Дрожжи прокариоты или эукариоты
Морфология и классификация прокариотов и эукариотов. Генетика микроорганизмов
министерство образования и науки рф
ФИЛИАЛ ФГБОУ ВПО «МОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ТЕХНОЛОГИЙ И УПРАВЛЕНИЯ им. К.Г. РАЗУМОВСКОГО» в г. ВЯЗЬМЕ СМОЛЕНСКОЙ обл.
Реферат
Дисциплина: Основы микробиологии
Тема: Морфология и классификация прокариотов и эукариотов. Генетика микроорганизмов.
Студент: Сергеева Светлана
Руководитель: Дмитриев Анатолий Иванович
Вязьма 2014
Содержание
Введение
.Морфология и классификация прокариотов (бактерий)
.Строение клетки прокариотов
.Биоразнообразие прокариотов
.Морфология и классификация эукариотов (микроскопических грибов и дрожжей)
.Строение клетки эукариотов
.Дрожжи. Общая характеристика
.Генетика микроорганизмов
Заключение
Список использованной литературы
Введение
Морфология микроорганизмов изучает их внешний вид, форму и особенности строения, способность к движению, спорообразованию, способы размножения. Морфологические признаки играют большую роль в распознавании и классификации микроорганизмов. С древнейших времен живой мир делили на два царства: царство растений и царство животных. Когда был открыт мир микроорганизмов, то их выделили в отдельное царство.
1. Морфология и классификация прокариотов (бактерий)
Прокариоты (бактерии и актиномицеты). Бактерии (прокариоты)-это большая группа микроорганизмов (около 1600 видов), большинство из которых одноклеточные. Форма и размеры бактерий. Основные формы бактерий: шаровидная, палочковидная и извитая. Шаровидные бактерии — кокки имеют обычную форму шара, встречаются уплощенные, овальной или бобовидной формы. Кокки могут быть в виде клеток одиночных — монококки (микрококки) или соединенных в различных сочетаниях: попарно — диплококки, по четыре клетки — тетракокки, в виде более или менее длинных цепочек — стрептококки, а также в виде скоплений кубической формы (в виде пакетов) из восьми клеток, расположенных в два яруса один над другим, — сарцины. Встречаются скопления неправильной формы, напоминающие грозди винограда, — стафилококки. Палочковидные бактерии могут быть одиночными или соединенными попарно — диплобактерии, цепочками по три-четыре и более клеток — стрептобактерии. Соотношения между длиной и толщиной палочек бывают самыми различными.
Извитые, или изогнутые, бактерии различаются длиной, толщиной и степенью изогнутости. Палочки, слегка изогнутые в виде запятой, называют вибрионами, палочки с одним или несколькими завитками в виде штопора — спириллами, а тонкие палочки с многочисленными завитками — спирохетами. Благодаря использованию электронного микроскопа для изучения микроорганизмов в естественных природных субстратах были обнаружены бактерии, имеющие особую форму клеток замкнутого или разомкнутого кольца (тороиды) ; с выростами (простеками); червеобразной формы — длинные с загнутыми очень тонкими концами; а также в виде шестиугольной звезды.
2. Строение клетки прокариотов
Цитоплазма полужидкая, вязкая, коллоидная масса, в нее входят белки, нуклеиновые к-ты, липиды, вода; Цитоплазматическая мембрана обладает полупроницаемостью, богата липидами и ферментами; Рибосомы — синтез белка; Мезосомы — энергетические процессы: окисление орг. в-в, синтез энергозапасающих в-в (АТФ); различные включения, являющиеся запасными питательными в-вами (гликоген, волютин); Ядро отсутствует, но имеется большое кол-во ядерного в-ва, в частности дезоксирибонуклеиновой к-ты (ДНК); Слизистый чехол (полисахариды) — необязательный компонент клетки, предохраняет от высыхания, от мех-ого повреждения, д-ия хим. агентов и лекарственных в-в; Клеточная стенка (также необязательна) состоит из муреинового комплекса (гликопептиды).
В мире прокариотов встречаются разные варианты структур и способов питания; прокариоты проявляют и элементарные, и довольно сложные поведенческие реакции.Бактерии и архей достигли высшего совершенства на низшем уровне биологической организации, соответствующем клетке-организму:
— они используют все структурные детали в рамках прокариотного морфотипа и даже могут образовывать многоклеточные ассоциации;
— они ассимилируют все биологически доступные типы энергии и конструктивных субстратов;
— они занимают все ниши, пригодные для существования живых организмов.
Широкая норма реакции прокариотов позволила им за миллиарды лет биологической эволюции не только выжить в агрессивной абиотической среде, но и дает возможность успешно конкурировать с ядерными химерами.
3. Биоразнообразие прокариотов
Биоразнообразие прокариотов — это набор фенотипов, который Г. А. Заварзин назвал «пространством логических возможностей». Структурные и функциональные признаки прокариотов могут образовывать почти любые сочетания; запрещенные комбинации очень редки (например, плавание при помощи жгутиков несовместимо со скользящей подвижностью, а оксигенная фототрофия — со способностью образовывать эндоспоры).
Биоразнообразие прокариотов в первую очередь выражается в разных типах питания. Оно дополняется цитоморфологическими различиями, а также разнообразием форм поведения. Одна из задач систематики прокариот, как и других живых существ, распределение множества организмов по группам. Для характеристики организмов используют разнообразные признаки: морфологические, цитологические, культуральные, физиологические, биохимические, иммунологические, генетические и др.
В настоящий момент описано около 5,2 × 10³ законных видов бактерий и архей. Это не более 0,3% видового разнообразия ядерных химер.
Однако законные виды прокариотов лишь вершина айсберга. Истинное биоразнообразие прокариотов состоит из четырех качественно и количественно неравноценных пулов.
. Культивируемые объекты. Фенотипически их можно охарактеризовать до той степени, насколько позволяют современные возможности многофакторного анализа.
. Некультивируемые объекты. Сведения об их фенотипе в основном ограничиваются морфологией. При переносе на питательную среду такие объекты испытывают стресс, теряют способность к размножению и переходят в латентное состояние, что не позволяет установить их физиологические свойства.
. Фантомные объекты. Их фенотип неизвестен. Такие объекты выявляются косвенно по присутствию в воде, почве и других природных материалах уникальных последовательностей генов рРНК, которые амплифицируются с помощью PCR-реакции, секвенируются и сравниваются с банком данных.
О существовании огромного разнообразия фантомных форм бактерий и архей стало известно с 1985 г., когда группа американских бактериологов под руководством Нормана Пейса (N.Pace) сообщила свои первые результаты молекулярно-биологического анализа «природной» ДНК.
. Криптические объекты. В отличие от актуалистических, они существуют только «на кончике пера», и их фенотипы можно прогнозировать, комбинируя признаки уже описанных объектов. Современные исследователи дают грубую оценку ожидаемого числа новых генотипов, аппроксимируя график их выявления на протяжении последних лет.
Классификация прокариотов представляет собой один из разделов биологической классификации, со своей спецификой и проблематикой.
Прокариоты — они размножаются исключительно вегетативным способом и теоретически могут проникнуть в любой ареал. Кроме того, им свойствен промискуитет, что выражается в «горизонтальном» переносе генов между неродственными объектами при помощи молекулярных векторов — вирусов и плазмид.
Вид у прокариотов — это совокупность генетически родственных клональных популяций из разных местообитаний или группа генетически родственных коллекционных штаммов.
В таком определении вида важная роль принадлежит понятиям «клон» и «штамм». Эти термины не относятся к таксономическим категориям, хотя они отражают генетическую обособленность культивируемого объекта; в частности, при отсутствии видового эпитета его место занимает регистрационный номер штамма.
Размножение прокариотов происходит агамным путем, т. е. посредством клонирования. В природных популяциях или в лабораторных культурах клонирование происходит спонтанно, и в итоге образуется смесь клонов. Клон у прокариотов — это потомство индивидуальной клетки или индивидуальной квази-многоклеточной структуры (трихома) при размножении в природе или в лаборатории.
С помощью понятия «коллекционный штамм», или просто «штамм» регистрируется самостоятельный акт выделения микроорганизма из природного материала.
Штаммы сохраняются в коллекциях в виде пересеваемых (развивающихся или покоящихся) культур, а также в анабиотическом состоянии.
