо больного ребенка.
При обследовании пациентов с клин. проявлениями нарушения половой дифференцировки и в других случаях, когда необходимо точное описание состояния половых хромосом, независимо от результатов анализа на П. х. прибегают к хромосомному анализу (см. Хромосомы).
Особенно часто определение П. х. используется для предварительной диагностики отклонений в числе или структуре половых хромосом, когда у исследуемого имеются нарушения полового развития. Простота и быстрота выполнения позволяют применять тест на П. х. при массовом обследовании новорожденных и других детей с целью выявления аномалий половых хромосом.
ончательный диагноз ставят после изучения хромосомного набора (см.). Отклонения в системе половых хромосом в сторону уменьшения,количества их материала, совместимые с жизнеспособностью индивида, происходят за счет гетерохроматинизированной Х-хромосомы и гетерохроматиновой части Y-хромосомы и поэтому могут быть установлены путем анализа полового хроматина. Отсутствие X-хроматина в клетках пациента с женским фенотипом наблюдается при полной агенезии гонад при синдроме Шерешевского — Тернера с формулой кариотипа 45,X (см.Тернера синдром) либо у лиц с кариотипом 46,XY при тестикулярной феминизации (см.) и при хрома-тин-отрицательной форме дисгенезии гонад. При дисгенезии яичников хромосомного генеза частота встречаемости и размеры тельца Барра зависят от характера отклонений в Х-хромосоме и коррелируют с клин. полиморфизмом болезни. Тельце Барра может иметь нормальную величину, но обнаруживаться редко или даже вовсе отсутствовать в отдельных тканях (мозаичная форма синдрома Шерешевского — Тернера, формула кариотипа обычно 46,XХ/45,X); оно может быть уменьшенным по сравнению с нормой (делеция короткого или длинного плеча Х-хромосомы, кольцевая Х-хромосома) или, наоборот, увеличенным (по длинному плечу Х-изохромосомы). Разнообразные клин, формы тестикулярной дисгенезии коррелируют с характером отклонений в Y-хромосоме и имеют разную картину Y-хроматина: от его отсутствия или низкого содержания (мозаицизм с формулой кариотипа 46,XY/45,X) до изменений в величине и форме (делеции длинного плеча изохромосомы, дицентрические хромосомы). Исследования П. х. при разных формах истинного и ложного гермафродитизма проводятся только как ориентировочные, за к-рыми должно следовать тщательное исследование хромосомных наборов.
При нарушениях полового развития, связанных с увеличением числа X-хромосом, в клетках пациенток обнаруживаются дополнительные тельца Барра. Их число определяется формулой n — 1, где n — общее количество X-хромосом у индивида. Так, при кариотипе 47,XXX в клетках женщины обнаруживаются два тельца Барра, при кариотипе 48,ХХХХ — три тельца Барра. Дополнительные Y-хромосомы проявляют себя появлением в клетке дополнительных телец Y (при кариотипе 47,XYY — два Y-тельца, при кариотипе 48,XYYY — три Y-тельца). При синдроме Клайнфелтера (см. Клайнфелтера синдром), когда Кариотип пациента включает две или более Х-хромосомы и одну или более Y-хромосому, в соматических клетках одновременно содержатся X- и Y-хроматин (по одному тельцу каждого при наиболее часто встречающейся форме синдрома с формулой кариотипа 47,XXY).
Источник: xn--90aw5c.xn--c1avg

Хроматин — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК.
При наблюдении некоторых живых клеток, особенно растительных или же клеток после фиксации и окраски, внутри ядра выявляются зоны плотного вещества. В состав хроматина входит ДНК в комплексе с белком. В интерфазных клетках хроматин может равномерно заполнять объем ядра или же располагаться отдельными сгустками (хромоцентры). Часто он особенно четко выявляется на периферии ядра (пристеночный, примембранный хроматин) или образует внутри ядра переплетения довольно толстых (около 0. 3 мкм) и длинных тяжей, образующих подобие внутриядерной цепи.
Хроматин интерфазных ядер представляет собой несущие ДНК тельца (хромосомы), которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной в ядрах разных клеток. Когда хромосома или ее участок полностью деконденсирован, тогда эти зоны называют диффузным хроматином. При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина (иногда называемого гетерохроматин). Показано, что степень деконденсации хромосомного материала в интерфазе может отражать функциональную нагрузку этой структуры. Чем более диффузен хроматин интерфазного ядра, тем выше в нем синтетические процессы. Падение синтеза РНК в клетках обычно сопровождается увеличением зон конденсированного хроматина.
Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных телец — хромосом. В этот период хромосомы не несут никаких синтетических нагрузок, в них не происходит включение предшественников ДНК и РНК.
