Наличие ядра у растений
 |
|
| Архитектура Астрономия Аудит Биология Ботаника Бухгалтерский учёт Войное дело Генетика География Геология Дизайн Искусство История Кино Кулинария Культура Литература Математика Медицина Металлургия Мифология Музыка Психология Религия Спорт Строительство Техника Транспорт Туризм Усадьба Физика Фотография Химия Экология Электричество Электроника Энергетика |
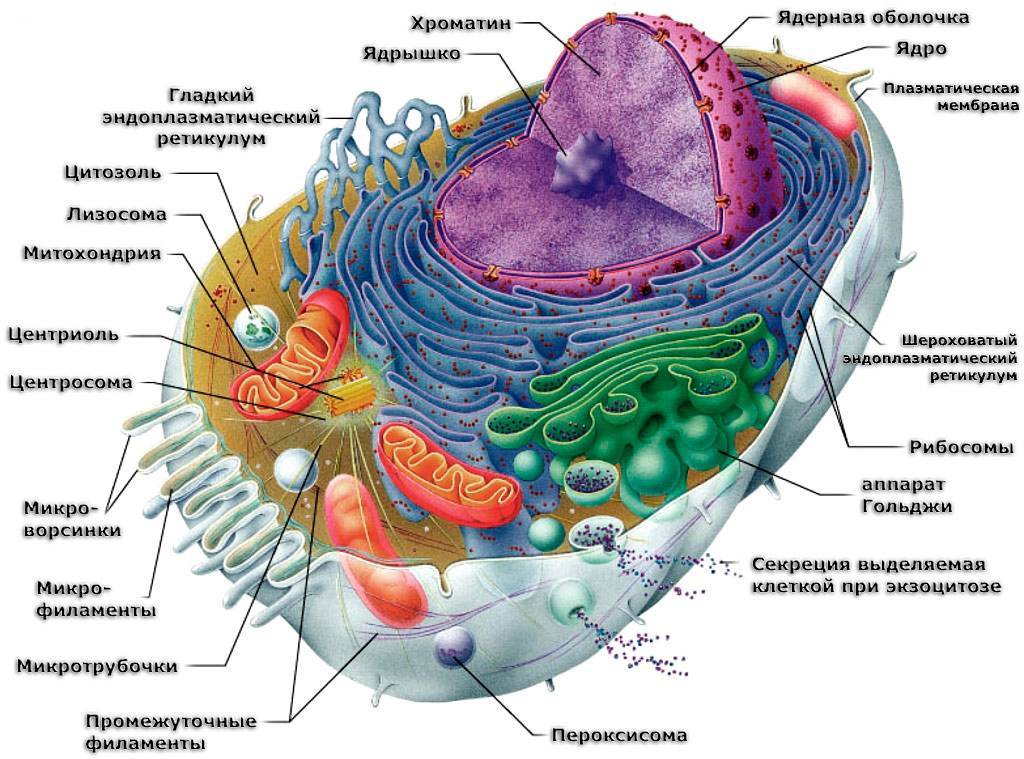
Ядро– обязательная часть эукариотической клетки. Это место хранения и воспроизведения наследственной информации. Ядро также служит центром управления обменом веществ и почти всех процессов, происходящих в клетке. Чаще всего в клетках имеется лишь одно ядро, редко — два или несколько. Форма его чаще всего шаровидная или эллипсоидальная. В молодых, особенно меристематических, клетках оно занимает центральное положение, но позднее обычно смещается к оболочке, оттесняемое растущей вакуолью. Снаружи ядро покрыто двойной мембраной – ядерной оболочкой, пронизанной порами, на краях которых наружная мембрана переходит во внутреннюю. Внутреннее содержимое ядра – кариоплазма с погруженными в нее хроматином и ядрышками, и рибосомами. В процессе клеточного деления хроматин все более уплотняется и в конце концов собирается в хромосомы. По химическому составу ядро отличается высоким содержанием ДНК. Основная масса ДНК сосредоточена в хроматине — особых нуклеопротеидных нитях, рассеянных по всему ядру. В ядре заметно одно или несколько ядрышек. Подобно хроматину, ядрышки не имеют мембраны и свободно лежат в кариоплазме, состоя в основном из белка. Они содержат РНК и имеют большую плотность, чем ядро. Основная функция ядрышек — синтез некоторых форм РНК и формирование предшественников рибосом. 7.Эндоплазматическая сеть: Эндоплазматический ретикулум (ЭПС) — система сообщающихся или отдельных трубчатых каналов и уплощенных цистерн, расположенных по всей цитоплазме клетки. Они отграничены мембранами (мембранными органеллами). Иногда цистерны имеют расширения в виде пузырьков. Каналы ЭПС могут соединяться с поверхностной или ядерной мембранами, контактировать с комплексом Гольджи. В данной системе можно выделить гладкую и шероховатую (гранулярную) ЭПС. Шероховатая ЭПС На каналах шероховатой ЭПС в виде полисом расположены рибосомы. Здесь протекает синтез белков, преимущественно продуцируемых клеткой на экспорт (удаление из клетки), например, секретов железистых клеток. Здесь же происходят образование липидов и белков цитоплазматической мембраны и их сборка. Плотно упакованные цистерны и каналы гранулярной ЭПС образуют слоистую структуру, где наиболее активно протекает синтез белка. Это место называется эргастоплазмой. Гладкая ЭПС На мембранах гладкой ЭПС рибосом нет. Здесь протекает в основном синтез жиров и подобных им веществ (например, стероидных гормонов), а также углеводов. По каналам гладкой ЭПС также происходит перемещение готового материала к месту его упаковки в гранулы (в зону комплекса Гольджи). В печеночных клетках гладкая ЭПС принимает участие в разрушении и обезвреживании ряда токсичных и лекарственных веществ (например, барбитуратов). В поперечно-полосатой мускулатуре канальцы и цистерны гладкой ЭПС депонируют ионы кальция.