Коллекционный штамм — это популяция микроорганизма, ставшая объектом хранения или исследования в результате однократного выделения из природного материала.
К перечисленным понятиям необходимо добавить понятие «культура», являющееся одним из важнейших в фундаментальной и прикладной микробиологии.
Как провести границу между видами прокариотов?
Единого фенотипического критерия для этого не существует. В зависимости от выраженности цитологических, физиологических или экологических различий то или иное из них выходит на передний план. По такому же принципу разграничиваются и таксономические группы более высокого ранга.
Генотипический критерий проще стандартизировать, хотя в этом случае границы видов становятся формальными. Их устанавливают путем консенсуса, вторично оценивая молекулярно-генетические различия у «хороших», т. е. четко дифференцируемых и общепризнанных видов.
Согласно рекомендации Международного комитета по систематике бактерий, разными видами являются штаммы, у которых:
содержание ГЦ-пар в ДНК различается больше, чем на 5%;
степень ДНК-ДНК гибридизации меньше, чем 30%;
различие нуклеотидных последовательностей 16S рРНК больше 2%.
Род как центральный таксон в систематике прокариотов; типовой род. Систематика ядерных организмов, особенно тех, которые размножаются половым путем, опирается на фундамент вида. Прокариоты размножаются только агамно, и их популяции не разделены географическими барьерами.
Поэтому фундаментом систематики прокариотов является не вид, а таксон более высокого ранга, который сильнее дивергировал от аналогичного таксона, чем дивергируют виды. В данном случае это род, состоящий из одного или нескольких видов.
Парадоксальность филогенетической системы прокариотов дает о себе знать на уровне рода, а также на уровне таксономических групп более высокого ранга — семейства, порядка, класса, филы и домена.
Из-за «парадоксальной» структуры большинства семейств в систематике прокариотов специфический смысл приобретает такое понятие, как «типовой род». Однако «типовой» не значит типичный, т. е. наиболее характерный. При выборе типового рода, как правило, учитывается приоритет, и чаще всего предпочтение отдается самому раннему диагнозу. Таким образом, типовой род — это «полномочный представитель» семейства. Данный алгоритм применяется и к таксономическим группам более высокого ранга (от семейства до филы).
Названия прокариотов. В большинстве случаев в систематике прокариотов используется бинарная номенклатура, т. е. родовое имя сопровождается видовым эпитетом. Отдельно от других стоят цианобактерии, в систематике которых категория вида используется лишь в виде исключения.
Название вида или таксона более высокого ранга считается законным, если оно входит в «Список признанных названий бактерий» 1980 г.
Новые законные описания должны быть опубликованы в «International Journal of Systematic Bacteriology» (переименован в «International Journal of Systematic and Evolutionary Microbiology» в 2001 г.). Незаконное родовое название вместе с видовым эпитетом ставится в кавычки так же, как нелегитимное название таксона более высокого ранга (например, класс «Alphaproteobacteria»).
В отличие от родовых названий животных и растений, которые в большинстве случаев являются нейтральными именами или отражают морфологические признаки, названия бактерий и архей характеризуют в первую очередь их физиологические свойства. Наряду с этим используются разные смысловые элементы, в том числе произвольного характера.
Родовое имя и видовой эпитет могут содержать по одному смысловому элементу, например Bacillus subtilis («палочка, тонкая») или по 2-4 смысловых элемента, например Ectothiorhodospira halochloris («окруженная серой пурпурная спирилла, имеет отношение к поваренной соли»).
В связи с тем, что многие архей занимают экстремальные ниши, сложилась традиция давать им названия в терминах потустороннего мира, например Acidianus infemus (лат. infernum — ад), Methanotorris igneus (лат. torris — головня и ignis — огонь), Pyrococcus furiosus (лат. furia — ярость) , Stygioglobus azoricus (лат. Styx — река в подземном царстве) , Natronococcus occultus (лат. occultus — потаенный) и т.д.
Прокариоты получают имена согласно правилам Международного кодекса номенклатуры бактерий. Не запрещено, хотя и не рекомендуется увековечивать персон, не имеющих отношения к микробиологии, а также неэтично называть объекты в честь самого себя, своих партнеров и прочих заинтересованных лиц.
В случае некультивируемых форм часто отсутствует информация о важнейших фенотипических признаках, например типе питания. Это делает традиционную классификацию невозможной. Такие организмы не имеют родового имени и видового эпитета, и их обозначают как Candidatus с добавлением предварительного названия, например Candidatus «Scalindua sorokinii».
4. Морфология и классификация эукариотов (микроскопических грибов и дрожжей)
Многие грибы употребляют в пищу, используют в промышленных условиях для получения органических кислот витаминов, ферментов, антибиотиков.
Многочисленные грибы, развивающиеся на пищевых продуктах, промышленных материалах и изделиях, вызывают их порчу и разрушение. Некоторые из них способны вырабатывать токсические для человека и животных вещества — микотоксины. Многие грибы поражают культурные растения в процессе их вегетации, нанося большой урон сельскому хозяйству. Есть грибы, вызывающие заболевания человека и животных.
Строение тела гриба. Вегетативное тело большинства грибов представляет собой грибницу, или мицелий, состоящий из ветвящихся нитей — гиф. Такие грибы называют мицелиальными (или плесенями).
С помощью сканирующего электронного микроскопа установлено (А. А. Кудряшова), что гифы грибов различаются внешним видом, строением стенки, длиной, толщиной и рельефом поверхности. Они могут быть прямыми, изогнутыми, спиралевидными, со вздутиями или утолщениями, с углублениями и короткими отростками "корешками", служащими для прикрепления к субстрату. Поверхность гиф бывает с шипами, гладкой, сетчатой, волокнистой, местами складчатой. Диаметр гиф колеблется от 2 до 25 мкм и более. Гифы растут вершиной или концами разветвлений, поэтому их клетки неоднородны по длине. Мицелий развивается частично в субстрате (субстратный мицелий), пронизывая его и высасывая из него воду и питательные вещества, а частично — на поверхности субстрата (воздушный мицелий) в виде пушистых, паутинообразных или тонких налетов, пленок. Гифы отдельных грибов могут плотно переплетаться и даже срастаться между собой. У некоторых грибов гифы соединяются параллельно в тяжи, достигающие иногда нескольких метров в длину, по ним притекают питательные вещества.
Немногие грибы не имеют мицелия. Это некоторые представители низших грибов, а также дрожжи, которые являют собой одиночные округлые или удлиненные клетки.
Грибы относятся к домену Eukarya, царству Fungi (Mycota, Mycetes). Недавно грибы и простейшие были разделены на самостоятельные царства: царство Eumycota (настоящие грибы), царство Chromista и царство Protozoa. Некоторые микроорганизмы, ранее считавшиеся грибами или простейшими, были перемещены в новое царство Chromista(хромовики). Грибы — многоклеточные или одноклеточные нефотосинтезирующие (бесхлорофильные) эукариотические микроорганизмы с толстой клеточной стенкой. Они имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и многослойную ригидную клеточную стенку, состоящую из нескольких типов полисахаридов (маннаны, глюканы, целлюлоза, хитин), а также белка, липидов и др. Некоторые грибы образуют капсулу. Цитоплазматическая мембрана содержит гликопротеины, фосфолипиды и эргостеролы (в отличие от холестерина — главного стерола тканей млекопитающих). Большинство грибов — облигатные или факультативные аэробы.
Грибы широко распространены в природе, особенно в почве. Некоторые грибы содействуют производству хлеба, сыра, молочнокислых продуктов и алкоголя. Другие грибы продуцируют антимикробные антибиотики (например, пенициллин) и иммунодепрессивные лекарства (например, циклоспорин). Грибы используют генетики и молекулярные биологи для моделирования различных процессов. Фитопатогенные грибы наносят значительный ущерб сельскому хозяйству, вызывая грибковые болезни злаковых растений и зерна. Инфекции, вызываемые грибами, называются микозами. Различают гифальные и дрожжевые грибы.