Исходя из этого можно считать, что хромосомы клеток могут находиться в двух структурно-функциональных состояниях:
В рабочем, частично или полностью деконденсированном, когда с их участием в интерфазном ядре происходят процессы транскрипции и редупликации;
В неактивном — в состоянии метаболического покоя при максимальной их конденсированности, когда они выполняют функцию распределения и перенося генетического материала в дочерние клетки.
В химическом отношении препараты хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов, в состав которых входит ДНК и специальные хромосомные белки — гистоны. В составе хроматина обнаружено также РНК. В количественном отношении ДНК, белок и РНК находятся как 1: 1, 3: 0, 2. О значении РНК в составе хроматина еще нет достаточно однозначных данных. Возможно, что эта РНК представляет собой сопутствующую препарату функцию синтезирующейся РНК и поэтому частично связанной с ДНК или это особый вид РНК, характерный для структуры хроматина.
Схема конденсации хроматина:
Источник: biology623.blogspot.com
Хроматин (греч.chroma — цвет, краска и греч.nitos — нить) — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК.
Эухроматин, активный хроматин — участки хроматина, сохраняющие деспирализованное состояние элементарных дезоксирибонуклеопротеидных нитей (ДНП) в покоящемся ядре, т. е. в интерфазе (в отличие от других участков, сохраняющих спирализованное состояние — гетерохроматина).
Эухроматин отличается от гетерохроматина также способностью к интенсивному синтезу рибонуклеиновой кислоты (РНК) и большим содержанием негистоновых белков. В нём, помимо ДНП, имеются рибонуклеопротеидные частицы (РНП-гранулы) диаметром 200—500, которые служат для завершения созревания РНК и переноса ее в цитоплазму. Эухроматин содержит большинство структурных генов организма.
Гетерохроматин — участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость.
Особенности структуры и состава гетерохроматина
Хроматин является нуклеопротеидом — комплексом ДНК с гистонами. Конденсация хроматина в гетерохроматин сопровождается как модификацией гистонов, так и усложнением состава нуклеопротеидного комплекса за счёт участия в нём белков гетерохроматина HP1 (HeterochromatinProtein 1).
Гистоны гетерохроматинового комплекса характеризуются низкой степенью ацетилированности по лизиновым остаткам, что увеличивает их основные свойства и, соответственно, связывание с кислымифосфатными группами ДНК, что способствует компактификации комплекса. Другой особенностью, ведущей к образованию гетерохроматина, является метилирование 9-го лизинового остатка гистона H3 метилтрансферазой Suv39h; такое метилирование ведёт к образованию высокоаффинного сайта связывания гистона H3 и белка гетерохроматина HP1. У дрозофилметилтрансфераза Suv39h функционально ассоциирована с гистондеацетилазой таким образом, что ацетилированное и метилированное состояние 9-го лизинового остатка гистона H3 являются взаимоисключающими, то есть обеспечивается единый механизм деацетилирования и метилирования гистона H3, ведущий к усилению связывания с гистоном как ДНК, так и белка гетерохроматина HP1.
22. Морфо – функциональная характеристика хромосом. Денверская и Парижская классификации хромосом.
Типы строения хромосом
Различают четыре типа строения хромосом:
телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце);
акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
субметацентрические (с плечами неравной длины, напоминающие по форме букву L);
метацентрические (V-образные хромосомы, обладающие плечами равной длины).
Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода.
Спутники (сателлиты)
Сателлит — это округлое или удлинённое тельце, отделённое от основной части хромосомы тонкой хроматиновой нитью, по диаметру равный или несколько меньший хромосоме. Хромосомы, обладающие спутником принято обозначать SAT-хромосомами. Форма, величина спутника и связывающей его нити постоянны для каждой хромосомы.
Зона ядрышка
Зоны ядрышка (организаторы ядрышка) — специальные участки, с которыми связано появление некоторых вторичных перетяжек.
Хромонема
Хромонема — это спиральная структура, которую удаётся увидеть в декомпактизованных хромосомах через электронный микроскоп. Впервые наблюдалась Баранецким в 1880 году в хромосомах клеток пыльниковтрадесканции, термин ввёл Вейдовский. Хромонема может состоять из двух, четырёх и более нитей, в зависимости от исследуемого объекта. Эти нити образуют спирали двух типов:
паранемическую (элементы спирали легко разъединить);
плектонемическую (нити плотно переплетаются).
Первичная перетяжка
Хромосомная перетяжка (X. п.), в которой локализуется центромера и которая делит хромосому на плечи.
Вторичные перетяжки
Морфологический признак, позволяющий идентифицировать отдельные хромосомы в наборе. От первичной перетяжки отличаются отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. У человека это 9, 13, 14, 15, 21 и 22 хромосомы.
