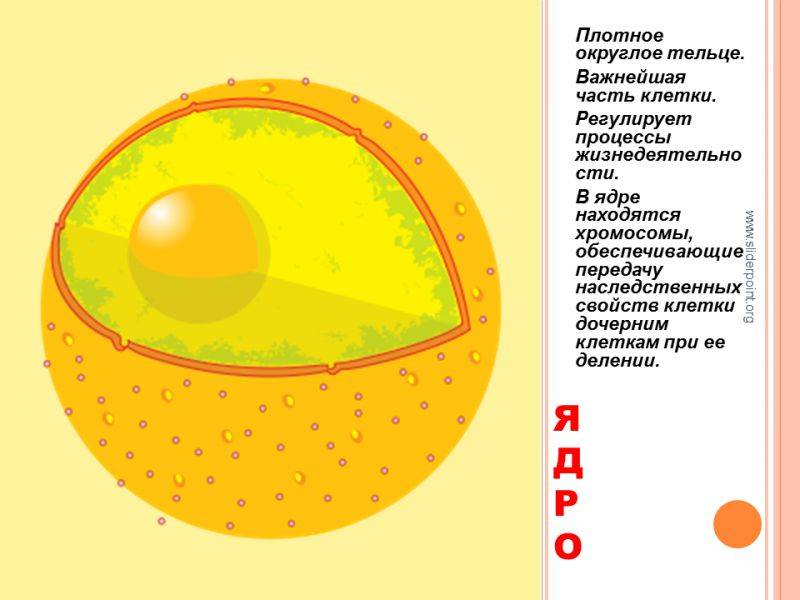
Значение клеточного ядра
Ядро – главное составляющее живой клетки, которое несет наследственную информацию, закодированную набором генов. Оно занимает центральное положение в клетке. Размеры варьируются, форма обычно сферичная или овальная. В диаметре ядро в разных клетках может быть от 8 до 25мкм. Есть исключения, примеру, яйцеклетки рыб имеют ядра диаметром в 1 мм.
Особенности строения ядра
Заполнено ядро жидкостью и несколькими структурными элементами. В нем выделяют оболочку, набор хромосом, нуклеоплазму, ядрышка. Оболочка двухмембранная, между мембранами находится перенуклеарное пространство.

Внешняя мембрана сходна по строению с эндоплазматическим ретикулумом. Она связана с ЭПР, который будто ответвляется от ядерной оболочки. Снаружи на ядре находятся рибосомы.
Внутренняя мембрана прочная, так как в ее состав входит ламина. Она выполняет опорную функцию и служит местом крепления для хроматина.
Мембрана имеет поры, обеспечивающие обменные процессы с цитоплазмой. Ядерные поры состоят из транспортных белков, которые поставляют в кариоплазму вещества путем активного транспорта. Пассивно сквозь поровые отверстия могут пройти только небольшие молекулы. Также каждая пора прикрыта поросомой, которая регулирует обменные процессы в ядре.
Количество ядер в разных по специализации клетках различно. В большинстве случаев клетки одноядерные, но есть ткани, построенные из многоядерных клеток (печеночная или ткань мозга). Есть клетки лишенные ядра – это зрелые эритроциты.
У простейших выделяют два типа ядер: одни отвечают за сохранение информации, другие – за синтез белка.
Ядро может прибывать в состоянии покоя (период интерфазы) или деления. Переходя в интерфазу, имеет вид сферического образования с множеством гранул белого цвета (хроматина). Хроматин бывает двух видов: гетерохроматин и эухроматин.
Эухроматин – это активный хроматин, который сохраняет деспирализированное строение в покоящемся ядре, способен к интенсивному синтезу РНК.
Гетерохроматин – это участки хроматина, которые находятся в конденсированном состоянии. Он может при необходимости переходить в эухроматиновое состояние.
При использовании цитологического метода окрашивания ядра (по Романовскому-Гимзе) выявлено, что гетерохроматин меняет цвет, а эухроматин нет. Хроматин построен из нуклеопротеидных нитей, названных хромосомами. Хромосомы несут в себе основную генетическую информацию каждого человека. Хроматин — форма существования наследственной информации в интерфазном периоде клеточного цикла, во время деления он трансформируется в хромосомы.
Строение хромосом
Каждая хромосома построена из пары хроматид, которые находятся параллельно друг к другу и связаны только в одном месте – центромере. Центромера разделяет хромосому на два плеча. В зависимости от длины плеч выделяют три вида хромосом:
- Равноплечие;
- разноплечие,
- одноплечие.