. Строение клетки эукариотов
Цитоплазма — коллоидный р-р аминокислот, углеводов, минеральных солей в воде (50-60% объема клетки); вязкость превышает вязкость воды в 800 раз; Митохондрии явл. «силовыми станциями» клетки; Рибосомы — органеллы, в которых происходит синтез белка; Лизосомы содержат ферменты, расщепляют чужие биополимеры, обязательно окружены мембраной (Автолиз — самопереваривание клетки, когда клетка старая: разрушаются мембраны); Аппарат Гольджи — упаковка ненужных в-в и транспорт их из клетки через мембрану; Эндоплазматическая сеть связывает ядро с рибосомами, это сложная сис-ма взаимосвязанных каналов, пронизывающих всю толщу клеток (гладкая, шероховатая — связана с рибосомами). Клетка представляет собой сис-му из 2х несмешивающихся между собой фаз: водной (цитоплазма со всеми переходами) и мембранной сис-мой (относительно жидкая, липопротеиновая фаза, которая пронизывает всю цитоплазму).
. Дрожжи. Общая характеристика
Дрожжи являются одноклеточными неподвижными микроорганизмами, широко распространенными в природе; они встречаются в почве, на листьях, стеблях и плодах растений, в разнообразных пищевых субстратах растительного и животного происхождения.
Широкое использование дрожжей в промышленности основано на их способности вызывать спиртовое брожение.
Форма и строение дрожжевой клетки. Форма клеток дрожжей чаще округлая, овально-яйцевидная или эллиптическая, реже цилиндрическая и лимоновидная. Встречаются дрожжи особой формы — серповидные, игловидные, стреловидные, треугольные. Размеры дрожжевых клеток обычно не превышают 10-15 мкм.
Форма и размеры дрожжей могут заметно изменяться в зависимости от условий развития, а также возраста клеток.
Строение клетки дрожжей сходно со строением клетки грибов. Дрожжи обладают всеми основными структурами, характерными для эукариотного типа клетки (ядро, отграниченное от цитоплазмы, эндоплазматическая сеть, аппарат Гольджи, лизосомы, митохондрии, рибосомы, вакуоли). В качестве запасных питательных веществ в клетках обнаруживаются капельки жира, гранулы гликогена, волютина.
7. Генетика микроорганизмов
бактерия спорообразование дрожжевой клетка
Преимущество микроорганизмов над другими организмами состоит, прежде всего, в высокой скорости размножения, гаплоидности и большой разрешающей способности методов генетического анализа этих организмов. Формирование на питательных средах многомиллиардных популяций бактерии в течение суток позволяет проводить быстрый и точный анализ происходящих в них количественных и качественных изменений. Сравнительная простота постановки эксперимента обусловливает эффективность селективного анализа микробной популяции и выделение единичных особей, мутировавших с частотой 10 и выше. Наконец, гаплоидность бактерий, имеющих в отличие от эукариотов одну хромосому, т.е. одну группу сцепления генов, обусловливает отсутствие у них явление доминантности, что способствует быстрому выявлению мутировавших генов.
Материальной основой наследственности, определяющей генетические свойства всех организмов, в том числе бактерий и вирусов, является хромосома, представляющая собой огромную молекулу ДНК в виде двойной спирали, замкнутой в кольцо. Она и является носителем генетической информации. Вдоль хромосомы линейно располагаются генифунхционально неоднородные генетические детерминанты, т.е. фрагменты молекулы ДНК", контролирующие синтез одного белка или пептида. Совокупность генов, локализованных в гаплоидном (одинарном) наборе хромосом данного организма, называется геномом или генотипом. В более широком смысле генотип — совокупность всех наследственных факторов организма, как ядерных (геном), так и неядерных, вне хромосомных (пластидные и цитоплазматические) наследственных факторов. Генотип — носитель наследственной информации, передаваемой от поколения к поколению. Он представляет собой систему, контролирующую фенотип организма. Фенотипом является внешний вид организма со всеми внешними и внутренними признаками. Фенотип бактерий есть результат взаимодействия ее генотипа и среды. Появление в организме новых наследственно передаваемых свойств может быт вызвано разными типами изменений в генетическом аппарате. Источником последовательной изменчивости могут быть мутации генов. Под мутацией понимают внезапные естественные (спонтанные) или вызванные искусственно (индуцированные) стойкие изменения наследственных структур (генов, хромосом), а также обусловленные ими различные изменения свойств и признаков организма. При этом в двойной спирали ДНК могут происходить следующие изменения:
замещение пары оснований, имевшихся в исходной молекуле ДНК другой парой;
выпадение пары оснований из молекулы ДНК;
внедрение новой пары оснований в молекулу ДНК;
инверсия — поворот нескольких пар оснований на 180 ° С.
Таким образом, в основе мутации лежат молекулярные изменения в хромосоме. Мутации у бактерий выявляются при наследственных изменениях любого признака микроба. Классифицируют мутации по происхождению, локализации на хромосоме клетки и другим признакам. Подобное деление носит условный характер. К спонтанным мутациям относят такие мутации, причину возникновения которых трудно и даже невозможно связать с действием определенного фактора (мутагена). Они образуются самопроизвольно в любой популяции микроорганизмов без видимого внешнего воздействия. Спонтанные мутанты образуются до воздействия селекционирующих факторов, которые лишь отбирают признаки, сформировавшиеся в популяции мутировавших бактериальных клеток. Так, например, антибиотикорезистентные клетки просуществуют в чувствительной бактериальной популяции, а не образуются в результате воздействия на нее соответствующего антибиотика. Последний только селекционирует резистентные особи, создавая условия для их размножения. Индуцирующие мутации возникают в результате обработки микроорганизмов мутагенными агентами (ультрафиолетовые и рентгеновые лучи, быстрые нейтроны и протоны, действие температуры, кислоты, щелочи, красители, соли металлов и др.)
Разные мутагены отличаются по активности и механизму действия. Одни вызывают изменения числа и структуры хромосом, другие изменяют последовательность азотистых оснований в молекуле ДНК. Мутагены используются в селекции сельскохозяйственных растений и микроорганизмам с целью получения продуктивных и полезных форм.
Микроорганизмам, как и высшим организмам, свойственны генетические рекомбинации. Рекомбинация — это перегруппировка генетического материала (ДНК) родительских генетических структур (хромосом, плазмид и др.), приводящая к появлению новых сочетаний генов у потомства. Основной механизм рекомбинации — кроссинговер- перекрест хромосом, при котором происходит разрыв участков двух генетических структур, их обмен и воссоединение. У микроорганизмов рекомбинация осуществляется в результате обмена участками двух молекул ДНК, либо их фрагментом. Генетические рекомбинации возникают в результате комплексных процессов, они могут захватывать большие участки хромосомы или происходить в пределах отдельных генов. Существуют специальные гены, детерминирующие рекомбинационную способность клеток — реципиентов.
Передача генетического материала от одних бактерий к другим осуществляется путем трансформации, трансдукции и конъюгации.
Трансформация бактерий — передача наследственных свойств от одних бактерий к другим при помощи экстрагированной ДНК, без прямого контакта клеток донора и реципиента и без участия бактериофага. Явление трансформации установлено у многих бактерий (пневмококки, анаэробы и др.). История открытия феномена трансформации связана с исследованием Ф. Гриффитса (1928 г.), который описал превращение бескапсульного авирулентного пневмококка в капсульный вирулентный вариант. В процессе трансформации различают пять стадий:
. Адсорбция трансформирующей ДНК на поверхности микробной клетки;
. Проникновение ДНК в клетку реципиента;
. Спаривание внедрившейся ДНК с хромосомными структурами клетки; 4. Включение участка ДНК клетки — донора хромосомной структурой реципиента.
. Дальнейшее изменение нуклеоида в ходе последующих делений. Эффективность спаривания трансформирующих ДНК с гомологичным участком хромосомы реципиента зависит от гомологии ДНК донора и реципиента. Чем больше гомология, тем эффективнее спаривание, что определяет конечный результат трансформации, т.е. количество формирующихся рекомбинантов. Трансформировать могут признаки: устойчивость и чувствительность к антибиотикам, способность к синтезу ферментов и т.д. Как правило, при трансформации изменяется один какой-либо признак.
Заключение
Микроорганизмы служат излюбленными объектами для решения общих вопросов генетики, биохимии, биофизики, космической биологии и др. Культуры бактерий применяются для количественного определения аминокислот, витаминов, антибиотиков. Плодородие почв в значительной мере связано с жизнедеятельностью бактерии, минерализующих растительные и животные остатки с образованием соединений, усваиваемых с.-х. растениями.