8.Рибосома:
Рибосомы присутствуют в клетках как эукариот, так и прокариот, поскольку выполняют важную функцию в биосинтезе белков. В каждой клетке имеются десятки, сотни тысяч (до нескольких миллионов) этих мелких округлых органоидов. Это округлая рибонуклеопротеиновая частица. Диаметр ее составляет 20—30 нм. Состоит рибосома из большой и малой субъединиц, которые объединяются в присутствии нити м-РНК (матричной, или информационной, РНК). Комплекс из группы рибосом, объединенных одной молекулой м-РНК наподобие нитки бус, называется полисомой. Эти структуры либо свободно расположены в цитоплазме, либо прикреплены к мембранам гранулярной ЭПС (в обоих случаях на них активно протекает синтез белка). Полисомы гранулярной ЭПС образуют белки, выводимые из клетки и используемые для нужд всего организма (например, пищеварительные ферменты, белки женского грудного молока). Кроме этого, рибосомы присутствуют на внутренней поверхности мембран митохондрий, где также принимают активное участие в синтезе белковых молекул. Синтез рибосом у эукариот происходит в специальной внутриядерной структуре — ядрышке. 9.Митохондрии: Митохондрии (от гр. mitos — «нить», chondrion — «зернышко, крупинка») — это постоянные мембранные органеллы округлой или палочковидной (нередко ветвящейся) формы. Толщин — 0,5 мкм, длина — 5—7 мкм. Количество митохондрий в большинстве животных клеток — 150—1500; в женских яйцеклетках — до нескольких сотен тысяч, в сперматозоидах — одна спиральная митохонондрия, закрученная вокруг осевой части жгутика. Основные функции митохондрий: 1)играют роль энергетических станций клеткок. В иих протекают процессы окислительного фосфорилирования (ферментативного окисления различных веществ с последующим накоплением энергии в виде молекул аденозинтрифосфата —АТФ); 2)хранят наследственный материал в виде митохондриальной ДНК. Митохондрии для своей работы нуждаются в белкаx, закодированных в генах ядерной ДНК, так как собственная митохондриальная ДНК может обеспечить митохондрии лишь несколькими белками. Побочные функции — участие в синтезе стероидных гормонов, некоторых аминокислот (например, глютаминовой). Строение митохондрий Митохондрия имеет две мембраны: наружную (гладкую) и внутреннюю (образующую выросты — листовидные (кристы) и трубчатые (тубулы)). Мембраны различаются по химическому составу, набору ферментов и функциям. У митохондрий внутренним содержимым является матрике — коллоидное вещество, в котором с помощью электронного микроскопа были обнаружены зерна диаметром 20—30 нм (они накапливают ионы кальция и магния,запасы питательных веществ,например,гликогена). В матриксе размещается аппарат биосинтеза белка органеллы: 2-6 копий кольцевой ДНК, лишенной гистоновых белков (как у прокариот), рибосомы, набор т-РНК, ферменты редупликации, транскрипции, трансляции наследственной информации. Этот аппарат в целом очень похож на таковой у прокариот (по количеству, структуре и размерам рибосом, организации собственного наследственного аппарата и др.), что служит подтверждением симбиотической концепции происхождения эукариотической клетки. В осуществлении энергетической функции митохондрий активно участвуют как матрикс, так и поверхность внутренней мембраны, на которой расположена цепь переноса электронов (цитохромы) и АТФ-синтаза, катализирующая сопряженное с окислением фосфорилирование АДФ, что превращает его в АТФ. Митохондрии размножаются путем перешнуровки, поэтому при делении клеток они более или менее равномерно распределяются между дочерними клетками. Так, между митохондриями клеток последовательных генераций осуществляется преемственность. Таким образом, митохондриям свойственна относительная автономность внутри клетки (в отличие от других органоидов). Они возникают при делении материнских митохондрий, обладают собственной ДНК, которая отличается от ядерной системой синтеза белка и аккумулирования энергии. 10.Аппарат Гольджи:названпоимениитальянскогоученого К. Гольджи, впервые описавшего его в животных клетках. В клетках растений аппарат Гольджи состоит из отдельныхдиктиосом, или телец Гольджи и пузырьков Гольджи. Каждая диктиосома представляет собой стопку из 5-7 и более уплощенных округлых цистерн диаметром около 1 мкм, ограниченных мембраной. По краям диктиосомы часто переходят в систему тонких ветвящихся трубок. Число диктиосом в клетке сильно колеблется (от 10-50 до нескольких сотен) в зависимости от типа клетки и фазы ее развития. Пузырьки Гольджи различного диаметра отчленяются от краев диктиосомных цистерн или краев трубок и направляются обычно в сторону плазмалеммы или вакуоли. Диктиосомы являются центрами синтеза, накопления и выделения полисахаридов, прежде всего пектиновых веществ и гемицеллюлоз матрикса клеточной стенки и слизей. Пузырьки Гольджи транспортируют полисахариды к плазмалемме. Особенно развит аппарат Гольджи в клетках, интенсивно секретирующих полисахариды. 11.Лизосомы:–органеллы, отграниченные от гиалоплазмы мембраной и содержащие гидролитические ферменты, способные разрушать органические соединения. Лизосомы растительных клеток представляют собой мелкие (0,5-2 мкм) цитоплазматические вакуоли и пузырьки – производные эндоплазматической сети или аппарата Гольджи. Основная функция лизосом — локальный автолиз – разрушение отдельных участков цитоплазмы собственной клетки, заканчивающееся образованием на ее месте цитоплазматической вакуоли. Локальный автолиз у растений имеет в первую очередь защитное значение: при временном недостатке питательных веществ клетка может сохранять жизнеспособность за счет переваривания части цитоплазмы. Другая функция лизосом – удаление изношенных или избыточных клеточных органелл, а также очищение полости клетки после отмирания ее протопласта, например при образовании водопроводящих элементов. Сферосомы или олеосомы— в цитологии, одномембранные органеллы растительных клеток, выполняющие функцию накопления липидов. Образуются гладкой эндоплазматической сетью. На основании обнаружения в сферосомах активности кислой фосфатазы и неспецифичных эстераз их отождествляют с лизосомами животных клеток, от которых они отличаются высоким содержанием липидов. 12.