Некоторые хромосомы имеют дополнительный участок, который крепится к основному нитевидными соединениями – это сателлит. Сателлиты помогают идентифицировать разные пары хромосом.
Метафазное ядро представляет собой пластинку, где располагаются хромосомы. Именно в эту фазу митоза изучается количество и строение хромосом. Во время метафазы сестринские хромосомы двигаются в центр и распадаются на две хроматиды.
Строение ядрышка
В ядре также находится немембранное образование — ядрышко. Ядрышки представляют собой уплотненные, округлые тельца, способные преломлять свет. Это основное место синтеза рибосомальной РНК и необходимых белков.
Число ядрышек различно в разных клетках, они могут объединяться в одно крупное образование или существовать отдельно друг от друга в виде мелких частиц. При активации синтетических процессов объем ядрышка увеличивается. Оно лишено оболочки и находится в окружении конденсированного хроматина. В ядрышке также содержатся металлы, в большей мере цинк. Таким образом, ядрышко – это динамичное, меняющееся образование, необходимое для синтеза РНК и транспорта ее в цитоплазму.
Нуклеоплазма заполняет все внутреннее пространство ядра. В нуклеоплазме находится ДНК, РНК, протеиновые молекулы, ферментативные вещества.
Функции ядра в клетке
- Принимает участие в синтезе белка, рибосомной РНК.
- Регулирует функциональную активность клетки.
- Сохранение генетической информации, точная ее репликация и передача потомству.
Роль и значение ядра
Ядро является главным хранилищем наследственной информации и определяет фенотип организма. В ядре ДНК существует в неизмененном виде благодаря репарационным ядерным ферментам, которые способны ликвидировать поломки и мутации. Во время клеточного деления ядерные механизмы обеспечивают точное и равномерное расхождение генетической информации в дочерние клетки.
Источник: animals-world.ru
Содержание
- 1 Тонкая структура клеточного ядра
- 1.1 Хроматин
- 1.2 Ядерная оболочка, ядерная ламина и ядерные поры (кариолемма)
- 1.3 Ядрышко
- 1.4 Ядерный матрикс
- 2 Эволюционное значение клеточного ядра
- 3 Происхождение ядра
- 4 Примечания
- 5 Ссылки
Тонкая структура клеточного ядра
Схема строения клеточного ядра.
Хроматин
Огромная длина молекул ДНК эукариот предопределила появление специальных механизмов хранения, репликации и реализации генетического материала. Хроматином называют молекулы хромосомной ДНК в комплексе со специфическими белками, необходимыми для осуществления этих процессов. Основную массу составляют «белки хранения», так называемые гистоны. Из этих белков построены нуклеосомы — структуры, на которые намотаны нити молекул ДНК. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: H2A, H2B, H3 и H4. В одну нуклеосому входят по два белка каждого типа — всего восемь белков. Гистон H1, более крупный чем другие гистоны, связывается с ДНК в месте ее входа на нуклеосому. Нуклеосома вместе с H1 называется хроматосомой.
Схема, показывающая цитоплазму, вместе с ее компонентами (или органеллами), в типичной животной клетке. Органеллы:
(1) Ядрышко
(2) Ядро
(3) рибосома (маленькие точки)
(4) Везикула
(5) шероховатый эндоплазматический ретикулум (ER)
(6) Аппарат Гольджи
(7) Цитоскелет
(8) Гладкий эндоплазматический ретикулум
(9) Митохондрия
(10) Вакуоль
(11) Цитоплазма
(12) Лизосома
(13) Центриоль и Центросома
Нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином, он хорошо виден под микроскопом. ДНК, находящаяся в гетерохроматине, не транскрибируется, обычно это состояние характерно для незначимых или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки. Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием и фосфорилированием.
Считается, что в ядре существуют так называемые функциональные домены хроматина(ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». К сожалению, вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины.
Ядерная оболочка, ядерная ламина и ядерные поры (кариолемма)
От цитоплазмы ядро отделено ядерной оболочкой, образованной за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. Ламины прикрепляются к внутренней мембране ядерной оболочки при помощи заякоренных в ней трансмембранных белков — рецепторов ламинов.
некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Пора не является дыркой в ядре, а имеет сложную структуру, организованную несколькими десятками специализированных белков — нуклеопоринов. Под электронным микроскопом она видна как восемь связанных между собой белковых гранул с внешней и столько же с внутренней стороны ядерной оболочки.
Различные структуры клеточного ядра видны из-за накопления в них зелёного флуоресцентного белка
Ядрышко
Основная статья: Ядрышко
Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специальные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации (NoLS, от англ. Nucleolus Localization Signal). Следует отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда неспецифически.
Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц.
Ядерный матрикс
Основная статья: Ядерный скелет
Ядерным матриксом некоторые исследователи называют нерастворимый внутриядерный каркас. Считается, что матрикс построен преимущественно из негистоновых белков, формирующих сложную разветвленную сеть, сообщающуюся с ядерной ламиной. Возможно, ядерный матрикс принимает участие в формировании функциональных доменов хроматина. В геноме клетки имеются специальные незначащие А-Т-богатые участки прикрепления к ядерному матриксу (англ. S/MAR — Matrix/Scaffold Attachment Regions), служащие, как предполагается, для заякоривания петель хроматина на белках ядерного матрикса. Впрочем, не все исследователи признают существование ядерного матрикса.
Принципиальная схема реализации генетической информации у про- и эукариот.
ПРОКАРИОТЫ. У прокариот синтез белка рибосомой (трансляция) пространственно не отделен от транскрипции и может происходить ещё до завершения синтеза мРНК РНК-полимеразой.
окариотические мРНК часто полицистронные, то есть содержат несколько независимых генов.
ЭУКАРИОТЫ. мРНК эукариот синтезируется в виде предшественника, пре-мРНК, претерпевающего затем сложное стадийное созревание — процессинг, включающий присоединение кэп-структуры к 5′-концу молекулы, присоединение нескольких десятков остатков аденина к ее 3′-концу (полиаденилирование), выщепление незначащих участков — интронов и соединение друг с другом значащих участков — экзонов (сплайсинг). При этом соединение экзонов одной и той же пре-мРНК может проходить разными способами, приводя к образованию разных зрелых мРНК, и в конечном итоге разных вариантов белка (альтернативный сплайсинг). Только мРНК, успешно прошедшая процессинг, экспортируется из ядра в цитоплазму и вовлекается в трансляцию.
Эволюционное значение клеточного ядра
Основное функциональное отличие клеток эукариот от клеток прокариот заключается в пространственном разграничении процессов транскрипции (синтеза матричной РНК) и трансляции (синтеза белка рибосомой), что дает в распоряжение эукариотической клетки новые инструменты регуляции биосинтеза и контроля качества мРНК.
В то время, как у прокариот мРНК начинает транслироваться еще до завершения ее синтеза РНК-полимеразой, мРНК эукариот претерпевает значительные модификации (так называемый процессинг), после чего экспортируется через ядерные поры в цитоплазму, и только после этого может вступить в трансляцию. Процессинг мРНК включает несколько элементов.
Из предшественника мРНК (пре-мРНК) в ходе процесса, называемого сплайсингом вырезаются интроны — незначащие участки, а значащие участки — экзоны соединяются друг с другом. Причем экзоны одной и той же пре-мРНК могут быть соединены несколькими разными способами (альтернативный сплайсинг), так что один предшественник может превращаться в зрелые мРНК нескольких разных видов. Таким образом, один ген может кодировать сразу несколько белков.
Кроме того, интрон-экзонная структура генома, практически невозможная у прокариот (так как рибосомы смогут транслировать незрелые мРНК), дает эукариотам определенную эволюционную мобильность. Учитывая протяженность интронных участков, рекомбинация между двумя генами зачастую сводится к обмену экзонами. Благодаря тому, что экзоны часто соответствуют функциональным доменам белка, участки получившегося в результате рекомбинации «гибрида», зачастую сохраняют свои функции. В то же время у прокариот рекомбинация между генами невозможна без разрыва в значащей части, что безусловно уменьшает шансы на то, что получившийся белок будет функционален.
Модификациям подвергаются концы молекулы мРНК. К 5′ -концу молекулы прикрепляется 7-метилгуанин (так называемый кэп). К 3′-концу нематрично присоединяются несколько десятков остатков аденина (полиаденирование).
