Растения образующие споры называются
Включают следующие ныне существующие отделы: моховидные (Bryophyta), плауновидные (Lycopodiophyta), псилотовидные (Psilotophyta), хвощевидные (Equisetophyta), папоротниковидные (Polypodiophyta).
Споровые растения появились в конце силурийского периода, более 400 млн. лет тому назад. Первые представители споровых были небольших размеров и имели простое строение, но уже у примитивных растений наблюдалась дифференциация на элементарные органы. Усовершенствованию органов соответствовало усложнение внутреннего строения и онтогенеза. В жизненном цикле происходит чередование полового и бесполого способов размножения и связанное с этим чередование поколений. Бесполое поколение представлено диплоидным спорофитом, половое – гаплоидным гаметофитом.
На спорофитеобразуются спорангии, внутри которых в результате мейотического деления образуются гаплоидные споры. Это мелкие, одноклеточные образования, лишенные жгутиков. Растения, у которых все споры одинаковые, называются равноспоровыми. У более высокоорганизованных групп споры двух типов: микроспоры (формируются в микроспорангиях), мегаспоры (формируются в мегаспорангиях). Это разноспоровые растения. При прорастании споры образуется гаметофит.
Полный жизненный цикл (от зиготы до зиготы) состоит из гаметофита(период от споры до зиготы) и спорофита (период от зиготы до образования спор). У плаунов, хвощей и папоротников эти фазы представляют собой как бы отдельные физиологически самостоятельные организмы. У мхов гаметофит является самостоятельной фазой жизненного цикла, а спорофит сведен до его своеобразного органа – спорогона(спорофит живет на гаметофите).
На гаметофите развиваются органы полового размножения: архегонии и антеридии. В архегониях, похожих на колбу, образуются яйцеклетки, а в мешковидных антеридиях — сперматозоиды. У равноспоровых растений гаметофиты обоеполые, у разноспоровых – однополые. Оплодотворение происходит лишь при наличии воды. При слиянии гамет образуется новая клетка – зигота с двойным набором хромосом (2n).
Отдел Моховидные – Bryophyta
Насчитывает до 27000 видов. Моховидные имеют тело либо в виде слоевища, либо расчленены на стебель и листья. Настоящих корней у них нет, их заменяют ризоиды. Проводящие ткани появляются лишь у высокоразвитых мхов. Частично обособлены ассимиляционная и механическая ткани.
В жизненном цикле доминирует гаметофит. Спорофит самостоятельно не существует, развивается и всегда находится на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий, на ножке, связывающей ее с гаметофитом.
Размножаются мхи спорами, могут размножаться и вегетативным путем – отдельными участками тела или особыми выводковыми почками.
Отдел делится на три класса: Антоцеротовые (100 видов, шесть родов талломных растений), Печеночные и Листостебельные мхи.
Класс Печеночные мхи (Hepaticopsida)
Класс насчитывает около 8500 видов. Это, в основном, слоевищные мхи, хотя есть виды, имеющие стебель и листья. Широко распространена маршанция обыкновенная (Marchantia polymorpha) (рис 11. 1).

Рис. 11. 1. Цикл воспроизведения маршанции: 1– таллом с мужскими подставками; 2 – таллом с женскими подставками; 3 – вертикальный разрез через мужскую подставку (в некоторых антеридиальных полостях находятся антеридии); 4 – антеридий в антеридиальной полости (н – ножка антеридия); 5 – двужгутиковый сперматозоид; 6– вертикальный разрез через женскую подставку (а – архегоний).
Гаметофитимеет темно-зеленое слоевище (таллом), дихотомически разветвленное на широкие лопастные пластинки с дорсовентральной (спинно-брюшной) симметрией. Сверху и снизу слоевище покрыто эпидермой, внутри имеются ассимиляционная ткань и клетки, выполняющие проводящую и запасающую функции. К субстрату слоевище прикрепляется ризоидами. На верхней стороне слоевища в особых «корзиночках» образуются выводковые почки, служащие для вегетативного размножения.
Талломы раздельнополые, органы полового размножения развиваются на особых вертикальных ветвях-подставках.
Мужские гаметофиты имеют восьмилопастные подставки, на верхней стороне которых находятся антеридии. На женских гаметофитах подставки со звездчатыми дисками, на нижней стороне лучей звездочки расположены (шейкой вниз) архегонии. При наличии воды сперматозоиды перемещаются, попадают в архегоний и сливаются с яйцеклеткой.
После оплодотворения из зиготы развивается спорогон. Он имеет вид шаровидной коробочки на короткой ножке. Внутри коробочки, в результате мейоза, из спорогенной ткани, образуются споры. В благоприятных условиях споры прорастают, из них развивается протонема в виде небольшой нити, из верхушечной клетки которой развивается таллом маршанции.
Класс Листостебельные мхи (Bryopsida, или Musci).
Листостебельные мхи распространены по всему земному шару, особенно в условиях холодного климата на сырых местах, в сосновых и еловых лесах, в тундре. На торфяных и моховых болотах часто образуют плотный ковер. Тело расчленено на стебель и листья, но настоящих корней нет, есть многоклеточные ризоиды. Класс состоит из трех подклассов: Бриевые, или Зеленые мхи; Сфагновые, или Белые мхи; Андреевые, или Черные мхи.
Андреевые мхи (три рода, 90 видов) распространены в холодных областях, внешне сходны с зелеными, по строению листьев и коробочки – со сфагновыми.
Подкласс Бриевые, или Зеленые мхи (Bryidae). Насчитывает около 700 родов, объединяющих 14000 видов, широко распространенных повсюду, особенно в тундровой и лесной зонах Северного полушария.
Широко распространен кукушкин лен (Polytrichium commune), образующий густые дерновины на сырых почвах в лесах, на болотах и лугах. Стебли до40 см высоты, неветвистые, с густыми жесткими и острыми листьями. От нижней части стебля отходят ризоиды.
Цикл развития кукушкина льна (рис. 11. 2).
Рис. 11. 2. Кукушкин лен: А– цикл развития мха; Б – коробочка: 1 – с колпачком, 2 – без колпачка, 3 – в разрезе (а –крышечка, б – урночка, в – спорангий, г – апофиза, д – ножка); В – поперечный разрез листа с ассимиляторами; Г – поперечный разрез стебля (ф – флоэма, крв – крахмалоносное влагалище, кор – кора, э – эпидерма, лс – листовые следы).
Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии.
Весной во время дождя или после росы сперматозоиды выходят из антеридия и проникают в архегоний, где сливаются с яйцеклеткой. Из зиготы здесь же, на верхушке женского гаметофита, вырастает спорофит (спорогон), имеющий вид коробочки на длинной ножке. Коробочка покрыта волосистым колпачком (калиптра) (остатком архегония). В коробочке – спорангий, где после мейоза образуются споры. Спора – небольшая клетка с двумя оболочками. На верхушке коробочки, по ее краю, расположены зубцы (перистом), которые, в зависимости от влажности воздуха, загибаются внутрь коробочки или отгибаются наружу, чем способствуют рассеиванию спор. Споры разносятся ветром и в благоприятных условиях прорастают, образуя протонему. Через некоторое время на протонеме образуются почки, из которых формируются листостебельные побеги. Эти побеги вместе с протонемой – гаплоидное поколение – гаметофит. Коробочка на ножке – диплоидное поколение – спорофит.