Пластиды:– органеллы, встречающиеся только в растительной клетке. три типа пластид : 1. хлоропласты – самые крупные, зеленые , имеющие форму двояковыпуклой линзы , выполняющие функцию фотосинтеза .2.Лейкопласты – бесцветные пластиды, округлой или овальной формы, выполняющие функции синтеза и накопления вторичного крахмала , белков и липидов . 3. Хромопласты – разнообразной формы; желтого, оранжевого, красного или бурого цвета , придающие рекламную окраску органам растений. Хлоропластыимеют наибольшее значение, в них протекает фотосинтез. Они содержат зеленый пигмент хлорофилл, придающий растениям зеленый цвет, и пигменты, относящиеся к группе каротиноидов. Каротиноиды имеют окраску от желтой и оранжевой до красной и коричневой, но обычно она маскируется хлорофиллом. Каротиноиды делят на каротины, имеющие оранжевую окраску, и ксантофиллы, имеющие желтую окраску. Это липофильные (жирорастворимые) пигменты, по химической структуре они относятся к терпеноидам. Хлоропласты растений имеют форму двояковыпуклой линзы и размеры 4-7 мкм, они хорошо видны в световой микроскоп. Число хлоропластов в фотосинтезирующих клетках может достигать 40-50. У водорослей роль фотосинтетического аппарата выполняют хроматофоры. Их форма разнообразна: чашевидная (хламидомонада), лентовидная (спирогира), пластинчатая (пиннулярия) и др. Хроматофоры значительно крупнее, число их в клетке – от 1 до 5. Хлоропласты имеют сложное строение. От гиалоплазмы они отграничены двумя мембранами – наружной и внутренней. Внутреннее содержимое называется строма. Внутренняя мембрана формирует внутри хлоропласта сложную, строго упорядоченную систему мембран, имеющих форму плоских пузырьков, называемых тилакоидами. Тилакоиды собраны в стопки — граны, напоминающие столбики монет. Граны связаны между собой тилакоидами стромы (межгранными тилакоидами), проходящими через них насквозь вдоль пластиды (рис. 2.5 ). Хлорофиллы и каротиноиды встроены в мембраны тилакоидов гран. В строме хлоропластов находятся пластоглобулы – сферические включения жирных масел, в которых растворены каротиноиды, а также рибосомы, сходные по величине с рибосомами прокариот и митохондрий, и нити ДНК. Часто в хлоропластах встречаются крахмальные зерна, это так называемый первичный, или ассимиляционный крахмал – временное хранилище продуктов фотосинтеза. Хлорофилл и хлоропласты образуются только на свету. Растения, выращенные в темноте, не имеют зеленой окраски и называются этиолированными. Вместо типичных хлоропластов в них образуются измененные пластиды, не имеющие развитой внутренней мембранной системы, — этиопласты. Основная функция хлоропластов – фотосинтез, образование органических веществ из неорганических за счет энергии света. Центральная роль в этом процессе принадлежит хлорофиллу. Он поглощает энергию света и направляет ее на осуществление реакций фотосинтеза. Эти реакции подразделяются на светозависимые и темновые (не требующие присутствия света). Светозависимые реакции состоят в преобразовании световой энергии в химическую и разложении (фотолизе) воды. Они приурочены к мембранам тилакоидов. Темновые реакции – восстановление углекислого газа воздуха водородом воды до углеводов (фиксация СО2) – протекают в строме хлоропластов. В хлоропластах, как и в митохондриях, происходит синтез АТФ. В этом случае источником энергии служит солнечный свет, поэтому его называют фотофосфорилированием. Хлоропласты участвуют также в синтезе аминокислот и жирных кислот, служат хранилищем временных запасов крахмала. Наличие ДНК и рибосом указывает, как и в случае митохондрий, на существование в хлоропластах своей собственной белоксинтезирующей системы. Действительно, большинство белков мембран тилакоидов синтезируется на рибосомах хлоропластов, тогда как основное число белков стромы и липиды мембран имеют внепластидное происхождение. Лейкопласты —мелкие бесцветные пластиды. Они встречаются в основном в клетках органов, скрытых от солнечного света, таких как корни, корневища, клубни, семена. Строение их в общих чертах сходно со строением хлоропластов: оболочка из двух мембран, строма, рибосомы, нити ДНК, пластоглобулы аналогичны таковым хлоропластов. Однако, в отличие от хлоропластов, у лейкопластов слабо развита внутренняя мембранная система. Лейкопласты – это органеллы, связанные с синтезом и накоплением запасных питательных веществ, в первую очередь крахмала, редко белков и липидов. Лейкопласты, накапливающие крахмал, называются амилопластами. Этот крахмал имеет вид зерен, в отличие от ассимиляционного крахмала хлоропластов, он называется запасным, или вторичным. Запасной белок может откладываться в форме кристаллов или аморфных включений в так называемых протеинопластах, жирные масла – в виде пластоглобул в элайопластах. Часто в клетках встречаются лейкопласты, не накапливающие запасные питательные вещества, их роль еще до конца не выяснена. На свету лейкопласты могут превращаться в хлоропласты. Хромопласты —пластиды оранжевого, красного и желтого цвета, который обусловлен пигментами, относящимися к группе каротиноидов. Хромопласты встречаются в клетках лепестков многих растений (ноготки, лютик, одуванчик), зрелых плодов (томат, шиповник, рябина, тыква, арбуз), редко — корнеплодов (морковь), а также в осенних листьях. Внутренняя мембранная система в хромопластах, как правило, отсутствует. Каротиноиды чаще всего растворены в жирных маслах пластоглобул (рис. 2.6), и хромопласты имеют более или менее сферическую форму. В некоторых случаях (корнеплоды моркови, плоды арбуза) каротиноиды откладываются в виде кристаллов различной формы. Кристалл растягивает мембраны хромопласта, и он принимает его форму: зубчатую, игловидную, серповидную, пластинчатую, треугольную, ромбовидную и др. Значение хромопластов до конца еще не выяснено. Большинство из них представляют собой стареющие пластиды. Они, как правило, развиваются из хлоропластов, при этом в пластидах разрушаются хлорофилл и внутренняя мембранная структура, и накапливаются каротиноиды. Это происходит при созревании плодов и пожелтении листьев осенью. Косвенное биологическое значение хромопластов состоит в том, что они обусловливают яркую окраску цветков и плодов, привлекающую насекомых для перекрестного опыления и