Вместе с тем, синтезируя живое вещество клеток, бактерии накапливают большие количества органических соединений в почве. В верхних слоях окультуренной почвы на площади в 1 га содержится несколько тонн бактериальных клеток. Живущие в почве азотфиксирующие бактерии обогащают почву азотом. Исключительно велика роль клубеньковых бактерий, фиксирующих газообразный азот. Заражение семян бобовых растений нитрагином — препаратом, содержащим клетки клубеньковых бактерий, повышает урожай растений и накопление азота в почве. С помощью бактерий, сбраживающих пектиновые вещества, осуществляют мочку льна, конопли, кенафа и других лубяных культур. Разные виды бактерии применяют при получении из молока кисломолочных продуктов, масла и сыра.
Список используемой литературы
1.Гринн Н., Стаут У., Тейлор Д. Биология. М., 1990
.Воробьев А.А., Быков А.С. Микробиология и иммунология. М., 1999
.Радчук Н.А., Дунаев Г.В. Ветеринарная микробиология и иммунология. М., 1991
.Новиков Д.К., Генералов ИИ. Медицинская микробиология. Витебск, 2002
.Общая микробиология и иммунология. Солонеко А.А., Гласкович А.А. Минск, 2002
.Галактионов ВТ. Иммунология. М., 2000
. Гольдфарб Д. М., Введение в генетику бактерий, М., 1966;
Источник: www.BiblioFond.ru
Похожие главы из других работ:
2. Общая характеристика
Лилейные — это многолетние травянистые луковичные или корневищные растения, редко лианы и деревья. Все представители данного семейства являются геофитами, а также частично и эфемероидами. Строение луковиц…
2. Общая характеристика
Луковые — многолетние травы с луковицами, клубнелуковицами или иногда корневищами (триба агапантовых — Agapantheae). Корни обычно тонкие, нитевидные, но бывают и утолщенными…
1. Общая характеристика
Витамины (от лат. YITA — жизнь) — группа органических соединений разнообразной химической природы…
1. Общая характеристика
фермент витамин гормон Гормоны — специфические вещества, которые вырабатываются в организме и регулируют его развитие и функцианирование. В переводе с греческого — гормоны- означают двигаю, возбуждаю…
2.1 Общая характеристика РЧ — воздействий
При обсуждении возможных неблагоприятных эффектов для здоровья при воздействии на человека радиочастотных (РЧ) полей важно не путать РЧ-поля с ионизирующим излучением — таким как, рентгеновские лучи…
1.1 Общая характеристика мейобентоса
Термин «мейобентос» был введен в науку М. Мэр в 1942 г. для обозначения совокупности микроскопических многоклеточных, выпадающих из внимания исследователей морского бентоса при количественных учетах донной фауны…
1. Общая характеристика австралопитеков
История изучения австралопитеков ведет свое начало с 1924 года, с находки черепа детеныша гоминоида 3—5 лет в Юго-Восточном Трансваале (ныне ЮАР) близ Таунга…
Дерма: общая характеристика
Дерма — это основная часть кожи, придающая ей прочность, эластичность и способность выдерживать значительное давление и растяжение. Дерма состоит из соединительной ткани. В ней выделяются два основных слоя: сосочковый (stratum papillare)…
1.Общая характеристика
Отряд стрекозы ( ODONATA ) многообразен и интересен не только для науки, но и для любого человека. Отряд ODONATA (стрекозы) насчитывает, в мировой фауне, по различным источникам от 3600 ( 7 ) до 4500 (10 ) видов…
Общая характеристика
Отряд КРОКОДИЛЫ (Crocodylia) Все крокодилы и аллигаторы вместе с их близкими родственниками кайманами и гавиалами принадлежат к отряду Crocodylia. Их безошибочно можно узнать по ящерицеобразной форме тела…
5. Общая характеристика
Яичные породы У кур яичных пород наиболее распространён листовидный гребень, который за 2-3-м зубцом спадает набок. Куры яичных пород весят 1,8-2,2 кг, петухи — 2,7-3,0 кг. Цыплята после вылупления весят 30-35 г. Птица яичных пород более скороспела….
1. Общая характеристика сорта
Гвоздика ремонтантная крупноцветковая (Dianthus caryohyllus L. var. semperflorens) относится к семейству гвоздичные (Caryohyllacea Juss.) . Это сложный гибрид, полученный в результате многочисленных скрещиваний разных видов и форм. Широко выращивают гвоздику группы Сим…
Общая характеристика
Самая древняя и многочисленная группа хордовых животных. Около 22 тыс. видов, заселивших моря, океаны и пресные воды…
1.2 Общая характеристика чешуекрылых
Бабочки являются одними из самых красивых насекомых, отряд Lepidoptera включает в себя более 140 (по некоторым данным 150) тысяч видов. Тем не менее, среди других насекомых это довольно «молодая» группа…
Общая характеристика
Семейство КОШАЧЬИ (Felidae) Кошачьи — наиболее специализированные из всех хищных, всецело приспособленные к добыванию животной пищи преимущественно путем скрадывания, подкарауливания, реже — преследования и к питанию мясом своих жертв…
Источник: bio.bobrodobro.ru
| |
В целом, эукариоты очень разнообразны. Большая часть одноклеточных эукариот адекватно описывается таксонами уровня Царства. Есть много одноклеточных водорослей , Хризофитовыые, они идут как класс. Но эти водоросли, хролопласт – пример вторичного эндосимбиоза, этот примитивный эукариотический организм поглотил другой одноклеточный эукариотический организм. Сначала был эукариотический эндосимбиоз одноклеточного эукариотического нефотосинтезирующего организима с одноклеточным эукариотическим фотосинтезирующим организимом, с течение временишлаинтенсивная редукция генома эндосимбионта, часть генов мигрировала в геном хозяина, но по крайней мере в некоторых видах сохранилось даже редуцированное ядро эндосимбионта, так называемый, нуклеоморф, часть была нуклеоморфов секвенирована. Оказалось, что это стандартный эукариотический геном, похожий на эукариотический, похожий на прокариотический тем, что лишней балластной ДНК нет, гены перекрываются, около 1.5 млн. Существует два нуклеоморфа, различные эндосимбиозы. Плазмодий несёт интересную органеллу – апикопласт, тип Апикомплекса. Апикопласт отвечает за метаболизм различных соединений. Хролопласт Криптофитовых и апикопласт ограничены четырьми мембранами. Одна внешняя клеточная мембрана, вторая мембраны эндосимбиоза и две мембраны хлоропласта. В этом случае протоплазмы эндосимбионта практически не остаётся, даже нуклеоморф где-то сбоку лежит. В апикопласте ядра не сохранилось, однако наличие четырёх мембран подчеркивает происхождение. Если детально исследовать как работают молекулярно-генетические механизмы таких интересных одноклеточных эукариот, тоже много чудес получится. Есть различные водоросли, у которых и нормального митоза нет, например Dinoflagellata, у инфузорий интересно идёт деление.