В 1960 году в Денвере (США) была разработана и принята Денверская классификация хромосом человека .В основе этой классификации лежит различение хромосом по :
1 Размеру
2 По соотношению плеч
Денверевская классификация позволяет четко различать хромосомы ,принадлежащие к различным группам .
Парижская классификация хромасом основывается на :дифференциальных методах окрашивания метафазных хромасом . На Парижской конференции по номенклатуре в цитогенетике человека была разработана и в настоящее время вошла в практику цитогенетического анализа система обозначения сегментов нормальных хромосом и хромосом, подвергшихся тем или иным структурным перестройкам (1971). Парижская классификация позволяет установить : является ли данный признак наследственным (по проявлению его у родственников); тип и характер наследования (доминантный или рецессивный, аутосомный или гоносомный);зиготность лиц родословной (гомо- или гетерозиготы);пенетрантность гена (частота его проявления); вероятность рождения ребенка с наследственной патологией (генетический риск).
Источник: studopedia.su
Определение понятия
Хроматином называется содержащий наследственную информацию материал клеточного ядра, представляющий собой сложный функциональный комплекс ДНК со структурными белками и другими элементами, обеспечивающими упаковку, хранение и реализацию кариотического генома. В упрощенной трактовке это вещество, из которого состоят хромосомы. Термин происходит от греческого «хрома» – цвет, краска.
Понятие было введено Флемингом еще в 1880 году, но до сих пор идут споры о том, что такое хроматин, с точки зрения биохимического состава. Неопределенность касается небольшой части компонентов, не участвующих в структурировании генетических молекул (некоторые ферменты и рибонуклеиновые кислоты).
На электронной фотографии интерфазного ядра хроматин визуализируется как многочисленные участки темной материи, которые могут быть мелкими и разрозненными или объединяться в крупные плотные скопления.
Конденсация хроматина во время клеточного деления приводит к образованию хромосом, которые видны даже в обычном световом микроскопе.
Структурные и функциональные компоненты хроматина
С целью определить, что такое хроматин на биохимическом уровне, ученые экстрагировали это вещество из клеток, переводили в раствор и в таком виде изучали компонентный состав и структуру. При этом использовались как химические, так и физические методы, включая технологии электронной микроскопии. Выяснилось, что химический состав хроматина на 40% представлен длинными молекулами ДНК и почти на 60% – различными белками. Последние подразделяются на две группы: гистоны и негистоновые.
Гистонами называют большое семейство основных ядерных белков, которые прочно связываются с ДНК, формируя структурный скелет хроматина. Их количество примерно равно процентному содержанию генетических молекул.
Остальная часть (до 20%) протеиновой фракции приходится на ДНК-связывающие и пространственно-модифицирующие белки, а также ферменты, принимающие участие в процессах считывания и копирования генетической информации.
Помимо основных элементов, в составе хроматина в небольшом количестве обнаруживаются рибонуклеиновые кислоты (РНК), гликопротеиды, углеводы и липиды, однако вопрос об их ассоциации с ДНК-упаковочным комплексом до сих пор открыт.
Гистоны и нуклеосомы
Молекулярная масса гистонов варьирует в пределах от 11 до 21 кДа. Большое количество остатков основных аминокислот лизина и аргинина придают этим белкам положительный заряд, способствуя формированию ионных связей с противоположно заряженными фосфатными группами двойной спирали ДНК.
Выделяют 5 разновидностей гистонов: H2A, H2B, H3, H4 и H1. Первые четыре типа участвуют в формировании основной структурной единицы хроматина – нуклеосомы, которая состоит из кора (белковой сердцевины) и обмотанной вокруг него ДНК.
Нуклеосомный кор представлен октамерным комплексом из восьми молекул гистонов, в который входят тетрамер H3-H4 и димер Н2A-H2B. Участок ДНК протяженностью около 146 нуклеотидных пар накручивается на поверхность белковой частицы, образуя 1,75 витка, и переходит в линкерную последовательность (примерно 60 н. п.), соединяющую нуклеосомы друг с другом. Молекула H1 связывается с линкерной ДНК, защищая ее от действия нуклеаз.
Гистоны могут подвергаться различным модификациям, таким как ацетилирование, метилирование, фосфорилирование, ADP-рибозилирование и взаимодействие с убивиктиновым белком. Эти процессы влияют на пространственную конфигурацию и плотность упаковки ДНК.