других животных для распространения плодов. В хромопласты могут превращаться и лейкопласты. Пластиды всех трех типов образуются из пропластид – мелких бесцветных телец, которые находятся в меристематических (делящихся) клетках корней и побегов. Пропластиды способны делиться и по мере дифференциации превращаются в пластиды разного типа. В эволюционном смысле первичным, исходным типом пластид являются хлоропласты, из которых произошли пластиды остальных двух типов. В процессе индивидуального развития (онтогенеза) почти все типы пластид могут превращаться друг в друга. Пластиды имеют много общих черт с митохондриями, отличающих их от других компонентов цитоплазмы. Это, прежде всего, оболочка из двух мембран и относительная генетическая автономность, обусловленная наличием собственных рибосом и ДНК. Такое своеобразие органелл легло в основу представления, что предшественниками пластид и митохондрий были бактерии, которые в процессе эволюции оказались встроенными в эукариотическую клетку и постепенно превратились в хлоропласты и митохондрии. 13.Физиологически активные вещества: В состав содержимого растительной клетки входят также физиологически активные вещества. К ним относятся ферменты, фитогормоны, витамины, антибиотики и фитонциды. Ферменты, или энзимы,—вещества белковой структуры, выполняющие функцию биологических катализаторов (ускорителей) химических реакций. Действуют значительно активнее, чем неорганические катализаторы. Содержатся в цитоплазме и клеточном соке. Обеспечивают управляемый ход .всех химических реакций в клетке — от синтеза до распада. К фитогормонам относятся вещества, обеспечивающие усиление физиологических процессов в «летке. Примером .могут служить гормоны роста (ауксин, гетеро-ауксин и др.). В настоящее время науке известно 40 различных по своим свойствам витаминов, незаменимое значение которых для нормального обмена веществ, роста и развитии растений, животных и человека общеизвестно. Антибиотики и фитонциды — вещества, встречающиеся в клеточном соке и цитоплазме и обладающие бактерицидными свойствами, т. е. способностью убивать бактерии. Широко известно бактерицидное действие пенициллина и других антибиотических препаратов. Фитонцидами богаты некоторые древесные породы (сосна, дуб, черемуха) и травянистые растения (лук, чеснок, горчица). 14.Вакуо́ль: — одномембранный органоид, содержащийся в некоторых эукариотических клетках и выполняющий различные функции (секреция, экскреция и хранение запасных веществ, аутофагия, автолиз и др.). Вакуоли и их содержимое рассматриваются как обособленный от цитоплазмы компартмент. Различают пищеварительные и сократительные (пульсирующие) вакуоли, регулирующие осмотическое давление и служащие для выведения из организма продуктов распада. Вакуоли особенно хорошо заметны в клетках растений: во многих зрелых клетках растений они составляют более половины объёма клетки. Одна из важных функций растительных вакуолей — накопление ионов и поддержание тургора (тургорного давления). Вакуоль — это место запаса воды. Вакуоли развиваются из цистерн эндоплазматической сети. Мембрана, в которую заключена вакуоль, называется тонопласт.В вакуолях содержатся органические кислоты, углеводы, дубильные вещества, неорганические вещества (нитраты, фосфаты, хлориды и др.), белки и др. 15.Клеточный сок: представляет собой водный раствор различных веществ, являющихся продуктами жизнедеятельности протопласта, в основном, запасными веществами и отбросами. Реакция клеточного сока обычно слабокислая или нейтральная, реже щелочная. Вещества, входящие в состав клеточного сока, чрезвычайно разнообразны. Это углеводы, белки, органические кислоты и их соли, аминокислоты, минеральные ионы, алкалоиды, гликозиды, танниды, пигменты и другие растворимые в воде соединения. Большинство из них относится к группе эргастических веществ – продуктов метаболизма протопласта, которые могут появляться и исчезать в различные периоды жизни клетки. Многие вещества клеточного сока образуются только в растительных клетках. Углеводы клеточного сока растений представлены моносахаридами (глюкоза, фруктоза), дисахаридами (сахароза) и полисахаридами (слизи, инулин). Из органических кислот в клеточном соке наиболее часто встречаются лимонная, яблочная, янтарная и щавелевая. Эти кислоты находятся в большом количестве в клеточном соке незрелых плодов, придавая им кислый вкус. При созревании плодов органические кислоты могут использоваться как субстраты дыхания, поэтому кислый вкус плодов обычно исчезает. Соли органических кислот вместе с минеральными ионами играют большую роль в осмотических процессах. 16.Запасные питательные вещества: Запасные питательные вещества накапливаются в клетках растений в течение вегетационного периода и используются частично зимой, а главное, весной на образование молодых побегов и корней. Перед листопадом или завяданием надземных частей многолетних трав резервные вещества оттягиваются в зимующие органы. У однолетников они концентрируются в семенах или плодах. С возобновлением роста или других активных физиологических процессов они мобилизуются, превращаясь из веществ полимерных в более простые органические соединения. Запасные питательные вещества используются двояко — как энергетический и как пластический материал. В первом случае они окисляются в процессе дыхания, и клеткой используется освобождающаяся энергия. Во втором случае из запасных питательных веществ или их производных образуются вещества конституционные, т. е. такие, из которых состоят живые части протопласта. В качестве запасных питательных веществ в клетках встречаются все виды органических соединений — углеводы, белки, липиды. 17и 18.