Дрожжи – удобный модельный объект, это редуцированные аскомицеты. Два наиболее изученных вида дрожжей – Schizosacharomyces pombe и Sacharomyces cerevisiae, оба одноклеточные, имеют похожие геномы, между ними полмиллиарда до миллиарда по различным оценкам лет эволюции. Понятно, что был какой-то общий аскомицетный предок, от которого эти виды произошли. Пример филогенетического древа, эти грибы родственны другим грибам, другие эукариоты довольно обособлены. Основный характеристики геномов на первый взгляд схожи. У Sacharomyces cerevisiae 16 хромосом, у Schizosacharomyces pombe их всего 3. Число генов также существенно различается, первые оценки были существенно больше для обоих видов однако с каждым годом число кодирующих генов уменьшается. Около 5500 для Schizosacharomyces pombe, для Sacharomyces cerevisiae около 5000 генов. Существенные отличия начинаются, если посмотреть, сколько генов с интронами, у Schizosacharomyces pombe стандарный эукариотический геном, значительная часть генов (43%) с интронами, в случае, если интроны есть, он может быть один, может быть много. В случае Sacharomyces cerevisiae по различным оценкам от 3 до 5 % генов с интронами, во всех случаях кодирующая последовательность никогда не прерывается, когда интрон есть он в самом начале 5′ некодирующей области располагается. В целом плотность генов от 1 гена на 2,5 тысячи нуклеотидов. Если посмотреть, какой процент генома кодирует белки, то это порядок 60-70 %. Однако помимо генов, кодирующих белки, есть гены тРНК, есть гены рРНК, могут быть локализованы в виде тандемных повторов, могут быть разбросанными по геному, быть локализованными в центромерных и теломерных областях, наконец есть гены малых ядерных РНК, какое-то количество транспозонов. У пивных дрожжей меньше, у хлебных больше, есть также какое-то количество длинных концевых повторов, эта часть транспозонов ретровирусные, в дрожжах очень эффективно идёт гомологичная рекомбинация, поскольку ретротранспозоны ограничены двумя прямыми повторами, рекомбинация между ними выбрасывает центральную часть ретротранспозона, остаётся одиночный повтор, у дрожжей они называются дельта-последовательности. Существенное различие между этими дрожжами состоит в строении их теломерных и центромерных областей. У Sacharomyces cerevisiae достаточно просто устроенная центромера, менее сотни н.п., у пивных дрожжей строение центромеры близко к строению стандартной центромеры эукариот — это несколько десятков тысяч н.п. , в которых есть различные наборы различных повторов, часть из них так и называется – центромерные повторы, которые больше нигде не встречаются, скорее всего именно с этими повторами взаимодействуют белки, отвечающие за формирование веретена деления, присутствуют гены тРНК (показаны на слайде значками) и во многих случаях транспозоны присутствуют, у дрожжей это не характерно, так как их мало в геноме, у других эукариот они присутствуют.
Пару слов о геномном дрожжевом проекте. Два дрожжевых генома совершенно в разное время секвенировали, Sacharomyces cerevisiae были первым секвенированным эукариотическим организмом, а Schizosacharomyces pombe секвенировали в районе 2000 года (Пол Нёрс , Нобелевская премия), интерес к сиквенсу пивных дрожжей связан с тем, что именно на них исследовалось клеточное деление, Предстояло выяснить, какие гены деления являются общими, выделить ключевые гены деления, характерные для большинства эукариот. Однако не нашлось генов, кроме тех, которые были выявлены классическими генетическими методами. Первый дрожжевой геном сделан был ещё классическим подходом — составление сначала физических карт всех 16 хромосом, которые показали наличие большого числа дубликаций в геноме, хромосомы были розданы по лабораториям, по всему миру занимались секвенированием этого генома, однако тогда ещё не во всех лабораториях стояли автоматические флюорисцентные секвенаторы, секвенирование делалось вручную на ручных секвенаторах, выходило очень много диссертаций с сиквенсом и характеристикой отдельных хромосом. В итоговой статье получилось 600 авторов. В ходе секвенирования шёл активный обмен информацией между ключевыми группами, сформировалось Дрожжевое сообщество. Смысл этого проекта был двояким – получить сам сиквенс и сформировать коллектив, который дальше бы занимался дальнейшими исследованиями, выяснением функций генов.
Schizosacharomyces pombe имеют стандартные теломеры и теломеразу, Sacharomyces cerevisiae такового фермента, как теломераза не имеют. Считается, что один из Ty-транспозонов (Ty-5) взял на себя эту функцию, то есть обратная транскриптаза отвечает за репликацию концов 16 хромосом. Была надежда, что процессы старения на дрожжах можно будет активно изучать, однако их можно изучать лишь на пивных дрожжах лишь с большой натяжкой, на Sacharomyces cerevisiae это невозможно, у них просто нет фермента теломеразы.
Лекция №12
Этот слайд (См. слайд 6.5) показывает, какие интроны где и как располагаются у дрожжей. На данном слайде приведен процент генов, полностью лишенных интронов, с одним интроном, с двумя интронами, с тремя интронами и т.д. На слайде так же показано, на каком расстноянии от начала гена располагается интрон. На это можно не обращать внимание, но видно, что в целом интроны сдвинуты ближе к началу гена. Примерно половина генов не имеет интронов. Чем больше интронов, тем их фракция в геноме меньше. Интронов в геноме дрожжей относительно немного. Все другие эукариоты имеют значительно большее число интронов.
См.слайд 6.7
На момент окончания геномного проекта где-то только половине генов были приписаны определенные функции. Сейчас приписаны функции почти всем генам (до 100%). Но реально экспериментально показаны примерно до 70%. Оставшиеся 30% — это гипотетические функции, которые при проверке обычно оказываются иными. Можно посмотреть, как распределяются по различным функциональным категориям белковые продукты этих генов. Чуть больше ¼ занимают базовые функции, то есть, то без чего эукариотическая клетка не может существовать (гены, которые отвечают за гомеостаз и прочие функциональные категории). На этом слайде отдельно выделен рост клетки, деление, синтез ДНК, транскрипция, синтез белка и транспорт белка. В данном случае получается, что метаболические функции несущественны для жизнедеятельности. Если инактивировать вот эти гены, то получается несколько другая ситуация. Только 20% мутаций дают какой-то видимый фенотип, а инактивация 80% генов дрожжей не ухудшает рост дрожжей в культуре и не приводит к видимым изменениям. Фактически из всех 5000 генов для нормального существования в лабораторных условиях требуется только 1 тысяча генов в геноме эукариот. Сколько реально нужно генов эукариотическому организму для поддержания своего существования?
Геномы многоклеточных эукариотических организмов
Первым из беспозвоночных был идентифицирован геном нематоды Caenorhabditis elegans. Существует огромное разнообразие нематод. Это один из наиболее широко представленных в окружающей среде типов животных. В каждом кубическом сантиметре почвы может содержатся до нескольких миллионов особей. Нематоды могут обитать в почве, в воде, на или в различных организмах. Среди нематод есть много паразитов. Нематоды встречаются и в ледниках Антрактиды (в анабиозе). План организации тела нематод является достаточно удачным для многоклеточных животных. Но такой план организации тела специфичен и стоит особняком от организации тела других многоклеточных животных. Но самой главной отличительной особенностью нематод является то, что план строения тела очень жестко детерминирован и программа развития организма прописана с точностью до одной клетки. Лет 30 назад Сидни Бренер сделал нематоду модельным организмом, до этого нематод практически никто не изучал. Как только выяснилось, что у них очень жесткая программа развития, Бренер решил, что нужно отрабатывать молекулярно биологические методики на нематодах и секвенировать геном этого организма. Благодаря Бренеру уже к концу 80-х годов было полностью известно, как развивается этот организм, была построена карта развития нематоды.
См.слайд 6.10
На этом слайде показана генеалогия каждой клетки в организме. Организм нематоды состоит приблизительно из 936 клеток. Точно известно, из какой клетки какая образуется, как формируется каждый орган и сколько в нем клеток. Особый интерес этот организм вызывал в связи с тем, что точно известны особенности организации нервной системы. Нервная система нематоды состоит из 300 нейронов. Досконально известно, как формируется и развивается нервная система нематоды. Ученые пытались определить, каким образом работает эта нервная сеть. Такое относительно небольшое число нейронов можно изучать как функционирует самый примитивный биологический компьютер. Такие исследования интенсивно ведутся, но пока результаты этих исследований еще не дошли до относительно популярной литературы. Они слишком специфические, математизированные и компьютиризованные, чтобы в них можно было нормально разобраться. Этот организм позволяет на молекулярно биологическом уровне работать с конкретными «вещами», потому что четко известно, что каждая клетка в организме делает. У других организмов (человек) каждый орган состоит из неопределенного количества клеток (+- 30% клеток). Кроме того нематоды имеют очень компактный геном (всего 100 млн нуклеотидных пар).