Подкласс Сфагновые, или Белые мхи (Sphagnidae)
К сфагновым мхам относятся свыше 300 видов единственного рода сфагнум(Sphagnum) (рис. 11. 3).
Рис 11. 3. Сфагнум: 1 – внешний вид; 2 – верхушка ветви со спорогоном; 3 – спорогон ( ш – остаток шейки архегония, кр – крышечка, сп – спорангий, кол – колонка, н – ножка спорогона, лн – ложная ножка); 4 – часть веточного листа ( хлк – хлорофиллоносные клетки, вк – водоносные клетки, п – поры); 5 – поперечный разрез листа.
Ветвистые стебли сфагнума усеяны мелкими листьями. На верхушке главной оси боковые веточки образуют розетку почковидной формы. Особенностью сфагновых мхов является непрерывное нарастание стебля верхушкой и отмирание нижней части. Ризоиды отсутствуют, и поглощение воды с минеральными веществами происходит стеблями. Листья этих мхов состоят из двух типов клеток: 1) живых ассимилирующих, длинных и узких, хлорофиллоносных; 2) гиалиновых – мертвых, лишенных протопласта. Гиалиновые клетки легко заполняются водой и долго ее сохраняют. Благодаря такому строению, сфагновые мхи могут накапливать воды в 37 раз больше своей сухой массы. Разрастаясь плотными дернинами, сфагновые мхи способствуют заболачиванию почв. На болотах наслаивание отмерших частей мха приводит к образованию торфяников. Путем сухой перегонки из торфа получают воск, парафин, фенолы, аммиак; путем гидролиза – спирт. Торфяные плиты являются хорошим теплоизоляционным материалом. Сфагновые мхи обладают бактерицидными свойствами.
Отдел Плауновидные – Lycopodiophyta
Появление плауновидных связывают с силурийским периодом палеозойской эры. В настоящее время отдел представлен травянистыми растениями со стелющимися, дихотомически ветвящимися стеблями и корнями, а также спирально расположенными чешуйчатыми листьями. Листья возникли как выросты на стебле, и их называют микрофиллы. У плаунов есть флоэма, ксилема, перицикл.
Выделяют два современных класса: равноспоровые Плауновые и разноспоровые Полушниковые.
Класс Плауновые (Lycopodiopsida)
Из всего класса до настоящего времени сохранилось четыре рода.
Род плаун(Lycopodium). К этому роду относят многочисленные (около 200 видов) многолетние вечнозеленые травы, распространенные от арктических областей до тропиков. Так плаун булавовидный (L. clavatum) встречается в травяном покрове хвойных лесов на достаточно влажных, но бедных гумусом почвах. В сырых хвойных лесах широко распространен плаун годичный (L. annotinum) (рис. 11. 4).
Рис. 11. 4. Плаун булавовидный.
Родбаранец(Huperzia).Представитель рода —баранец обыкновенный (H. selago) распространен в тундровой, лесотундровой и северной лесной зонах и произрастает в южно-таежных еловых лесах и ольшаниках, а также в мшистых лесах и на альпийских лугах.
Роддифазиаструм (Diphasiastrum). Представитель рода дифазиаструм сплюснутый (D. complanatum) растет на сухих песчаных почвах в сосновых лесах.
Цикл развития на примере плауна булавовидного (рис. 11. 5).
Рис. 11. 5. Цикл развития плауна булавовидного:1 – спорофит; 2 – спорофилл со спорангием; 3 – спора; 4 – гаметофит с антеридиями и архегониями; 5 – развивающийся на гаметофите из зародыша молодой спорофит.
Стелющиеся побеги плауна булавовидного достигают до 25 см в высоту и более 3 м в длину. Стебли покрыты спирально расположенными ланцетно-линейными мелкими листьями. В конце лета на боковых побегах образуется обычно два спороносных колоска. Каждый колосок состоит из оси и мелких тонких спорофиллов – видоизмененных листьев, в основании которых расположены спорангии почковидной формы.
В спорангиях после редукционного деления клеток спорогенной ткани образуются одинаковой величины, одетые толстой желтой оболочкой гаплоидные споры. Они прорастают после периода покоя через 3-8 лет в обоеполые заростки, представляющие собой половое поколение и живущие сапротрофно в почве, в виде клубенька.
нижней поверхности отходят ризоиды. Через них в заросток врастают гифы гриба, образуя микоризу. В симбиозе с грибом, который обеспечивает питание, живет заросток, лишенный хлорофилла и неспособный к фотосинтезу. Заростки многолетние, развиваются очень медленно, лишь через 6-15 лет на них образуются архегонии и антеридии. Оплодотворение совершается при наличии воды. После оплодотворения яйцеклетки двужгутиковым сперматозоидом образуется зигота, которая без периода покоя прорастает в зародыш, развивающийся во взрослое растение.
В официнальной медицине споры плауновприменялись в качестве детской присыпки и обсыпки для пилюль. Побеги баранца обыкновенного используют для лечения больных, страдающих хроническим алкоголизмом.
Класс Полушниковые (Isoetopsida)
Селагинелла (Selaginella) среди современных родов насчитывает самое большое (около 700) число видов.
Это нежное многолетнее травянистое растение, требующее высокой влажности. Селагинеллы в отличие от плаунов характеризуются разноспоровостью. В спороносных колосках образуется два вида спор – четыре мегаспорыв мегаспорангиях и многочисленные микроспоры в микроспорангиях. Из микроспоры образуется мужской гаметофит, состоящий из одной ризоидальной клетки и антеридия со сперматозоидами. Мегаспора развивается в женский гаметофит, не покидающий ее оболочки и состоящий из мелкоклеточной ткани, в которую погружены архегонии. После оплодотворения из яйцеклетки развивается зародыш, и затем новый спорофит.
Отдел Хвощевидные – Equisetophyta
Появились хвощи в верхнем девоне, достигли наибольшего разнообразия в карбоне, когда древесный ярус заболоченных тропических лесов в значительной степени состоял из древовидных хвощевидных, вымерших к началу мезозоя. Современные хвощевидные появились на Земле с мелового периода.
До настоящего времени сохранился лишь один род – хвощ (Equisetum), представленный 30-35 видами, распространенными на всех континентах.
У всех видов хвоща стебли имеют членистое строение с выраженным чередованием узлов и междоузлий. Листья редуцированы до чешуй и располагаются мутовками в узлах. Здесь же образуются и боковые ветви. Ассимилирующую функцию выполняют зеленые стебли, поверхность которых увеличивается ребристостью, стенки клеток эпидермы пропитаны кремнеземом. Подземная часть представлена сильно развитым корневищем, в узлах которого формируются придаточные корни. У хвоща полевого(Equisetum arvense) боковые ветви корневища служат местом отложения запасных веществ, а также органами вегетативного размножения (рис. 11. 6).