Негистоновые белки
Существует несколько сотен разновидностей негистоновых белков с различными свойствами и функциями. Их молекулярная масса варьирует от 5 до 200 кДа. Особую группу составляют сайт-специфические белки, каждый из которых комплементарен определенному участку ДНК. В эту группу входят 2 семейства:
- «цинковые пальцы» – узнают фрагменты длиной в 5 нуклеотидных пар;
- гомодимеры – характеризуются структурой «спираль-поворот-спираль» во фрагменте, связанном с ДНК.
Лучше всего изучены так называемые белки высокой подвижности (HGM-белки), постоянно ассоциированые с хроматином. Такое наименование семейство получило из-за высокой скорости перемещения белковых молекул в электрофорезном геле. Эта группа занимает большую часть негистоновой фракции и включает в себя четыре основных типа HGM-белков: HGM-1, HGM-14, HGM-17 и HMO-2. Они выполняют структурную и регуляторную функции.
К негистоновым белкам относят также ферменты, обеспечивающие транскрипцию (процесс синтеза матричной РНК), репликацию (удвоение ДНК) и репарацию (устранение повреждений в генетической молекуле).
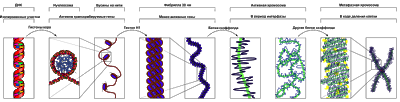
Уровни компактизации ДНК
Особенность структуры хроматина такая, что позволяет нитям ДНК с суммарной длиной более метра поместиться в ядро диаметром около 10 мкм. Такое возможно благодаря многоступенчатой системе упаковки генетических молекул. Общая схема компактизации включает пять уровней:
- нуклеосомная нить диаметром 10–11 нм;
- фибрилла 25–30 нм;
- петлевые домены (300 нм);
- волокно толщиной 700 нм;
- хромосомы (1200 нм).
Такая форма организации обеспечивает уменьшение длины исходной молекулы ДНК в 10 тысяч раз.
Нить диаметром 11 нм образована рядом нуклеосом, связанных линкерными участками ДНК. На электронной микрофотографии такая структура напоминает нанизанные на леску бусы. Нуклеосомная нить сворачивается в спираль по типу соленоида, образуя фибриллу толщиной 30 нм. В ее формировании участвует гистон H1.
Соленоидная фибрилла складывается в петли (иначе – домены), которые закрепляются на поддерживающем внутриядерном матриксе. Каждый домен содержит от 30 до 100 тысяч пар нуклеотидов. Такой уровень компактизации характерен для интерфазного хроматина.
Структура толщиной 700 нм образуется при спирализации доменной фибриллы и называется хроматидой. В свою очередь, две хроматиды формируют пятый уровень организации ДНК – хромосому диаметром 1400 нм, которая становится видна на стадии митоза или мейоза.
Таким образом, хроматин и хромосома – это формы упаковки генетического материала, зависящие от жизненного цикла клетки.
Хромосомы
Хромосома состоит из двух идентичных друг другу сестринских хроматид, каждая из которых образована одной суперспирализованной молекулой ДНК. Половинки соединяются особым фибриллярным тельцем, называемым центромерой. Одновременно эта структура является перетяжкой, разделяющей каждую хроматиду на плечи.
В отличие хроматина, представляющего собой структурный материал, хромосома – это дискретная функциональная единица, характеризующаяся не только структурой и составом, но и уникальным генетическим набором, а также определенной ролью в реализации механизмов наследственности и изменчивости на клеточном уровне.
Эухроматин и гетерохроматин
Хроматин в ядре существует в двух формах: менее спирализованной (эухроматин) и более компактной (гетерохроматин). Первая форма соответствует транскрипционно-активным участкам ДНК и поэтому структурирована не так плотно. Гетерохроматин подразделяется на факультативный (может переходить из активной формы в плотную неактивную в зависимости от стадии жизненного цикла клетки и необходимости реализовать те или иные гены) и конститутивный (постоянно уплотнен). Во время митотического или мейотического деления весь хроматин неактивен.
Конститутивный гетерохроматин обнаружен возле центромер и в концевых участках хромосомы. Результаты электронной микроскопии показывают, что такой хроматин сохраняет высокую степень конденсации не только на стадии деления клетки, но и во время интерфазы.
Биологическая роль хроматина
Основная функция хроматина заключается в плотной упаковке большого количества генетического материала. Однако просто уместить ДНК в ядре для жизнедеятельности клетки недостаточно. Необходимо, чтобы эти молекулы должным образом «работали», то есть, могли передавать заключенную в них информацию по системе ДНК-РНК-белок. Кроме этого, клетке нужно распределять генетический материал во время деления.
Устройство хроматина полностью отвечает этим задачам. Белковая часть содержит все необходимые ферменты, а особенности структуры позволяют им взаимодействовать с определенными участками ДНК. Поэтому, второй важной функцией хроматина является обеспечение всех процессов, связанных с реализацией ядерного генома.
Источник: www.syl.ru