См.слайд 6.11
На сегодняшний день геном нематод является минимальным геномом многоклеточного организма, который секвенирован. Нематода была первым отсеквенированным многоклеточным организмом. При секвинировании генома нематоды в основном использовалась космидная библиотека, потому что на момент старта геномного проекта еще не было разработано дрожжевых искусственных хромосом. К концу геномного проекта уже появляются дрожжевые искусственные хромосомы, но все-таки основная масса сиквенса, как показывает данная схема, была сделана на космидных клонах. ЯКи были добавлены потом, они помогли состыковать космиды между собой. Основная масса секвинирования осуществлялась с помощью космид. Размеры ЯК- клонов очень небольшие. Они представляют собой не мегабазные вставки, а вставки размером приблизительно пару сотен нуклеотидных пар. Стандартный размер космиды – 45000 пар нуклеотидов. Такие клоны с небольшим размером вставки оказываются относительно стабильными.
См. слайд 6.12
Геном нематоды Caenorhabditis elegans был состыкован из 2,5 тысяч космид, 113 фазмид и 257 ЯК-клонов. С теми последовательностями, которые никак не удалось состыковать, проводилась модифицированная ПЦР. Модифицированная ПЦР позволяет «отПЦРить» несколько десятков нуклеотидных пар. В результате еще в 1998 году был получен геном первого эукариотического организма, качество которого для эукариот остается до сих пор непревзойденным. Во-первых отсеквинирован практически весь геном, в отличие даже от человеческого генома. В сиквенсе генома нематоды Caenorhabditis elegans содержится меньше всего «дырок». Это стало возможным благодаря тому, что у нематод относительно мало гетерохроматина (практически нет гетерохроматина) и геном нематод маленький. Удалось «прочитать» значительную часть гетерохроматина нематод. Осталось менее 1% несеквенировано. Начиная с первого одноклеточного генома, процент недосеквенирования только рос. На момент публикации генома человека около 10% генома оставалось недосеквенированным. Сейчас остается недосеквенированным более 1% генома.
Около четверти генома Caenorhabditis elegans являются кодирующей частью генома. В данном геноме относительно немного геномных повторов.
См. слайд 6.13
Геном нематоды Caenorhabditis elegans включает приблизительно 19000 генов, кодирующих белок. В этой таблице приведены некоторые данные о геноме нематод на момент публикации генома (т. е. 10 лет назад). К сожалению, нематоды стоят особняком и гомология с генами других организмов совсем невысокая. На момент завершения геномного проекта были понятны функции только 40% генов нематоды Caenorhabditis elegans (т. е. 40% генов нематоды более или менее совпадали с функционально охарактеризованными генами других организмов). По современным данным 24% белков нематоды не имеют функциональной гомологии. 34% генов нематоды Caenorhabditis elegans имеют гомологию с генами нематод, секвенированных позднее. Однако такая гомология не способствует пониманию функциональной роли данных генов (белков) Caenorhabditis elegans. В результате функции почти 60% (24% + 34%) генов Caenorhabditis elegans остаются неизвестными. 34% генов Caenorhabditis elegans имеют гомологию с другими представителями класса нематод, но такая гомология никак не помогает пониманию функций соответствующих генов. В настоящий момент только у 40% всех генов Caenorhabditis elegans можнопредполагать какую-либо смысловую нагрузку. Геном нематоды характерен тем, что достаточно много (более 1000) генов не кодируют белок. Эти гены не являются «мусором», а необходимы нематодам. Большинство данных генов кодируют транспортную РНК (почти 900 генов). Минимум генов транспортных РНК, необходимых организму, — 43 или 44 тРНК в зависимости от модификации генетического кода и с учетом нестрого взаимодействия. Есть варианты, когда генетический код немного модифицируется и количество генов тРНК снижается. Любой организм, даже прокариотический, имеет избыток генов тРНК. У эукариот такая избыточность может быть значительной. Эта высокая избыточность генов тРНК у эукариот способствует тому, что оптимальный спектр используемых кодонов очень существенно варьирует у различных эукариот. Чем больше транспортных РНК, соответствующих определенному кодону, тем эффективнее идет трансляции соответствующих кодонов. Если какому-то кодону соответствует всего лишь одна тРНК, то трансляции будет идти очень неэффективно. При попытке экспрессии чужеродного гена в соответствующем организме необходимо учитывать оптимальные кодоны, имеющиеся в этом организме. Но в некоторых случаях не надо обращать на это внимание, потому что трансляция все равно будет эффективной. Следует учитывать, из какого организма взят ген и в какой организм он вводится. Если стандартный ген E. coli ввести в растение, то оптимизация кодонного состава повысит экспрессию только в 1,5 раза и поэтому в данном случае оптимизация кодонного состава не имеет смысла. Если стандартный ген E. coli ввести в животное, то он будет очень плохо экспрессироваться.
В геноме Caenorhabditis elegans от генов тРНК берут свое начало почти 200 псевдогенов (т. е. уже инактивированных генов тРНК). В сумме семейство генов тРНК содержит больше тысячи генов, это семейство генов составляет значительную часть генома Caenorhabditis elegans. Следующий класс не кодирующих белок генов – это малые ядерные РНК. Малые ядерные РНК входят в состав сплайсосомы. Малые ядерные РНК содержатся в U-частицах, которые вместе формируют сплайсосому. Сплайсосома – это крупный рибонуклеопротеиновый комплекс, который занимает существенный процент массы клетки. Для того, чтобы получить нормальный уровень сплайсосомных частиц и чтобы сплайсинг происходил эффективно, обычно клетка имеет множественные дупликации этих генов (от 5 – 20 генов). SRP (signal recognition particle) — это частица, которая обеспечивает направление секретируемого пептида в полость эндоплазматического ретикулама. В состав SRP входят 7S РНК (больше 300 нуклеотидных пар) и 6 белков. Частиц SRP в клетке достаточно много, поэтому и соответствующие гены дублируются. Еще два важных класса РНК – это SL1 и SL2 РНК. S1 и S2 РНК специфичны только для нематод. У каждого организма гены рРНК несколько по-разному организованы. Гены рРНК чаще всего располагаются целым кластером на одной хромосоме или могут быть диспергированы по геному несколькими кластерами (отдельными генами). Но обычно это один крупный кластер на одной хромосоме. У Caenorhabditis elegans кластер генов рРНК располагается на первой хромосоме.
См. слайд 6.14
Кодирующие белок гены Caenorhabditis elegans. Наиболее часто встречающиеся белковые домены Caenorhabditis elegans являются трансмембранными рецепторами с семи трансмембранными доменами, которые представлены несколькими семействами (3 семейства). Такие рецепторы имеют больше тысячи доменов и кодируются еще большим числом генов. Около двух тысяч генов Caenorhabditis elegans кодируют рецепторы. Значительное количество кодирующих рецепторы генов является принципиальным отличием от одноклеточных эукариот. У дрожжей тоже есть рецепторы, но их совсем мало (пару сотен на 1 клетку). Такая ситуация очень четко показывает наиболее принципиальное отличие в организации многоклеточного организма на молекулярном уровне от таковой у одноклеточных. Необходима четкая межклеточная коммуникация и координация согласованного действия всех клеток многоклеточного организма, что требует наличия нормальных систем коммуникации. Для функционирования таких систем коммуникации нужны рецепторы. В данной таблице нет генов, которые отвечают за синтез соответствующих лигандов. Для каждого рецептора существует свой лиганд. В геноме многоклеточного организма должны содержатся кодирующие лиганд гены, в количестве (как минимум) равном количеству генов рецепторов. В геноме многоклеточного организма должны присутствовать контролирующие синтез рецептора и лиганда гены и гены, обеспечивающие передачу сигнала от рецептора к ядру. В итоге получается, что, как минимум четверть всего протеома клетки многоклеточного организма отвечает исключительно за межклеточную коммуникацию.
SL1 и SL2 — это сплайслидерная РНК.
См. слайд 6.15
Практически все транскрипты у трипаносомы не кепируются, потому что не имеют сайтов к которым кеп присоединяется. У этого организма система кепирования функционирует совершенно по-другому. Кепируется только 1 единственная молекула РНК – SL1 РНК, которая распознается системой кепирования. Трансляция у эукариот осуществляется только в том случае, если матричная РНК кепирована. Кеп распознается эукариотическими факторами инициации трансляции, на кепе происходит сборка рибосомы. Далее происходит сканирование до первого кодона и инициируется трансляция. Кепа нет – нет и трансляции (отличие от прокариот)! Поскольку стандартный транскрипт у трипанасомы не имеет кепа на своем конце (т. е. он не модифицируется), то единственный способ обеспечить трансляцию такого транскрипта это пришить кепированную последовательность. Пришивание кепированной последовательности к транскрипту осуществляется в процессе транссплайсинга. Сплайсинг относится к реакциям трансэтерификации, которые осуществляются между двумя молекулами РНК. При трансэтерефикации происходит изомеризация фосфодиэфирной связи. Эта реакция в норме идет без затраты энергии. Но клетка тратит энергию для контролирования данной реакции.