Клеточная оболочка, ее видоизменения (лигнификация, суберинизация, кутинизация, минерализация, ослизнение): Клеточная стенка, обладающая прочностью способна к росту, она прозрачная и хорошо пропускает солнце, легко проникает вода. Основа оболочки составляют молекулы целлюлозы собранные в сложные пучки – фибриллы, образующий каркас, погруженный в основу – матрикс, состоящий из гемицеллюлозы, пектинов, гликопротеидов. Первоначально число фибрилл невелико, но с возрастом они увеличивается и клетка теряет способность к растяжению. В матриксе часто обнаруживается неуглеводный компонент – легнин. Одревеснение клеточной оболочки происходит в результате отложения лигнина, Лигнин повышает устойчивость тканей к разрушительному действию бактерий и грибов. Одревесневшие оболочки не теряют способности пропускать воду. Клетки с одревесневшими стенками могут оставаться живыми, но чаще становятся мертвыми. Стенки некоторых клеток могут включать: воск, кутину, суберин. Функции: придает клетке форму; отделяет одну клетку от другой, является скелетом для каждой клетки и придает прочность всему растению, выполняет защитную функцию. Опробковение вызывается особым жироподобным веществом — суберином. Опробковевшие оболочки становятся непроницаемыми для воды и газов, и содержимое клеток с опробковевшими оболочками отмирает. В местах ранения растения также образуются клетки с опробковевшими стенками, которые отделяют здоровые ткани от поврежденных. Кутинизация заключается в выделении жироподобного вещества кутина. Обычно кутинизируются наружные стенки кожицы листьев и "травянистых стеблей. Это делает их менее проницаемыми для воды, уменьшает испарение у растений. Кутин образует на поверхности органа пленку, называемую кутикулой. . Минерализация клеточных оболочек — это отложение: кремнезема и солей кальция. Наиболее сильно инкрустируются оболочки клеток кожицы листьев и стеблей злаков, осок, хвощей. Листьями злаков и осок можно поранить руки. Ослизнение оболочек – превращение целлюлозы и пектиновых веществ в слизи и камеди. Ослизнение хорошо наблюдается на семенах льна, находившихся в воде. Образование слизей способствует лучшему поглощению воды семенами и прикреплению их к почве. 19.Способы деления эукариотических клеток: митоз, мейоз, амитоз: Митоз — основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала.
Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют митотический цикл. Ниже приводится краткая характеристика фаз цикла.
Интерфаза состоит из трех периодов: пресинтетического, или постмитотического, — G1, синтетического — S, постсинтетического, или премитотического, — G2. Пресинтетический период (2n 2c, где n — число хромосом, с — число молекул ДНК) — рост клетки, активизация процессов биологического синтеза, подготовка к следующему периоду. Синтетический период (2n 4c) — репликация ДНК. Постсинтетический период (2n 4c) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей. Профаза (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом. Метафаза (2n 4c) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом. Анафаза (4n 4c) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами). Телофаза (2n 2c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки. Биологическое значение митоза. Образовавшиеся в результате этого способа деления дочерние клетки являются генетически идентичными материнской. Митоз обеспечивает постоянство хромосомного набора в ряду поколений клеток. Лежит в основе таких процессов, как рост, регенерация, бесполое размножение и др. Мейоз — это особый способ деления эукариотических клеток, в результате которого происходит переход клеток из диплоидного состояния в гаплоидное. Мейоз состоит из двух последовательных делений, которым предшествует однократная репликация ДНК. Первое мейотическое деление (мейоз 1) называется редукционным, поскольку именно во время этого деления происходит уменьшение числа хромосом вдвое: из одной диплоидной клетки (2n 4c) образуются две гаплоидные (1n 2c). Интерфаза 1 (в начале — 2n 2c, в конце — 2n 4c) — синтез и накопление веществ и энергии, необходимых для осуществления обоих делений, увеличение размеров клетки и числа органоидов, удвоение центриолей, репликация ДНК, которая завершается в профазе 1. Профаза 1 (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом, конъюгация гомологичных хромосом и кроссинговер. Конъюгация — процесс сближения и переплетения гомологичных хромосом. Пару конъюгирующих гомологичных хромосом называют бивалентом. Кроссинговер — процесс обмена гомологичными участками между гомологичными хромосомами. Профаза 1 подразделяется на стадии: лептотена (завершение репликации ДНК), зиготена (конъюгация гомологичных хромосом, образование бивалентов), пахитена (кроссинговер, перекомбинация генов), диплотена (выявление хиазм, 1 блок овогенеза у человека), диакинез (терминализация хиазм). Метафаза 1 (2n 4c) — выстраивание бивалентов в экваториальной плоскости клетки, прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом. Анафаза 1 (2n 4c) — случайное независимое расхождение двухроматидных хромосом к противоположным полюсам клетки (из каждой пары гомологичных хромосом одна хромосома отходит к одному полюсу, другая — к другому), перекомбинация хромосом. Телофаза 1 (1n 2c в каждой клетке) — образование ядерных мембран вокруг групп двухроматидных хромосом, деление цитоплазмы. У многих растений клетка из анафазы 1 сразу же переходит в профазу 2. Второе мейотическое деление (мейоз 2) называется эквационным. Интерфаза 2, или интеркинез (1n 2c), представляет собой короткий перерыв между первым и вторым мейотическими делениями, во время которого не происходит репликация ДНК. Характерна для животных клеток. Профаза 2 (1n 2c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления. Метафаза 2 (1n 2c) — выстраивание двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом; 2 блок овогенеза у человека. Анафаза 2 (2n 2с) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами), перекомбинация хромосом. Телофаза 2 (1n 1c в каждой клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия) с образованием в итоге четырех гаплоидных клеток. Биологическое значение мейоза. Мейоз является центральным событием гаметогенеза у животных и спорогенеза у растений. Являясь основой комбинативной изменчивости, мейоз обеспечивает генетическое разнообразие гамет. Амитоз Амитоз — прямое деление интерфазного ядра путем перетяжки без образования хромосом, вне митотического цикла. Описан для стареющих, патологически измененных и обреченных на гибель клеток. После амитоза клетка не способна вернуться в нормальный митотический цикл. Клеточный цикл Клеточный цикл — жизнь клетки от момента ее появления до деления или смерти. Обязательным компонентом клеточного цикла является митотический цикл, который включает в себя период подготовки к делению и собственно митоз. Кроме этого, в жизненном цикле имеются периоды покоя, во время которых клетка выполняет свойственные ей функции и избирает дальнейшую судьбу: гибель или возврат в митотический цикл. 20.