Рис. 11. 6. Хвощ полевой: а, б – вегетативный и спороносный побеги спорофита; в – спорангиофор со спорангиями; г, д – споры; е – мужской гаметофит с антеридиями; ж – сперматозоид; з – обоеполый гаметофит; и — архегонии.
Весной на обычных или специальных спороносных стеблях образуются колоски, состоящие из оси, которая несет особые структуры, имеющие вид шестигранных щитков (спорангиофоры). Последние несут 6-8 спорангиев. Внутри спорангиев образуются споры, одетые толстой оболочкой, снабженной гигроскопичными лентовидными выростами – элатерами. Благодаря элатерамспоры сцепляются вместе в комки, хлопья. Групповое распространение спор способствует тому, что при их прорастании разнополые заростки оказываются рядом, и это облегчает оплодотворение.
Заростки имеют вид небольшой длинно-лопастной зеленой пластинки с ризоидами на нижней поверхности. Мужские заростки меньше женских и несут по краям лопастей антеридии с многожгутиковыми сперматозоидами. На женских заростках в средней части развиваются архегонии. Оплодотворение происходит при наличии воды. Из зиготы развивается зародыш нового растения – спорофита.
В настоящее время большой роли в образовании растительного покрова хвощи не имеют. В лесах, на избыточно увлажненной почве, широко распространен хвощ лесной (E. sylvaticum) с сильно ветвящимися, поникающими боковыми веточками. На лугах, паровых полях, в посевах встречается трудно искореняемый сорняк хвощ полевой (E. arvense). У этого хвоща рано весной появляются неветвящиеся побеги, несущие спороносные колоски. Позднее от корневища развиваются зеленые вегетативные побеги. В лесной зоне на песчаных почвах и в оврагах широко распространен хвощ зимующий(E. hyemale).
Вегетативные побеги хвоща полевого (E. arvense) в официнальной медицине применяют: в качестве мочегонного средства при отеках на почве сердечной недостаточности; при заболеваниях мочевого пузыря и мочевыводящих путей; как кровоостанавливающее средство при маточных кровотечениях; при некоторых формах туберкулеза.
Отдел Папоротниковидные – Polypodiophyta
Возникли папоротники в девоне, когда древовидные папоротники вместе с ныне ископаемыми плаунами и хвощами господствовали в растительном покрове земли. Большинство их вымерло, оставшиеся дали начало мезозойским формам, которые были представлены очень широко. Папоротники намного превосходят по числу современных видов все остальные отделы высших споровых (около 25000).
У большинства живущих сейчас (исключая тропические) папоротников отсутствует наземный прямостоячий стебель, но есть подземный в виде корневища. От корневища отходят придаточные корни и крупные листья (вайи), имеющие стеблевое происхождение и длительно нарастающие верхушкой. Молодые листья обычно свернуты «улиткой». Среди существующих в настоящее время папоротников имеются как равноспоровые, так и разноспоровые.
В лесах Австралии, Южной Америки, Азии произрастают древовидные представители с колонновидными, неветвящимися, до 20 метров высоты стволами. В средней полосе нашей страны папоротники – это многолетние корневищные травы. Многие папоротники, как и мхи, являются индикаторами почв и типов леса. В светлых лесах, на песчаных или сухих подзолистых почвах распространен орляк обыкновенный(Pteridium aquilinum); на влажных богатых почвах кочедыжники(Athyrium) и крупныелесныещитовники (Dryopteris)(рис. 11. 7).
Рис. 11. 7. Щитовник мужской: А – спорофит: а – общий вид; б – сорусы на нижней стороне вайи; в – разрез соруса (1 – индузий, 2 – планцента, 3 – спорангий); г – спорангий (4 – кольцо); Б– гаметофит: 5 – сперматозоиды; 6 – заросток с нижней стороны (т – таллом, р – ризоиды, арх — архегонии, ан – антеридии); 7 – выход сперматозоидов из антеридия; 8 – архегоний с яйцеклеткой.
Цикл развития равноспоровых папоротников
В середине лета на нижней стороне зеленых листьев (у некоторых на специальных спороносных листьях) в виде коричневых бородавочек появляются группы спорангиев (сорусы). Сорусы у многих папоротников прикрыты сверху своеобразным покрывалом — индузием.Спорангии образуются на особом выросте листа (плаценте)и имеют чечевицеобразную форму, длинные ножки и многоклеточные стенки. В спорангиях хорошо выражено механическое кольцо, которое имеет вид узкой несмыкающейся полосы, опоясывающей спорангий. Когда кольцо подсыхает, стенки спорангия разрываются, и споры высыпаются.
Споры, образовавшиеся в спорангиях, одноклеточны и имеют толстую оболочку. При созревании они разносятся током воздуха и в благоприятных условиях прорастают, образуя сердцевидную зеленую многоклеточную пластинку (заросток), прикрепляющуюся к почве ризоидами. Заросток представляет собой половое поколение папоротников (гаметофит). На нижней стороне заростка образуются антеридии (со сперматозоидами) и архегонии (с яйцеклетками). При наличии воды сперматозоиды проникают в архегонии и оплодотворяют яйцеклетки. Из зиготы развивается зародыш, имеющий все основные органы (корень, стебель, лист и особый орган – ножку, прикрепляющую его к заростку). Постепенно зародыш начинает существовать самостоятельно, а заросток отмирает.
У разноспоровых папоротников гаметофиты редуцированы до микроскопических размеров (особенно мужские).
Из корневищпапоротника мужского(Dryopteris filix-mas), получают густой экстракт, который является эффективным противоглистным средством (ленточные черви).
Источник: studopedia.ru
Высшие растения
К высшим растениям (Emryobionta) принадлежат высшие споровые и семенные растения. Известно свыше 300 тыс. видов. Эти растения вышли на сушу и приспособились к новому месту обитания. Появляются вегетативные органы (корень и побег). Все высшие растения имеют одинаковые фотосинтетические пигменты (хлорофилл а и b, каротиноиды). Большинство высших растений имеют сложную транспортную систему веществ, могут регулировать испарение воды листьями. Клетки высших растений образуют ткани. Спорофит преобладает в жизненном цикле (кроме мохообразных). Гаметофит постепенно редуцируется до нескольких клеток. Органы полового и бесполого размножения многоклеточные. Оплодотворение перестает зависеть от воды. Зигота дает начало многоклеточному зародышу. Основную часть биомассы на Земле (около 90 %) составляют наземные растения.
Общая характеристика высших споровых растений
Высшие растения, которые размножаются спорами, ботаники объединяют общим названием – высшие споровые растения. К ним относятся как современные, так и вымершие виды. Наука, которая изучает вымершие виды, называется палеоботаникой.
Высшие споровые ведут наземный образ жизни. Тело их делится на надземную и подземную части, развиты вегетативные органы (корень и побег).