См. слайд 6.16
Нематоды уникальные организмы (но сейчас показано, что есть и другие осуществляющие транссплайсинг организмы: трипанасомы, и т.д.), т. к. у них происходит транссплайсинг. На нематодах впервые было показано, что у эукариот может быть оперонная организация. Около 25% всех генов Caenorhabditis elegans «упакованы» в опероны, но опероны эукариот имеют проблему с инициацией трансляции внутренних рамок считывания. Даже если в первичный транскрипт будет сплайсирован, то только трансляция данной рамки будет происходить нормально. Инициация трансляции на выходе невозможна, т. к. нет механизма, который используется у прокариот, типа трансляционного сопряжения, либо внутренней последовательности Шайна-Дальгарно. Кроме того, в малой субъединице рибосомной РНК нет соответствующего комплементарного участка, т. е. 16S РНК имеет делецию 5 нуклеотидов, которые соответствуют прокариотической последовательности Шайна-Дальгарно. Каким-то образом должна быть обеспечена инициация трансляции на внутренних рамках считывания. Эта проблема решается достаточно просто. За счет транссплайсинга к внутренним рамкам точно также, как и у трипанасомы к любой рамки, пришивается кепированная лидерная РНК. Единственная модификация данного процесса у нематод заключается в том, что у нематод есть две лидерных РНК. SL1 РНК точно такая же как и у трипаносом, она пришивается к самому началу транскрипта. А SL2 РНК присоединяется к внутренним рамкам. Сейчас показано, что у многих беспозвоночных животных (даже у представителей класса хордовых) осуществляются процессы трансплайсинга, которые позволяют иметь оперонную организацию. У низших хордовых показано наличие транссплайсинга. Возможно, даже у человека есть системы транссплайсинга, но пока нет экспериментальных данных подтверждающих это. Скорее всего процессы транссплайсинга имеют более широкое распространение, чем мы себе представляем.
См. слайд 6.18
Чем еще интересен организм Caenorhabditis elegans в качестве экспериментального объекта? В организме Caenorhabditis elegans очень легко проходит РНК-интерференция. Существует множество методов индукции РНК-интерференции. Первоначально такую индукцию проводили за счет инъекции интерферирующей молекулы РНК в нематоду. Но такой способ трудно технически осуществить, т. к. организм нематоды мелкий. Совершенно случайно открыли, что можно замочить нематоду в растворе РНК. Можно также накормить нематоду бактериями, которые экспрессируют
интерферирующую РНК. Те методы, которые запустили развитие этих технологий, связаны с кормлением нематоды бактериальным газоном, клетки которого экспрессируют соответствующую интерферирующую РНК.
См. слайд 6.19
На момент выхода только этих геномов был сделан интересный анализ «на кого больше всего похожа нематода?». Тогда было отсеквенировано лишь небольшое число геномов (дрожжи, E. coli и т. д.). В результате получилось, что нематода больше всего похожа на Homo sapiens. В ходе данного анализа выбирались известные гены для данного организма, сравнивались в двух направлениях. Далее определяли какой процент генов (например, E. coli) имеет сходство с генами Caenorhabditis elegans (26%) и наоборот сколько генов Caenorhabditis elegans имеет сходство с E. coli (процент получается значительно меньше, т. к. генов у Caenorhabditis elegans больше, чем у E. coli и значительная часть генов эволюционно новая и не имеет сходства с E. coli). Подобная ситуация складывалась и при сравнении генома Caenorhabditis elegans с другими маленькими геномами. 51 % генов дрожжей Saccharomyces cerevisiae (пекарские дрожжи) имеет сходство с генами нематоды и только 26% генов нематоды имеют сходство с генами Saccharomyces cerevisiae. В эти 26% генов нематоды не попадают гены, кодирующие сигнальные системы, которые необходимы многоклеточному животному и гены, специфические только для нематоды. 74% генов человека имеют гомологию с генами Caenorhabditis elegans и 36% генов нематоды имеют сходство с генами человека. Это объясняется тем, что на момент проведения данного анализа еще не было сиквенса полного генома человека и речь шла только об изученных генах человека. К этим изученным генам человека относились жизненно важные гены, имеющие отношение к базовому гомеостазу, поэтому они были гомологичны генам Caenorhabditis elegans.
См. слайд 6.20
Первым из членистоногих был отсеквинирован геном дрозофилы. Дрозофила – очень удобный экспериментальный объект. На момент начала геномного проекта 2,5 тысяч генов уже было охарактеризовано, клонировано, секвенировано и нанесено на генетическую карту. Генетические карты дрозофил существуют примерно сто лет (1913 г). Среди двукрылых встречается большое количество хозяйственно важных организмов, на пример, малярийный комар. В Африке малярия до сих пор уносит миллионы жизней каждый год. На территории бывшего СССР удалось подавить малярийного комара за счет применения ДДТ и мелиорации (ликвидации экологических ниш). В Средней Азии малярийный комар все еще встречается, а на европейской территории проблема была решена. Раньше на Украине малярийный комар массово размножался в …. Днепра. После того, как повсему Днепру были построены водохранилища, малярийные комары исчезли вследствие ликвидации мест их размножения. Геном малярийного комара был расшифрован через несколько лет после генома дрозофилы. Исследования организма малярийного комара актуально, т. к. он переносит и другие заболевания.
См. слайд 6.21
Сиквенс генома дрозофилы был сделан под руководством Вентера.
См. слайд 6.23
Дрозофила оказалась удачным объектом для секвенирования и сточки зрения геномики, т. к. она имеет очень компактный геном (всего 180 млн нуклеотидных пар).
См. слайд 6.24
Геном дрозофилы является самым небольшим среди известных геномов членистоногих. Около трети генома дрозофилы существует в виде гетерохроматина (на слайде показаны черными блоками). В связи с тем, что гетерохроматиновые участки вообще не клонируются в бактериальных векторах (токсичны для них), Вентер не сиквенировал гетерохроматин дрозофилы. Т. к. в гетерохроматине содержится только небольшое количество генов, решение Вентера является достаточно обоснованным. После секвенирования генома дрозофилы никто и не пытается с начала клонировать и изучать гетерохроматиновые участки. В первую очередь секвенируется эухроматиновая часть генома, а только потом, по возможности, пытаются с концов «вгрызца» в гетерохроматин (не всегда удается). Чтобы иметь полное представления о всем геноме человека, сейчас делаются попытки секвенирования человеческого гетерохроматина, который составляет приблизительно 3% от нашего генома. Человеческий гетерохроматин менее компактен, чем гетерохроматин дрозофилы. У дрозофилы есть всего 2 больших аутосомы, половые хромосомы (Х и У хромосомы) и одна маленькая аутосома. Маленькая аутосома содержит большое количество гетерохроматина. У хромосома почти полностью состоит из гетерохроматина. Небольшое количество хромосом у дрозофилы также значительно облегчило осуществление данного геномного проекта. В геноме дрозофилы имеется больше повторов, чем у Caenorhabditis elegans и фракция кодирующего генома значительно меньше, что представляло проблему для расшифровки генома. Поэтому Вентер использовал модификацию шотган-подхода.