Общая характеристика и классификация растительных тканей: Ткань – это группа клеток, которые имеют общее происхождение, выполняют одну или несколько функций и занимают свойственное им положение в органах растения. Ткани делят на простые и сложные. Простые — состоящие из клеток более или менее одинаковых по форме и функциям. Сложные — из клеток, разных по форме и функциям, но тесно взаимосвязанных. Ткани делят на образовательные (меристемы) и постоянные. Образовательные — клетки сохраняют длительную способность к делению, обеспечивая рост растения и отдельных его органов. С учетом положения в теле растения их делят на верхушечные (находятся на апексах корня и побега), вставочные (свойственны побегу – стеблю и листьям) и боковые (представлены главным образом в осевых органах – в корне и стебле голосемянных и двудольных покрытосемянных).Постоянные — клетки которых утратили способность к делению и выполняют другие функции: защитной, запасающей, механической, проводящей и т.д. С учетом происхождения, преобладающей функции и положения в теле растения постоянные ткани, в свою очередь, делят на покровные, основные, проводящие. 21. Образовательные ткани. Строение и месторасположение клеток меристем, функции ткани: Образовательная ткань (меристема) дают начало всем постоянным тканям, обеспечивают рост органов. По происхождению они могут быть первичными ( производными зародышевых тканей семени ) или вторичными , возникающими позже . По топографии ( расположению в теле растений ) – верхушечными , боковыми , вставочными . Апикальные, или верхушечные, меристемы возникают в зародыше семени, сохраняются в апексах — на кончике корня и на верхушке побега, обеспечивают рост органов в длину. Латеральные, или боковые, меристемы располагаются вдоль тела растений , формируют в основном проводящую систему ( проводящий цилиндр ) и вызывают рост органов в ширину К ним относятся: первичные — прокамбий, перицикл и вторичные — камбий, феллоген (пробковый камбий). Прокамбий образует первичные проводящие ткани : I луб ( флоэму ) и I древесину ( ксилему ), камбий – II луб и II древесину. Пробковый камбий ( феллоген) формирует вторичную покровную ткань – перидерму Интеркалярные, или вставочные, меристемы первичны. Это остатки апикальных меристем . Обеспечивают рост черешков , тычиночных нитей , междоузлий и т . д . Травматические, или раневые, меристемы вторичны, возникают в местах повреждения. Деление меристем стимулируют фитогормоны. Клетки меристемы паренхимные, живые, тонкостенные, плотно сомкнутые, с густой цитоплазмой, крупным ядром, большим количеством рибосом. Пластиды в форме пропластид и лейкопластов, вакуоли отсутствуют или очень мелкие. Те клетки меристемы, что находятся в состоянии постоянного деления, называются инициалями, а те, что образуются из них и подвергаются дифференцировке, это производные инициалей, или основная меристема.
|
Источник: studopedya.ru
6.Общая характеристика и классификация растительных тканей
Ткань – это группа клеток, которые имеют общее происхождение, выполняют одну или несколько функций и занимают свойственное им положение в органах растения. Ткани делят на простые и сложные. Простые — состоящие из клеток более или менее одинаковых по форме и функциям. Сложные — из клеток, разных по форме и функциям, но тесно взаимосвязанных. Ткани делят на образовательные (меристемы) и постоянные. Образовательные — клетки сохраняют длительную способность к делению, обеспечивая рост растения и отдельных его органов. С учетом положения в теле растения их делят на верхушечные (находятся на апексах корня и побега), вставочные (свойственны побегу – стеблю и листьям) и боковые (представлены главным образом в осевых органах – в корне и стебле голосемянных и двудольных покрытосемянных).Постоянные — клетки которых утратили способность к делению и выполняют другие функции: защитной, запасающей, механической, проводящей и т.д. С учетом происхождения, преобладающей функции и положения в теле растения постоянные ткани, в свою очередь, делят на покровные, основные, проводящие.
7.Основные ткани растений, их характеристика и функции.++
занимают наибольший объем. Снаружи они защищены покрывными тканями, а изнутри пронизаны проводящими и механическими пучками. Образующие их клетки отличаются обычно паренхимным строением, из-за этого основные ткани нередко называют основной паренхимой. Основные ткани являются питающими тканями, хотя могут выполнять многие функции, начиная от ассимиляционной, кончая выделительной. В зависимости от специализации, основную паренхиму делят на ассимиляционную, вентиляционную, всасывающую, запасающую и выделительную .Ассимиляционные (фотосинтези-рующие) ткани они имеют зеленую окраску – хлоренхима. Ассим. ткань состоит из однородных тонкостенных клеток. Доступ углекислоты к клеткам облегчается тем, что в ней имеется развитая система межклетников, сообщающаяся с атмосферой. Наличие межклетников является характернейшей особенностью ассимиляционной ткани. Межклетники обеспечивают газообмен с окружающей средой. Хлоренхима располагается в местах, наиболее доступных свету: в надземной части растений непосредственно под кожицей листьев и стеблей. Нередко хлоренхима дифференцирована на столбчатую (палисадную) и губчатую ткань. Палисадная ткань содержит большое количество хлоропластов, здесь происходят световые реакции фотосинтеза. Губчатая ткань- протекает газообмен и темновая стадия фотосинтеза. Вентиляционная ткань (Аэренхима) во всех тканях имеются межклетники, образующие единую систему. Межклетники через проходные отверстия или устьица сообщаются с атмосферой. Однако газовый состав в межклетниках сильно отличается от газового состава атмосферы. Запасающие ткани Поглощенные растением синтезированные вещества могут откладываться в виде запасов: накопление воды, пластические вещества.сахар, инулин, аминокислоты, белки, крахмал.