Жизненный цикл состоит из чередования поколений: спорофита и гаметофита. Спорофит хорошо развит, живет продолжительное время (исключением являются мохообразные, где преобладает гаметофит, спорофит развивается на гаметофите и живет за его счет). На нем развиваются многоклеточные спорангии, в которых после мейоза образуются недвижимые гаплоидные споры. Они одноклеточные, маленькие по размерам. Распространяются преимущественно ветром. Споры прорастают в гаметофит. Споры могут быть одинаковыми по размерам или разными. Растения, которые образуют одинаковые споры, называются равноспоровыми, разные – разноспоровыми. Споры, которые имеют меньшие размеры, называются микроспорами (от греч. микрос – маленький), большие – мегаспорами (от греч. мегас – большой). Из микроспоры развивается мужской гаметофит, а из мегаспоры – женский.
Гаметофит меньше развит, чем спорофит, имеет меньшие размеры, живет непродолжительное время. На нем развиваются многоклеточные половые органы: мужские – антеридии (от греч. антерос – цветущий), женские – архегонии (от греч. архе – начало, гон – рождение). Антеридии имеют вид небольших шарообразных или овальных телец, внешняя стенка которых состоит из одного или нескольких слоев бесполых клеток. При созревании антеридии разрываются, и из них выходят сперматозоиды. Архегонии – это небольшие шарообразные тельца, которые имеют нижнюю расширенную часть и верхнюю суженную. Снаружи они тоже покрыты бесполыми клетками. В брюшной части находится яйцеклетка. Яйцеклетки неподвижные, остаются в архегониях. При их созревании клетки канальца суженной части архегониев ослизняются. Архегоний на верхушке открывается. Сперматозоид по слизи проходит в яйцеклетку. Половой процесс – оогамия. Оплодотворение происходит лишь при наличии воды, что дает возможность сперматозоидам попасть к яйцеклетке. По расположению половых органов различают двуполые (и мужские, и женские на одном растении) и однополые (на разных растениях) растения.
Считают, что семенные растения происходят от высших споровых растений.
Отдел риниофиты или риниеобразные
К отделу Риниофиты, или Риниеобразные (Rhyniophyta), относятся исключительно ископаемые растения. Жили приблизительно 450 млн. лет назад. Встречались на берегах водоемов, мелководье. Имели примитивное строение: тело ветвилось дихотомически (на две части). Настоящих листьев и корней не было. Имели ризоиды. Внутреннее строение более сложное, чем у водорослей: имели устьица, специализированную проводящую ткань. Небольшие по размерам (не выше 50 см). Не было механической ткани. Не найдены гаметофиты.
Представители: куксония (7 см в высоту), риния (50 см). Считается, что именно от риниофитов происходят все отделы современных растений.
Отдел псилофиты или псилотообразные
Представители отдела Псилофиты, или Псилотообразные (Psilophyta), сохранились до нашего времени (около 20 видов), они произрастают в тропическом и субтропическом климате. Ветвятся, как и риниофиты, дихотомически. Имеют внутреннее строение, подобное риниофитам. Листьев не имеют. Размеры не очень большие – до 1 м. Гаметофит нитевидный, мелкий (2 см), дихотомически разветвленный, развивается в почве или на коре деревьев.
Отдел мохообразные или бриофиты
Общая характеристика мохообразных
К отделу Мохообразные или Бриофиты (Bryophyta), относятся вечнозеленые, преимущественно многолетние растения. Появились приблизительно 400 млн. лет назад. Насчитывают около 23-27 тыс. видов. Встречаются на всех континентах, от пустынь до болот, особенно распространены в Северном полушарии. Предпочтение отдают хорошо увлажненным местам. Поселяются на коре деревьев, крышах и т. п. Встречаются виды, которые живут в воде. На протяжении лет могут сохранять жизнеспособность, находясь в высушенном состоянии. После дождя восстанавливают свою жизнедеятельность. Способны вбирать влагу из воздуха. Размеры колеблются от 1 мм до нескольких десятков сантиметров. Мхи изучает наука бриология (от греч. брион – мох и логос – учение).
В жизненном цикле наблюдается чередование поколений: спорофита и гаметофита. Особенностью мохообразных является преобладание гаметофита, тогда как у всех других высших растений преобладает спорофит. Поэтому эти растения рассматривают как самостоятельную боковую (тупиковую) ветвь эволюции. Спорофит (спорогон) развивается на теле гаметофита и существует за его счет. Состоит он из коробочки с ножкой. К гаметофиту крепится с помощью бесцветной клетки, погруженной в его ткани, которую называют стопа или гаустория (от лат. gaus thor– пьющий). Перед образованием спор происходит мейоз. Из споры развивается зеленая нить или пластинка, которая ветвится дихотомически – протонема (предросток).
Размножаются половым и бесполым способами. Вегетативно размножаются с помощью частей тела или образуют особые органы (выводковые корзинки, почки).
Мохообразные (гаметофиты) по строению тела делят на слоевищные и листостебельные. Слоевищные имеют вид листовидной пластинки, которая состоит из нескольких слоев клеток. Листостебельные разделены на стебель и листья (филоиды), спирально размещенные на нем. Настоящие корни отсутствуют. Большинство мохообразных имеют ризоиды. Не имеют настоящих сосудов. Слабо развита механическая ткань.
Многообразие мохообразных
К слоевищным мохообразным относят, например, маршанцию, к листостебельным, или настоящим мхам, – кукушкин лен, сфагнум.
Маршанция многообразная (Marchantia polymorpha)
Небольшое по размерам растение. Имеет слоевище, которое ветвится дихотомически, плотно прилегает к почве. Встречается в заболоченных, чрезмерно влажных местах. Прикрепляется ризоидами.
Размножается половым и бесполым способами. Для вегетативного размножения используются части таллома, или на поверхности растения могут образовываться выводковые корзинки с выводковыми почками, или по краю таллома – выводковые почки. Маршанция – двудомное растение. Половое размножение происходит при помощи сперматозоидов, которые формируются в антеридиях, расположенных на выростах-подставках в виде диска, и яйцеклеток – в архегониях на выростах-подставках звездчатой формы. Оплодотворение происходит при помощи воды. Из зиготы раз-вивается спорофит – спорогон. Он состоит из короткой ножки и коробочки. После созревания спор коробочка открывается. Для распространения спор она имеет специальные пружинные нити – элатеры (от греч. элатер – погонщик, кнут).
Кукушкин лен или рунянка, или политрих обыкновенный (Polytrichum commune)
Относится к зеленым мхам. Встречается на болотах, во влажных местах и т. п. Образует густые дернины. Может достигать 30 см в высоту. Имеет прямой, не разветвленный стебель, покрытый сидячими удлиненными ланцетовидными листьями. На верхней поверхности листьев есть особые пластинчатые фотосинтезирующие выросты, между которыми задерживается вода. Листья состоят из нескольких слоев клеток. В середине стебля есть удлиненные клетки, которые выполняют проводящую функцию. К почве прикрепляется с помощью многоклеточных ризоидов.