Процессинг мРНК тесно сопряжен с синтезом этих молекул и необходим для контроля качества. Непроцессированная или не полностью процессированная мРНК не сможет выйти из ядра в цитоплазму или будет нестабильна и быстро деградирует. У прокариот нет таких механизмов контроля качества, и из-за этого прокариотические мРНК имеют меньший срок жизни — нельзя допустить, чтобы неправильно синтезированная молекула мРНК, если такая появится, транслировалась в течение долгого времени.
Происхождение ядра
Клеточное ядро является важнейшей чертой эукариотических организмов, отличающей их от прокариот и архей. Несмотря на значительный прогресс в цитологии и молекулярной биологии, происхождение ядра не выяснено и является предметом научных споров. Выдвинуто 4 основных гипотезы происхождения клеточного ядра, но ни одна из них не получила широкой поддержки.
Гипотеза, известная как «синтропная модель», предполагает что ядро возникло в результате симбиотических взаимоотношений между археей и бактерией (ни археи, ни бактерии не имеют оформленных клеточных ядер). По этой гипотезе, симбиоз возник, когда древняя архея (сходная с современными метаногенными археями), проникла в бактерию (сходную с современными Миксобактериями). Впоследствии архея редуцировалась до клеточного ядра современных эукариот. Эта гипотеза аналогична практически доказанным теориям происхождения митохондрий и хлоропластов, которые возникли в результате эндосимбиоза прото-эукариот и аэробных бактерий. Доказательством гипотезы является наличие одинаковых генов у эукариот и архей, в частности генов гистонов. Также миксобактерии быстро передвигаются, могут образовывать многоклеточные структуры и имеют киназы и G-белки, близкие к эукариотическим.
Согласно второй гипотезе, прото-эукариотическая клетка эволюционировала из бактерии без стадии эндосимбиоза. Доказательством модели является существование современных бактерий из отряда Planctomycetes, которые имеют ядерные структуры с примитивными порами и другие клеточные компартменты, ограниченные мембранами (ничего похожего у других прокариот не обнаружено).
Согласно гипотезе вирусного эукариогенеза, окруженное мембраной ядро, как и другие эукариотические элементы, произошли вследствие инфекции прокариотической клетки вирусом. Это предположение основывается на наличии общих черт у эукариот и некоторых вирусов, а именно геноме из линейных цепей ДНК, кэпировании мРНК и тесном связывании генома с белками (гистоны эукариот принимаются аналогами вирусных ДНК-связывающих белков). По одной версии, ядро возникло при фагоцитировании (поглощении) клеткой большого ДНК-содержащего вируса. По другой версии, эукариоты произошли от древних архей, инфицированных поксвирусами. Эта гипотеза основана на сходстве ДНК-полимеразы современных поксвирусов и эукариот. Также предполагается, что нерешенный вопрос о происхождении пола и полового размножения может быть связан с вирусным эукариогенезом.
Наиболее новая гипотеза, названная экзомембранной гипотезой, утверждает, что ядро произошло от одиночной клетки, которая в процессе эволюции выработала вторую внешнюю клеточную мембрану; первичная клеточная мембрана после этого превратилась в ядерную мембрану, и в ней образовалась сложная система поровых структур (ядерных пор) для транспорта клеточных компонентов, синтезированных внутри ядра.
Примечания
- ↑ Pennisi E. (2004). «Evolutionary biology. The birth of the nucleus». Science 305 (5685): 766–768. DOI:10.1126/science.305.5685.766. PMID 15297641.
- ↑ Margulis, Lynn. Symbiosis in Cell Evolution. — San Francisco: W. H. Freeman and Company, 1981. — P. 206–227. — ISBN 0-7167-1256-3.
- ↑ Lopez-Garcia P, Moreira D. (2006). «Selective forces for the origin of the eukaryotic nucleus». Bioessays 28 (5): 525–533. DOI:10.1002/bies.20413. PMID 16615090.
- ↑ Fuerst JA. (2005). «Intracellular compartmentation in planctomycetes». Annu Rev Microbiol. 59: 299–328. DOI:10.1146/annurev.micro.59.030804.121258. PMID 15910279.
- ↑ Bell PJ (September 2001). «Viral eukaryogenesis: was the ancestor of the nucleus a complex DNA virus?». J. Mol. Evol. 53 (3): 251–6. DOI:10.1007/s002390010215. PMID 11523012.
- ↑ Takemura M (2001). «Poxviruses and the origin of the eukaryotic nucleus». J Mol Evol 52 (5): 419–425. DOI:10.1007/s002390010171. PMID 11443345.