См. слайд 6.22
Основная часть сиквенса осуществлялась с использованием pUC со вставками приблизительно 2000 нуклеотидных пар и библиотеки такого же типа, но с более крупными вставками (10000 нуклеотидных пар) и векторами меньшей копийности, чем pUC. Копийность векторов с большими вставками следует снижать, чтобы избежать гибели значительной части клеток, содержащих такие векторы. Для секвенирования генома Caenorhabditis elegans также использовалась библиотека на основе БАКов (бактериальные искусственные хромосомы). БАКи стабильно «держат» вставки приблизительно 100000 – 150000 (иногда даже до 300000) нуклеотидных пар. Обычно в геномных проектах такие вставки составляют от 100 до 150 тысяч нуклеотидных пар. В данном случае средний размер вставки в БАКах составлял 120 -130 тысяч нуклеотидных пар. Библиотека на основе БАКов использовалась для «стыковки» фрагментов генома между собой, в пределах концов каждого БАКа. Поскольку известно, что два таких сиквенса принадлежат к концу одного фрагмента размером 130000 нуклеотидных пар, можно было «состыковать» различные контиги между собой. Таким образом, становилось понятно, какие фрагменты ДНК нужно прицельно секвенировать на стадии финиширования. Кроме того с БАКами проводилось STS-картирование. Вентер использовал уже имеющиеся физические карты и сделал STS-карту генома дрозофилы, используя БАК библиотеку в качестве реагента для картирования. Тогда STS-маркеры было достаточно легко секвенировать, потому что значительная часть генома дрозофилы на тот момент уже была секвенирована. В данном геномном проекте активно использовалась очень старая карта политенных хромосом, карта политенных хромосом дрозофилы, которая была сделана в 30-е года 20 века (позже она была насыщена). К моменту осуществления геномного проекта была произведена привязка некоторых БАКов за счет FISH гибридизации (флуоресцентная гибридизация in situ). Карта политенных хромосом была сделана с достаточно высоким двадцатикилобазным разрешением (очень хорошее разрешение) с привязкой ключевых локусов, что потом существенно упростило сборку всего генома. Основными ресурсами для физического картирования являются STS-карта, карта политенных хромосом, карта на основе БАК библиотеки (позволяет состыковать фрагменты). Окончательную стыковку и завершение физической карты удалось сделать при черновом секвенировании, на БАК библиотеке было сделано приблизительно полуторакратное перекрытие генома, что позволило закончить полную физическую карту генома дрозофилы.
См. слайд 6.25
Ciona intestinalis (оболочник)
Оболочники живут в теплых морях, в большом количестве обитают в Средиземном море. Первым этот организм достаточно подробно описал Аристотель. Потом интерес к этому организму угас. Только в 19 веке Ковалевский возродил интерес к оболочникам. Ковалевский показал, что оболочники относятся к хордовым животным. Ciona intestinalis легко культивируется и размножается в условиях лаборатории, является достаточно простым экспериментальным объектом. Но к 30-м годам 20 века после изучения этого организма учеными-физиологами интерес к Ciona intestinalis снова спал, т. к. не было никаких других методов анализа устройства этого организма. Вновь интерес к оболочникам возник в геномную эру (в 90-е годы 20 века). Ciona intestinalis – относительно удобный модельный организм, который относится к хордовым и на котором можно будет проследить взаимосвязь между беспозвоночными и позвоночными животными. Ученым удалось выиграть небольшое финансирование на расшифровку генома оболочника. В начале 21-го века был сделан сиквенс генома Ciona intestinalis, но его качество в связи с нехваткой финансирования оказалось невысоким. Фактически полученный сиквенс является черновиком генома, содержащим много «дырок», но этот сиквенс все таки дает представление об этом геноме и о присутствующих в нем генах. Данный сиквенс позволяет определить, на геном позвоночного или беспозвоночного животного похож геном Ciona intestinalis.

Оболочники относятся ко вторичноротым животным, к которым кроме позвоночных животных относятся еще иглокожие, полухордовые, головохордовые (ланцетник).
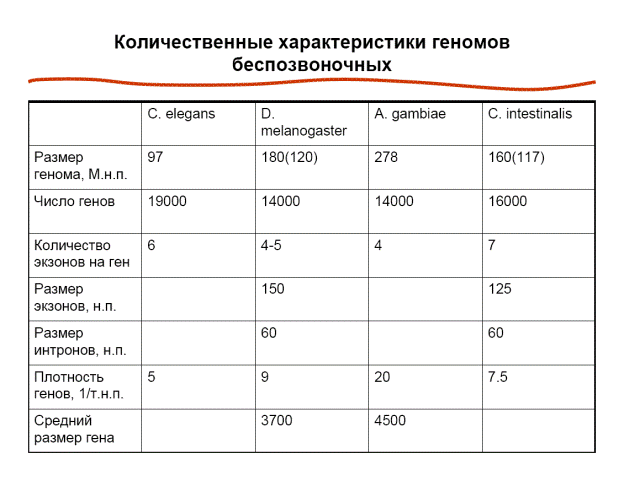
Сводная таблица, которая характеризует геномы беспозвоночных животных.
Геном может варьировать в достаточно широких пределах. Так получилось, что модельные организмы, которые были первыми секвенированы, имеют меньшие геномы. Из 180 млн нуклеотидных пар дрозофилы реальный геном составляют приблизительно 120 млн нуклеотидных пар. Среди комаров размер генома варьирует в широких пределах. Даже в роде Anopheles есть виды, у которых количество нуклеотидных пар в гегноме в 2 — 4 раза больше. Из 160 млн нуклеотидных пар в геноме Ciona intestinalis (очень приблизительная оценка), только 117 млн нуклеотидных пар входят в состав эухроматина. Из данной таблици можно сделать вывод, что стандартный размер генома беспозвоночного животного составляет приблизительно 100 млн или чуть больше нуклеотидных пар. 100 млн нуклеотидных пар – это минимальный размер генома, необходимый при такой организации для размещения в нем нужного количества генов (19000 для нематод, около 14000 для дрозофилы, 14000 для Anopheles, приблизительно 16000 для Ciona intestinalis). Количество кодирующих белок генов в большинстве геномов беспозвоночных вписывается в интервал от 14 до 19 тысяч. Некодирующие белок гены обычно рассматриваются отдельно. Количество некодирующих белок генов беспозвоночных животных приблизительно в 20 раз меньше, чем количество кодирующих белок генов. Количество некодирующих белок генов беспозвоночных животных также может несколько варьировать. Гены беспозвоночных, в отличие от дрожжей, в большинстве случаев представляют собой стандартные прерывистые гены. В среднем ген беспозвоночного животного содержит от 4 до 7 экзонов. Среди беспозвоночных уже не встречается такая ситуация, когда значительная часть генов лишена интронов. Интроны практически всегда присутствуют в генах беспозвоночных животных. Количество интронов в гене может варьировать. Размер экзонов беспозвоночного животного может существенно различаться. У нематод экзоны довольно крупные. Нормальный размер экзона у беспозвоночных и позвоночных животных принципиально не отличается. В целом нормальный размер экзона у всех эукариот, включая и человека, составляет 100 – 150 нуклеотидных пар. Беспозвоночные и позвоночные животные различаются по размеру интронов. У беспозвоночных интроны короче. Средний размер интрона беспозвоночного животного приблизительно равен приблизительно 60 нуклнеотидным парам. У человека размеры интрона составляют тысячи и десятки тысяч нуклеотидных пар. За счет компактности генома у беспозвоночных животных плотность генов высокая. У нематод максимальная плдотность генов, приблизительно 1 ген на 5000 нуклеотидных пар, у дрозофилы – чуть больше. Для Anopheles плотность генов меньше, т. к. в его геноме содержится большое количество минисателлитных повторов. У Ciona intestinalis — порядка 2 генов на 7,5 тысяч нуклеотидных пар (очень приблизительная цифра). Средний размер гена беспозвоночного животного, который определяется размером интронов и экзонов, количеством экзонов и интронов, составляет несколько тысяч нуклеотидных пар.
Бактерии
Borrelia burgdorferi
Borrelia burgdorferi – возбудитель лайм-боррелиоза, достаточно часто встречается в РБ. Первым симптомом этого заболевания является воспаление и покраснение места укуса клеща. В этом случае рекомендуется принять рифампицин и сделать соответствующие анализы. Боррелез лечится, но может иметь серьезные последствия для здоровья человека. В норме укус клеща не вызывает воспалительной реакции. Случаи заражения боррелиозом сейчас встречаются чаще, чем случаи заражения энцефалитом. Место укуса энцефалитного клеща никак не отличается от места укуса здорового клеща. Borrelia относится к спирохетам. К спирохетам также относятся возбудитель сифилиса (бледная трепонема – Treponema pallidum). Геномы Borrelia и Treponema pallidum имеют много общего. Основу геномов Borrelia и Treponema pallidum составляет 1 ли
Источник: megalektsii.ru