Кукушкин лен – двудомный: на верхушках одних растений развиваются архегонии, на верхушках других – антеридии. Антеридии похожи на мешочки на коротких ножках, в которых образуются сперматозоиды с двумя жгутиками. Архегонии собраны группами, колбообразной формы, имеют сдутое брюшко и удлиненную шейку. Листья, которые окружают антеридии, большие, красноватые или желтоватые. Листья, которые окружают архегонии, не отличаются от стебельных. Оплодотворение происходит при помощи воды. Женские половые органы выделяют наружу особые вещества, которые растворяются в воде и «указывают» дорогу сперматозоидам. Из зиготы прорастает спорофит – спорогон, состоящий из стопы, ножки и коробочки. Коробочка покрыта колпачком с тонкими, направленными вниз волосками, под которыми находится крышечка с зубчиками по краю. Эти зубчики способствуют распространению спор и называются перистомом. Колпачок снаружи похож на кукушку, что дало название мху. Перед созреванием спор происходит мейоз, и они становятся гаплоидными. После созревания в сухую погоду зубчики отгибаются наружу, споры высыпаются. Во влажную погоду – загибаются вглубь и препятствуют попаданию влаги и высыпанию.
Из споры развивается нитчатая протонема (предросток). На ней – почки, из которых образуются гаметофиты.
Сфагнум бурый (Sphagnum fuscum)
Относится к белым мхам. Насчитывает род свыше 300 видов. Распространен на болотах (доминируют среди других растений) и очень увлажненных местах от тропиков до субарктических областей. Стебель гаметофита сфагнумов достигает 50 см в высоту, очень разветвлен у верхушки. Ризоиды отсутствуют. Листья не имеют жилок, состоят из одного слоя клеток. Имеют два вида клеток: мертвые и живые. Специальные мертвые бесцветные клетки – водоносные – широкие, ромбовидные, способны накапливать много воды или воздуха, так как имеют специальные поры, посредством которых вбирают воду (в 20 – 30 раз больше массы мха). Эти клетки определяют цвет растения. Когда сфагнум отмирает, водоносные клетки заполняются воздухом (в сухом состоянии мох имеет беловатый цвет). Живые клетки – мелкие, узкие, имеют хлорофилл и потому зеленые. Растет сфагнум верхушкой, очень медленно (до 3 см в год), а нижняя часть постепенно отмирает.
Размножается сфагнум половым и бесполым способами. Вегетативное размножение происходит при помощи веточек. В спорангиях на спорофите (спорогоне), который развивается на гаметофите, состоит из нити, стопы и коробочки, развиваются споры. При вызревании спор крышечка с коробочки резко откидывается. Споры разлетаются при этом на расстояние до 10 см.
Спора прорастает в пластинчатую протонему с ризоидами, на которой развивается гаметофит. Ризоиды позднее исчезают. Гаметофит однодомный (у некоторых видов двудомный). На верхушках стеблей в пазухах листков расположены архегонии, на боковых веточках в пазухах листьев – антеридии. Оплодотворение происходит при помощи воды.
Значение мохообразных
Мхи играют важную роль в регулировании водного баланса территории и в процессе торфообразования. Способность к накоплению воды приводит к заболачиванию, которое может распространяться на большие площади. Основной составляющей торфа являются сфагновые мхи. Отмершие части сфагнумов погружаются в воду и оседают на дне болот на протяжении многих лет. Недостаток кислорода и способность мхов выделять кислоты и убивать микроорганизмы не дают перегнивать отмершим частям. Таким образом, образуется торф. Скорость отложения незначительна – приблизительно 1 см за 10 лет.
Торф – это ценное вещество, которое широко используется в хозяйстве. Его используют как топливо, удобрение, в строительстве, химической промышленности (изготовляют пластмассы, древесный спирт, парафин, нафталин, карболовую кислоту и др.). В медицине используют как бактерицидное вещество (сфагнол), против гниения.
Животными мхи почти не поедаются. Выступают в роли растений – пионеров, так как поселяются на песке, камнях и т. п.
Отдел плаунообразные или ликоподиофиты
Общая характеристика плаунообразных
Современные представители отдела Плаунообразные или Ликоподиофиты (Lycopodiophyta) –только травянистые, преимущественно многолетние вечнозеленые небольшие (до 30 см в высоту) растения, внешним видом напоминают мхи. Насчитывают около 1300 видов. Распространены в увлажненных местах суши. Преимущественно распространены в тропиках. Древнейшие среди современных высших споровых растений, у которых в жизненном цикле преобладает спорофит. Отмершие растения были деревьями и кустарниками, принимали участие в образовании каменного угля. Возникли в середине девонского периода палеозойской эры. Достигли расцвета в каменноугольный период. Происходят от риниофитов. Произрастали в полупогруженном состоянии в заболоченной почве.
В жизненном цикле чередуются спорофит и гаметофит. Спорофит преобладает. Он имеет ползучие (подземные или надземные) и прямостоячие побеги. Подземные части побегов имеют вид типичного корневища. Стебель ветвится вильчасто (дихотомически). Листья мелкие, представляют собой выросты стебля. Имеют одну жилку и устьица. Стебель снаружи покрыт однослойной эпидермой с устьицами. Под ней – кора. В центральной части стебля – проводящая ткань. Рост происходит в точке роста. Камбий в стебле отсутствует. Корень имеет подобное внутреннее строение. Для плаунообразных характерны дополнительные корни. На особых листьях (на верхней стороне или в пазухах) – спорофиллах (от греч. спора и филон – лист) – развиваются спорангии со спорами. Спорофиллы преимущественно расположены на верхушках побега. Они снаружи или не отличаются от обычных листьев, или отделены от вегетативной части стебля и образуют зону, подобную по строению шишке (имеют ось со спирально размещенными спорофиллами), которую называют стробилой (от греч. стробилос – шишка). Плаунообразные бывают равноспоровыми и разноспоровыми.
Гаметофит небольшой, имеет различную форму, называется заросток.
Размножаются половым и бесполым способами. Преобладает вегетативное размножение (побегами, выводковыми почками, клубеньками). В спорангиях происходит мейоз и получаются гаплоидные споры. Из споры развивается гаметофит. Половые органы представлены архегониями и антеридиями. В антеридиях развиваются двужгутиковые или многожгутиковые сперматозоиды. Оплодотворение происходит при наличии воды. Из зиготы вырастает спорофит.
Разнообразие плаунообразных
Представители равноспоровых плаунообразных: плаун булавовидный, плаун – баранец.
Представители разноспоровых: селягинелла и др.
Споры равноспоровых плаунов все одинаковые, не различаются. Гаметофиты (заростки) равноспоровых имеют маленькие размеры (2-20 мм), мясистые, напоминают формой луковицу. В процессе развития приобретают блюдцевидную форму. Они подземные или полуподземные, двуполые, сапрофиты или полусапрофиты (полуавтотрофы), то есть надземная часть имеет хлоропласты. Могут жить в симбиозе с грибами (микориза). Созревают от 12 до 20 лет. Равноспоровые плауны следует охранять.
Споры разноспоровых различаются: мелкие (микроспоры) дают начало мужским гаметофитам, большие по размерам (мегаспоры) – женским. Гаметофиты разноспоровых не зеленые, однополые. В особенности редуцированный мужской гаметофит. Они созревают в течение нескольких недель за счет питательных веществ спор. В процессе созревания не выходят за границы оболочки споры.