8. Образовательные ткани. Строение и месторасположение клеток меристем, функции ткани ++
Образовательная ткань (меристема) дают начало всем постоянным тканям, обеспечивают рост органов. По происхождению они могут быть первичными ( производными зародышевых тканей семени ) или вторичными , возникающими позже . По топографии ( расположению в теле растений ) – верхушечными , боковыми , вставочными . Апикальные, или верхушечные, меристемы возникают в зародыше семени, сохраняются в апексах — на кончике корня и на верхушке побега, обеспечивают рост органов в длину. Латеральные, или боковые, меристемы располагаются вдоль тела растений , формируют в основном проводящую систему ( проводящий цилиндр ) и вызывают рост органов в ширину К ним относятся: первичные — прокамбий, перицикл и вторичные — камбий, феллоген (пробковый камбий). Прокамбий образует первичные проводящие ткани : I луб ( флоэму ) и I древесину ( ксилему ), камбий – II луб и II древесину. Пробковый камбий ( феллоген) формирует вторичную покровную ткань – перидерму Интеркалярные, или вставочные, меристемы первичны. Это остатки апикальных меристем . Обеспечивают рост черешков , тычиночных нитей , междоузлий и т . д . Травматические, или раневые, меристемы вторичны, возникают в местах повреждения. Деление меристем стимулируют фитогормоны. Клетки меристемы паренхимные, живые, тонкостенные, плотно сомкнутые, с густой цитоплазмой, крупным ядром, большим количеством рибосом. Пластиды в форме пропластид и лейкопластов, вакуоли отсутствуют или очень мелкие. Те клетки меристемы, что находятся в состоянии постоянного деления, называются инициалями, а те, что образуются из них и подвергаются дифференцировке, это производные инициалей, или основная меристема.
Источник: StudFiles.net
Клетки растений весьма разнообразны по форме и размерам. Форма и величина их определяется местом положения в организме растения, а также выполняемыми функциями. Клетки, входящие в состав различных тканей и органов, значительно различаются по ширине и длине, но чаще всего они вытянуты и имеют заостренные концы. Например, длина клетки покрытосеменных колеблется в пределах от 100 до 1000 мкм. Паренхимные клетки плодов и клубней растений достигают более 1 мм. Большие размеры имеют клетки лубяных волокон. Так, у льна и копли длина волокна составляет 20-40 мм, а у хлопчатника – 65 мм. Однако, чаще всего клетки мелкие, величиной 20-50 мкм и их можно видеть только под микроскопом.
Общая численность клеток, составляющих растение, выражается астрономическими цифрами, достигая нескольких сотен и тысяч миллиардов.
Клетка растений состоит из двух основных структур – цитоплазмы и ядра. Цитоплазма ( от греческого cytos – клетка, plasma – первичная масса) и ядро тесно связаны между собой и представляют единую живую систему. Цитоплазма без ядра существовать не может, так же как и ядро без цитоплазмы. В начале развития учения о клетке чешский физиолог Пуркинье (1839) назвал содержимое клетки протоплазмой (от греческого protos – первый). Клетка одета клеточной оболочкой (мембраной), состоящей из клетчатки и пектиновых веществ (целлюлоза, лигнин, воск и др.). мембрана имеет поры через которые вещества могут проникать из одной клетки в другую.
Основную часть объема клетки занимает цитоплазма. Слой цитоплазмы, прилегающий к оболочке, называется экзоплазмой. Он более вязкий и лишен гранул по сравнению с внутренним слоем, называемым эндоплазмой. В электронном микроскопе цитоплазма представляется однородной зернистой массой. Она состоит из прозрачного вещества гиалоплазмы (от греческого hyalos – стекло и плазма) и взвешенных в ней мельчайших частиц – гранул. Гиалоплазму называли вначале матриксом. В цитоплазме находятся органоиды и включения, протекают биохимические реакции, осуществляется транспорт веществ.
В состав цитоплазмы входит 10-20 % белков, 2-3 % липидов, 1-2 % углеводов и примерно 1 % минеральных солей и других веществ. В водной среде цитоплазмы растворены минеральные вещества и находятся органические соединения, представленные двумя группами: полупродукты синтеза и распада (аминокислоты, моносахариды, глицерин, жирные кислоты, азотистые основания и др.) и конечными продуктами синтеза (белки, углеводы, липиды, нуклеиновые кислоты, ферменты, витамины и др.). Наибольшее значение в жизнедеятельности растительной клетки играют белки. Недаром голландский ученый-химик И. Мудлер назвал белки протеинами, что в переводе означает первостепенно важные. Белки выполняют структурную роль, являются составной частью витаминов, ферментов, гормонов, участвуют в многочисленных реакциях обмена, имеют первостепенное значение в защитных реакциях и т. д.
Липиды являются главным образом запасными веществами клетки. Они – источник энергии. Некоторые липиды входят в состав ядерных и клеточных оболочек и многочисленных мембран.
Углеводы представлены в цитоплазме в виде моно и дисахаридов. В состав клетки входит также крахмал, играющий роль запасного вещества. Источником внутриклеточной энергии является глюкоза. Исключительно важное значение имеет рибоза и дезоксирибоза. Первая входит в состав рибонуклеиновой кислоты (РНК), вторая – дезоксирибонуклеиновой кислоты (ДНК).
Минеральные вещества находятся в цитоплазме в виде свободных соединений и в связанном состоянии с белками, жирами и углеводами.
Вода входит в состав коллоидов цитоплазмы, она обеспечивает процессы гидролиза и окисления веществ.
Ядро покоится в цитоплазме. Впервые его обнаружил английский ученый Р. Броун (1831), рассматривая под микроскопом клетки растений из семейства орхидных. Ядро – важнейший и постоянный компонент всех эукариотических клеток. исключительная важность ядра для жизнедеятельности клеток доказана опытным путем, например, с одноклеточной водорослью ацетобулярией. Клетка водоросли состоит из шляпки и ножки длиной 4-6 см. шляпка содержит цитоплазму, а ядро находится в нижней части ножки. При отделении шляпки от ножки она погибает, а ножка в которой находится ядро. Продолжает жить и образовывать шляпку, т.е. часть растения, содержащая ядро обладает способностью регенерации.