- ↑ Villarreal L, DeFilippis V (2000). «A hypothesis for DNA viruses as the origin of eukaryotic replication proteins». J Virol 74 (15): 7079–7084. DOI:10.1128/JVI.74.15.7079-7084.2000. PMID 10888648.
- ↑ Bell PJ (November 2006). «Sex and the eukaryotic cell cycle is consistent with a viral ancestry for the eukaryotic nucleus». J. Theor. Biol. 243 (1): 54–63. DOI:10.1016/j.jtbi.2006.05.015. PMID 16846615.
- ↑ de Roos AD (2006). «The origin of the eukaryotic cell based on conservation of existing interfaces». Artif Life 12 (4): 513–523.. DOI:10.1162/artl.2006.12.4.513. PMID 16953783.
Ссылки
| Клеточное ядро на Викискладе? |
Molecular Biology Of The Cell, 4е издание, 2002 г. — учебник по молекулярной биологии на английском языке
| Клеточное ядро | |
|---|---|
| Ядерная мембрана • Ядерная ламина • Ядерные поры (см. также нуклеопорины) • Ядрышко (см. также тельце Кахаля) • Хроматин • Ядерные тельца • Ядерный скелет • Нуклеоплазма • Нуклеозоль |
| Органоиды эукариотической клетки | ||
|---|---|---|
|
Акросома • Аппарат Гольджи • Вакуоль • Везикула • Клеточная мембрана • Клеточная стенка • Лизосома • Меланосома • Миофибриллы • Митохондрия • Пероксисома • Пластиды • Реснички/Жгутики • Рибосома • Сократительная вакуоль • Стигма • Хлоропласты • Центриоль • Центросома • Цитоплазма • Эндоплазматический ретикулум • Ядро • Ядрышко |
||
клеточное ядро атома, клеточное ядро вікіпедія, клеточное ядро земли, клеточное ядро клетки
Клеточное ядро Информацию О
Источник: www.turkaramamotoru.com
Основное функциональное отличие клеток эукариот от клеток прокариот заключается в пространственном разграничении процессов транскрипции (синтеза матричной РНК) и трансляции (синтеза белка рибосомой), что дает в распоряжение эукариотической клетки новые инструменты регуляции биосинтеза и контроля качества мРНК.
В то время, как у прокариот мРНК начинает транслироваться еще до завершения ее синтеза РНК-полимеразой, мРНК эукариот претерпевает значительные модификации (так называемый процессинг), после чего экспортируется через ядерные поры в цитоплазму, и только после этого может вступить в трансляцию. Процессинг мРНК включает несколько элементов.
Из предшественника мРНК (пре-мРНК) в ходе процесса, называемого сплайсингом вырезаются интроны — незначащие участки, а значащие участки — экзоны соединяются друг с другом. Причем экзоны одной и той же пре-мРНК могут быть соединены несколькими разными способами (альтернативный сплайсинг), так что один предшественник может превращаться в зрелые мРНК нескольких разных видов. Таким образом, один ген может кодировать сразу несколько белков.
Кроме того, интрон-экзонная структура генома, практически невозможная у прокариот (так как рибосомы смогут транслировать незрелые мРНК), дает эукариотам определенную эволюционную мобильность. Учитывая протяженность интронных участков, рекомбинация между двумя генами зачастую сводится к обмену экзонами. Благодаря тому, что экзоны часто соответствуют функциональным доменам белка, участки получившегося в результате рекомбинации «гибрида», зачастую сохраняют свои функции. В то же время у прокариот рекомбинация между генами невозможна без разрыва в значащей части, что безусловно уменьшает шансы на то, что получившийся белок будет функционален.
Модификациям подвергаются концы молекулы мРНК. К 5′ -концу молекулы прикрепляется 7-метилгуанин (так называемый кэп). К 3′-концу нематрично присоединяются несколько десятков остатков аденина (полиаденирование).
Процессинг мРНК тесно сопряжен с синтезом этих молекул и необходим для контроля качества. Непроцессированная или не полностью процессированная мРНК не сможет выйти из ядра в цитоплазму или будет нестабильна и быстро деградирует. У прокариот нет таких механизмов контроля качества, и из-за этого прокариотические мРНК имеют меньший срок жизни — нельзя допустить, чтобы неправильно синтезированная молекула мРНК, если такая появится, транслировалась в течение долгого времени.
Источник: studopedia.su