Значение плаунообразных
Плауны животные не едят. Некоторые из них содержат яд, сходный по действию на кураре. Споры богаты невысыхаемым маслом (до 50 %) и применяются как пересыпки литейных форм, при изготовлении ракет для фейерверков (треск возникает вследствие громкого щелкания оболочек при закипании масла). Растения используют как декоративные, лекарственные. В фармацевтике использовали раньше для обсыпания таблеток и как детскую присыпку. Некоторые из плаунов занесены в Красную книгу (баранец обыкновенный).
Отдел хвощеобразные или эквизетофиты
Общая характеристика хвощеобразных
Отдел Хвощеобразные (Equisetophyta) охватывает как современные, так и вымершие растения. Современные виды являются травянистыми, преимущественно многолетними растениями. Вымершие растения преимущественно были деревьями.
Достигли наибольшего развития в каменноугольном периоде. Представлены только одним родом Хвощ, который включает свыше 30 видов. Размеры спорофитов сильно варьируют (от 5 см до нескольких метров). Самые высокие растения встречаются в тропических лесах. Могут жить как во влажных, так и в сухих местностях. Распространены в разных климатических зонах, повсюду, кроме Австралии и Новой Зеландии.
В жизненном цикле происходит чередование поколений – полового и бесполого. Преобладает спорофит. Размножаются половым и бесполым способами. Вегетативно размножаются с помощью корневища.
Спорофит имеет особенность – расчленение тела на узлы и междоузлия. У него хорошо развито подземное корневище (тоже расчлененное) с дополнительными корнями и надземные побеги. В корневище запасаются питательные вещества и образуются в узлах на нем крахмалоносные клубни. В узлах – боковые веточки, расположенные кольцами (мутовками). От корневища отходят вертикальные надземные побеги. Листья не имеют хлорофилла, видоизменены в пленки, которые имеют форму зубчиков, срастаются у основания и образуют влагалище, которое прикрывает узел (защищает образовательную ткань). Фотосинтез осуществляется стеблем. Стебель пропитан кремнеземом, (двуокисью кремния), ребристый, имеет полость, заполненную воздухом или водой. Растут хвощи благодаря вставочной (интеркалярной) и верхушечной образовательной ткани (меристеме). Спорангии образуются на щитках (выростах оси) стробил, на верхушках вегетативных побегов или на специализированных спороносных побегах на нижней стороне спорофитов. После образования спор спороносные побеги отмирают. Имеют вид неразветвленных стеблей.
Хвощи – преимущественно равноспоровые. Из одинаковых спор преимущественно развиваются однополые гаметофиты. Споры имеют специальные приспособления – гигроскопические выросты – элатеры (от греч. элатер – кнут). С помощью элатер они сцепляются и распространяются группами. Это увеличивает возможность оплодотворения. Распространяются споры ветром.
Гаметофит однополый или реже – двуполый, имеет вид нити или зеленой расчлененной пластинки (от 1 до 30 мм в длину), существует непродолжительное время. На нем развиваются половые органы: архегонии и антеридии. Сперматозоиды преимущественно многожгутиковые. Оплодотворение происходит при помощи воды. Зародыш образуется из зиготы. До образования корня и надземных побегов питается за счет гаметофита.
Многообразие хвощей
В нашей стране встречаются хвощ луговой, хвощ лесной, хвощ болотный. Наиболее распространенным является хвощ полевой. Одним из крупнейших по размерам представителей хвощевидных является хвощ большой (до 1 м в высоту). Он занесен в Красную книгу.
Хвощ полевой (Equisetum arvense)
Многолетнее растение. Растет в условиях повышенной влажности на полах, лугах и т. п. Имеет два типа побегов: вегетативные и спороносные. Спороносные побеги развиваются весной. Они прямые, неветвистые (но расчлененные), незеленые (розово-бурые), до 20 см в высоту. На верхушках образуются спороносные колоски (стробилы): ось покрыта спорофиллами, которые имеют форму шестигранных щитков. На спорофиллах содержится 6 – 10 спорангиев. После созревания побеги отмирают.
Вегетативные побеги появляются весной, не гибнут летом, отмирают зимой. Они хорошо разветвлены, зеленые, без стробил, с хорошо развитыми междоузлиями. Листья имеют вид чешуек, маленькие. Они срастаются и образуют влагалище, которое одевает нижнюю часть междоузлия. Клетки эпидермиса пропитаны кремнеземом. Проводящая ткань представлена ситовидными элементами, разными типами трахеид, иногда сосудами.
Споры зеленоватые, шарообразной формы, оснащенные четырьмя элатерами. Заростки преимущественно однополые. Мужской заросток – маленькая зеленая расчлененная на лопасти пластинка, которая ризоидами прикрепляется к почве. На концах пластинки развиваются антеридии с многожгутиковыми сперматозоидами. Женский заросток имеет большие размеры. Из зиготы вырастает предросток, который превращается во взрослое растение.
Хвощ полевой – вредный сорняк на лугах и полях.
Значение хвощей
Практическое значение невелико. Молодые спороносные побеги и клубни хвоща полевого когда-то употребляли в пищу. Животными поедаются мало из-за кремнезема в стеблях. Встречаются ядовитые для скота виды. Хвощи из подрода гипохете могут употребляться в пищу оленями, кабанами, коровами, конями.
Стебли используют для шлифовки металла, дерева, чистки посуды. Могут быть вредными сорняками. Хвощи являются индикатором кислых почв (необходимо проведение известкования). В медицине используются (вегетативные побеги) как кровоостанавливающее и мочегонное средства. Вымершие хвощи принимали участие в образовании каменного угля.
Отдел папоротникообразные или полиподиофиты
Общая характеристика папоротникообразных
Первые представители отдела Папоротникообразные (Polypodiaphyta) появились около 400 млн. лет назад. Папоротникообразные – древние растения, значительная часть которых вымерла. В наше время они значительно превосходят количество видов других высших споровых растений. Известно свыше 12 000 видов. Современные папоротники – преимущественно травянистые растения, но в тропиках встречаются древовидные формы. Именно в тропических лесах папоротники достигают наибольшего видового многообразия и численности. Некоторые травянистые растения во влажных лесах поселяются на деревьях (эпифиты). Встречаются среди папоротникообразных лианы. Папоротники могут образовывать сплошные заросли. Размеры колеблются от нескольких миллиметров до нескольких метров. Растут во влажных местах или в воде.
В жизненном цикле наблюдается чередование бесполого и полового поколений. Преобладает спорофит. Спорофит имеет корневище и хорошо развитые листья. От корневища отходят придаточные корни. Он имеет эпидермис, механическую и проводящую (сосудистые пучки) ткани. Гаметофит очень чувствителен к недостатку влаги.
Размножение половое и бесполое. Вегетативное размножение происходит при помощи корневища, выводковых почек. Различают равноспоровые и разноспоровые папоротники. Почти все папоротники, за исключением водных, являются равноспоровыми растениями.