Ядра клеток разнообразны по форме и размерам. Обычно форма ядер связана с формой клеток, но иногда отличается от последней. В основном ядра имеют округлую или овальную форму. У большинства растений размер их колеблется в пределах 10-20 мкм. Форма и величина ядра зависит от возраста клетки, физиологического состояния и факторов внешней среды. Ядро клетки занимает около 1/5 ее объема. В нем различают ядерную оболочку, ядерный сок (кариолимфу), хроматин и ядрышки. Ядро отграничено от цитоплазмы ядерной оболочкой (мембраной), которая состоит из внутреннего сплошного и внешнего пористого листков. Листки мембраны представлены в основном протеинами и липидами. Главная функция ядерной мембраны – регулирование поступления веществ из цитоплазмы в ядро и обратно.
Ядерный сок представляет собой жидкое или полужидкое содержимое ядра. Кариолимф а и содержащиеся в ней глыбки хроматина (от греческого chroma – цвет) называется хромоплазмой. Субмикроскопический состав хромоплазмы аналогичен составу цитоплазмы. Электронной микроскопией в ней выявлены тонкие нити и гранулы.
В ядре клеток обнаруживается 1-2 ядрышка, которые содержат большое количество РНК. Методом авторадиографии установлено, что РНК ядрышков проникает в рибосомы цитоплазмы и принимает активное участие в синтезе белков.
Для химического состава ядра характерно наличие большого количества ДНК и белков-гистонов. ДНК входит в состав хромосом, являющихся компонентами ядра и материальными носителями наследственности.
Нуклеиновые кислоты впервые обнаружил швейцарский биохимик Мишер (1869) в ядрах животных клеток. Название их происходит от латинского nucleus – ядро.
Ядро является центром, управляющим всеми процессами жизнедеятельности клетки, в нем сосредоточены материальные носители наследственности всех признаков организма.
Цитоплазма растительных клеток представляет собой довольно сложную структурную систему.
Электронная микроскопия позволила выявить, что она – совокупность коротких и длинных, узких и широких, замкнутых и не замкнутых внутренних мембран и канальцев была названна эндоплазмотической сетью, которая открыта в 1945 г (Портер, Клод и др.). Реальное существование ее окончательно доказано к 1958 году. Внутренне пространство эндоплазмотической сети заполненное гомогенным веществом, состав которого остается малоизученным.
Различают две разновидности эндоплазмотической сети: гранулярную и агранулярную. Первая характеризуется наличием на поверхности мембран мелких гранул, получивших название рибосом. Гранулярная эндоплазмотическая сеть участвует в синтезе белков, а агранулярная – в синтезе липидов и углеводов. Эндоплазмотическя сеть связана со всеми структурами клетки. Ее оценивают как органоид общего значения, участвующий в процессах синтеза, обмене веществ, обеспечивающий взаимосвязь элементов клетки между собой и с окружающей средой.
В цитоплазме всех растительных клеток имеются мелкие частицы рибосомы ( от греческого soma – тело и от начала слова рибонуклеиновая кислота), которые можно видеть только в электронный микроскоп. Они свободно располагаются в цитоплазме или прикреплены к мембране эндоплазмотической сети и ядерной оболочке. Иногда рибосомы располагаются в виде скоплений (от 5 до 70). Такие группы рибосом получили название полисом или полирибосом. Рибосомы состоят из равного количества белка и РНК. В незначительном количестве в них обнаружены соли магния и кальция. РНК рибосом составляет 80-90 % от общего количества этой кислоты, содержащегося в этой клетке.
Рибосомы обеспечивают процессы внутриклеточного синтеза белка. Их называют своеобразными «фабриками» белка, на «конвейерах», которых происходит сборка из аминокислот белковых молекул. Белок, синтезированный рибосомами, поступает в каналы эндоплазмотической сети, а затем во все органоиды клетки, в том числе и ее ядро. Рибосомы обладают высокой синтезирующей способностью, производя за 1 час белка больше своего веса.
Митохондрии (от греческого mitos – нить, chondros – зерно) – органоиды клетки, ее «силовые станции». Их можно обнаружить в обычный световой микроскоп. Длина митохондрий составляет – 0,5-0,7 мкм, ширина – 0,5-1 мкм. Количество митохондрий в клетке зависит от ее функционального состояния и возраста. В среднем число их колеблется от 2 до 2,5 тысяч. Митохондрии имеют двойную оболочку, которая состоит из наружной и внутренней мембран. Жидкое содержимое митохондрий называется матриксом. Внутренняя мембрана имеет складки, называемые кристаллами. Состоят митохондрии из белка (65-70 %), липидов (25-30 %) и небольшого количества РНК и ДНК.
Основная роль митохондрий заключается в синтезе аденозинтрифосфорной кислоты (АТФ), которая является универсальным источником энергии, которая необходима для обеспечения жизнедеятельности клетки и организма вцелом.
Комплекс Гольджи — сложная структура, состоящая из мембран, гранул и вакуолей. Впервые ее образование открыто итальянским ученым К. Гольджи (1898) и названо его именем. В растительных клетках комплекс Гольджи имеет вид дискретных частиц, равномерно рассеяных по всей цитоплазме. Полагают, что комплекс Гольджи накапливает различного рода ненужные клетке продукты ее жизнедеятельности и избытки воды, подлежащие удалению, т.е. способствует регуляции определенного уровня концентрации веществ в клетке.
Пластиды ( от греческого plastos – вылепленный, eidos – подобный) – органоиды. Присущие клеткам растений. Они устроены очень сложно, способны к самовоспроизведению, тесно связаны со способом питания растений. Пластиды имеют общее происхождение и могу превращаться друг в друга. Различают следующие типы пластид: хлоропласты, хромопласты, лейкопласты.
Хлоропласты содержат пигмент хлорофилл, осуществляющий процесс фотосинтеза, обуславливает зеленый цвет растения. Хромопласты содержат каратиноиды. Обуславливают оранжевую, желтую, красную окраску растений. Каратиноиды выполняют важную роль в процессах обмена веществ в клетке. Лейкопласты ( бесцветные пластиды) являются органоидами. Синтезирующими и запасающими крахмал.
Источник: biofile.ru