Разнообразие папоротникообразных
Равноспоровые папоротники
Это преимущественно многолетние растения. Спорофит имеет подземное или надземное хорошо развитое корневище с придаточными корнями. Надземная часть – листья стебельного происхождения с цельной или расчлененной (перистой или многократно рассеченной) листовой пластинкой. Они называются вайи (от греч. байон – пальмовая ветвь). В молодом возрасте листья папоротника закручены улиткообразно и растут, как побег, верхушкой. Листья развиваются медленно. На листьях размещаются спорангии. У некоторых видов бывает два типа вай – фотосинтезирующие (вегетативные) и спороносные (спорофиллы). Спорангии расположены на нижней поверхности вай и собраны в сорусы (от греч. сорус – кучка). Сверху сорусы прикрыты покрывальцем или индузием (от лат. индузиум – древнеримская одежда – верхняя туника). Перед вызреванием спор происходит мейоз. Споры дают начало двуполому гаметофиту – заростку.
Гаметофит маленький, зеленый, у разных видов разной формы, преимущественно питается самостоятельно. На нем развиваются половые органы: архегонии и антеридии. Яйцеклетки и сперматозоиды одного заростка созревают в разное время (чтобы предупредить самооплодотворение). Сперматозоиды преимущественно многожгутиковые. Оплодотворение возможно при наличии воды. Из зиготы формируется предросток (спорофит), который имеет гаусторию (ножку, погруженную в ткани гаметофата), первичный корешок, стебелек и листья. Первые дни он питается за счет гаметофита, пока не начнет самостоятельно синтезировать. Зародышевый корешок позднее редуцируется – его заменяют придаточные корни. Гаметофит отмирает.
Представителями равноспоровых папоротников являются, например, орляк, щитник мужской, безщитник женский и др.
Щитник мужской (Dryopteris filix-max)
Является типичным представителем равноспоровых папоротников. Его спорофит образует толстое подземное ползучее корневище. На нем ежегодно появляется розетка листьев. Листья большие, простые, двуперисторассеченные. На нижней стороне вай развиваются округлые сорусы. Когда споры выстывают, покрывальце становится красноватого цвета, кончики его заворачиваются и споры высыпаются. Из споры прорастает гаметофит.
Гаметофит (заросток) имеет вид небольшой (0,5-0,9 см в диаметре) сердцевидной зеленой пластинки, которая ризоидами прикрепляется к почве. На нижней стороне заростка образуются архегонии и антеридии. Сперматозоиды спирально закручены, многожгутиковые. Из зиготы развивается спорофит.
Разноспоровые папоротники
Это водные растения. Мужские и женские гаметофиты развиваются из микро- и мегаспор, которые развиваются в микро- и мегаспорангиях. Мужской гаметофит очень редуцированный. Женский – многоклеточный, развивается в середине мегаспоры. Половой орган женского гаметофита покрывается слизью, которая имеет вещества, привлекающие сперматозоиды. После оплодотворения развивается многолетний спорофит. Прорастание спор, оплодотворение и развитие спорофита происходит в воде.
Представители разноспоровых: сальвиния плавающая, марсилия четырехлистная и др.
Сальвиния плавающая (Salvinia natans)
Встречается в стоячих или с медленным течением водоемах на поверхности. Сохранилась как вид с древних времен – является реликтовым (от лат. relictum – остаток) растением. Корневище без корней. На нем развивается несколько мутовок из 3 листьев, два из которых – на поверхности воды, а третий, рассеченный на много тонких корневидных частей, покрытых волосками, – под водой. На этих частях с течением времени развиваются микро- и мегасорусы. Сальви-ния плавающая редко встречается и требует охраны.
Марсилия четырехлистная (Marsilea quadrifolia)
Встречается в плавнях Днестра и Карпатах. Растет на берегах водоемов и мелководьях. Имеет ползучее корневище, от которого отходят вертикально листья, которые имеют длинный черешок и 4 листика. Споры развиваются в сорусах, которые закрыты во вместилищах, которые раскрываются после смачивания. Редчайшее растение, требует охраны.
Значение папоротников
Принимали участие в процессе углеобразования. Отмершие папоротники (преимущественно деревья) вместе с другими споровыми растениями (хвощами, плаунами), занесенные глиной, песком и т. д., при незначительном доступе кислорода почти не разлагались. Отсутствие воздуха и высокое давление слоев Земли превращали остатки растений в каменный уголь. Каменный уголь – это источник энергии. Он используется в промышленности, хозяйстве.
Папоротники образуют растительные группы. Молодые листья некоторых травянистых папоротников, а также сердцевину древовидных употребляют в пищу. Папоротники используют как декоративные растения (адиантум, нефролепис).
В медицине используются как глистогонное средство, для лечения открытых ран, при лечении болезней горла и кашля.
Некоторые папоротники занесены в Красную книгу: вудсия альпийская, адиантум Венерин волос, сальвиния плавающая, марсилия четырехлистная и др.
Источник: xn—-9sbecybtxb6o.xn--p1ai
Споровые растения: первое знакомство
Начнем знакомство с небольшого описания. Слово «спора» пришло к нам из греческого языка. В переводе оно означает «семя» или «семечка». Речь идет об очень маленьком образовании, размер которого — примерно 1 мкм.
Сформировались споровые растения очень давно. Фактически они — прямые потомки той флоры, которая попала из океана на сушу. К споровым растениям относится не только папоротник. Ученые подразделяют их на две категории: высшие и низшие. В первой категории – папоротники, плауны, мох и хвощи. Во второй – водоросли и лишайники.
Жизненный цикл споровых растений
Если говорить о высших споровых организмах, то у них очень интересный жизненный цикл. Здесь можно увидеть чередование особей бесполого и полового видов. Соответственно, и размножение, в зависимости от вида, идет половым или бесполым путем. Полный жизненный цикл непрерывен. Растение образовывает гаметофит (орган полового размножения) и спорофит (орган бесполого размножения).
Эволюция позволила развиваться этим видам растений в двух направлениях. Получились две обширные группы: гаплоидная и диплоидная. Описывая споровые растения, примеры гаплоидной группы которых – мхи, ученые доказали, что у них более развит половой гаметофит. Спорофит у гаплоидной группы имеет подчиненный статус. Диплоидное направление споровых организмов (хвощ и папоротники) имеет сильно развитый спорофит, а гаметофит в виде заростка.
Половое поколение всегда имеет антеридии и архегонии. Это мужские и женские органы. Мужские сперматозоиды подвижны, женская половая клетка статична. Чтобы ее оплодотворить, сперматозоид должен попасть в водную среду, по которой сможет добраться до цели. Оплодотворенная яйцеклетка образует зародыш, из которого прорастает неполовое поколение, то есть спорофит. Следующий этап размножения произойдет спорами, которые развиваются в спорангиях.
Отличительные признаки
Не все знают, как отличить споровые растения. Примеры рассуждений на эту тему могут выглядеть так:
- Споровые растения не цветут. Этот вид биологически не способен к цветению.
- Имеют уникальный жизненный цикл. Половое и бесполое размножение.
- Невозможность полового оплодотворения без присутствия воды.
Если три признака присущи рассматриваемому растению, то это споровый вид.
Споровые растения: папоротники
Трудно найти человека, который никогда не видел папоротник. Это древнее растение используют для украшения парков и садовых участков. Комнатные виды папоротников выращивают в вазонах, а любители лесных прогулок много раз видели пышные и зеленые папоротниковые заросли.
У всех папоротников преобладают перисто-рассеченные сложные листья. Эти споровые растения (папоротники) имеют самые заметные спорангии. Место расположения этих органов – нижняя сторона листьев.
Для информации добавим, что папоротников в природе больше десяти тысяч. Все это разнообразие объединено в 300 родов.
Строение спорового растения на примере мхов
Мхи можно отнести к наиболее примитивным видам высших растений. Все моховидные – это небольшие по размеру представители без проводящей ткани. Подразделение на стебель и листья у мхов условное. Эти удивительные споровые растения – примеры умения приспосабливаться к условиям природы.
Итак, тело мха условно разделено на стебель, листья и корни. Да, корни этому растению заменяют нитевидные выросты – ризоиды. Их основное отличие от настоящих корней – отсутствие проводящей ткани. У мха каждый ризоид – одна или несколько живых клеток.
Мхи хорошо чувствуют себя в болотистых местах, в тени или просто в сырости. Мхи активно испаряют влагу, но восполняют потерю всей поверхностью растения. Несмотря на то, что споровым растениям необходима вода, чтобы размножаться, некоторые виды мхов способны пережидать периоды засухи, они даже приспособились выживать на скальных участках. Как это происходит, до конца не понятно.
Доминирующее поколение мхов – половое. Спорофит полностью зависим от гаметофита.
Споры не могут образовываться на листьях, как у папоротника, поскольку сами листья весьма условны. Для этих целей у мхов приспособлена споровая коробочка, возвышающаяся над половым гаметофитом на ножке-ниточке.
Особенностью мхов считается возможность вегетативного размножения. В этом процессе задействованы почки и клубеньки. Если вегетативную часть отделить от основного растения, то она развивается в самостоятельную особь.
Немного о низших видах
Не будем перечислять все низшие споровые растения. Примеры, которые интересно описать, – водоросли. Эти растения не так многочисленны, как папоротники и лишайники, их чуть больше ста видов. Среда обитания этой флоры – вода. Водоросли не имеют листьев и корней. Крепятся к грунту или камням прозрачными крючками. Подразделяют водоросли на 11 отделов, 4 из которых человек научился использовать в своих целях.
Источник: FB.ru
Споры в том или ином виде образуют все растения. Процесс их образования называется спорогенезом, а клетка или орган, в котором они формируются, носит название спорангия. При этом особь поколения, образующего споры, называют спорофитом . Как правило, спорофит имеет диплоидное число хромосом и только у водорослей и некоторых грибов споры могут образовываться и гаплоидными особями.
Споры — одноклеточные образования, из которых без слияния их друг с другом могут развиваться самостоятельные организмы. Очень часто споры окружены специальной оболочкой и способны длительно переносить неблагоприятные условия, например зиму или период засухи. Поскольку споры очень легки, они свободно переносятся по воздуху ветром или другими агентами и служат для расселения растений. Споры водных растений обычно имеют ундулиподии и способны активно перемещаться.
Споры могут образовываться без редукционного деления на спорофите , реже на гаметофите и тогда имеют то же число хромосом, что и материнское растение. Либо, претерпевая спорическую редукцию, споры образуются путем мейоза . В этом случае на диплоидном спорофите образуются гаплоидные споры.
Образование спор без редукционного деления на спорофите или гаметофите встречается у примитивных организмов довольно часто. Для целого ряда грибов характерно бесполое размножение спорами, образующимися на гаплоидном мицелии. Такие споры развиваются либо в спорангиях ( спорангиоспоры ), либо, называясь конидиями , отпочковываются верхушкой приподнимающейся специализированной гифы, так называемым конидиеносцем. Нередко гаметофиты бурых, а иногда и других водорослей образуют гаплоидные зооспоры , развивающиеся в аналогичные гаметофиты. Гаметофиты , образующие споры, в таких случаях называют гаметоспорофитами . Их спорангии представляют обычно одну клетку, в которой развивается один зооспорангий . В определенных условиях зооспоры , образованные гаметоспорофитами , могут попарно сливаться и образовывать зиготу, представляя изогамный половой процесс.
Развиваются диплоидные споры у низших растений либо в клетках, не отличающихся по форме от других клеток организма, либо в спорангиях, которые более или менее сильно отличаются от вегетативных клеток по форме и размеру. Спорангии обычно развиваются из одной клетки, однако у бурых водорослей они изредка образуются и несколькими клетками. Это так называемые многогнездные спорангии. Все их клетки при созревании превращаются в споры.
У большинства водорослей при образовании спор на диплоидном слоевище происходит редукция числа хромосом вследствие мейоза. Такие гаплоидные споры дают начало гаметофиту , на котором образуются половые органы. Очень редко, например у некоторых бурых водорослей , гаплоидные споры, образуемые спорофитом , функционируют как гаметы. Гаплоидные, как и диплоидные, споры образуются в одноклеточных спорангиях и обычно снабжены ундулиподиями . Как и подвижные жгутиковые диплоидные споры, они называются зооспорами .
Гаплоидные споры, образовавшиеся на спорофите и дающие начало гаметофиту , могут быть одинаковыми, или диморфными. Гаметофиты равноспоровых организмов обычно развивают и мужские и женские половые органы на одном растении. Гаметофиты разноспоровых организмов, развиваемые микроспорами, образуют, как правило, только мужские гаметы, а гаметофит, развивающийся из мегаспоры, образует женские половые клетки — яйцеклетки.
У низших растений спорофит и гаметофит в жизненном цикле сосуществуют в разной степени доминирования, а у высших растений , за исключением мхов , гаметофит все более и более редуцируется.
Споры. Фото: Kristine Paulus
Совокупность оболочек споры, спородерма, имеет сложное строение, дифференцируясь на наружный толстый слой, экзину, и внутренний — интину. Экзина состоит из спорополленина, одного из самых стойких органических веществ, способного выдерживать длительные температурные и химические воздействия. Благодаря стойкости экзины, споры обычно длительное время сохраняют способность к прорастанию и могут сохраняться в отложениях на протяжении геологических эпох. Спородерма часто имеет скульптурированные оболочки, покрытые гребешками, бугорками, шипами и т. д.
К споровым относятся растения, размножение и распространение которых осуществляется посредством спор, причем споры образуются двумя путями – бесполым и половым. Споровыми растениями являются водоросли, грибы, лишайники и высшие споровые растения (папоротники, плауны, хвощи, мхи, некоторые ископаемые растения).
В процессе эволюции около 400 млн. лет назад от многоклеточных зеленых водорослей возникли риниофиты – первые высшие растения, размножающиеся спорами, которые дали начало всем современным высшим споровым и семенным растениям. Это вымершая группа растений.
В жизненном цикле высших споровых растений, как и у некоторых водорослей, чередуются особи бесполового и полового поколений, которые размножаются, соответственно, бесполым и половым путем. В полном жизненном цикле, который обеспечивает непрерывность жизни организмов, происходит чередование гаметофита (полового) и спорофита (бесполого поколения). На спорофите образуются органы бесполого размножения, на гаметофите – полового.
Высшие споровые растения после выхода на сушу в ходе эволюции претерпевали метаморфозы в двух направлениях. Так сформировались две крупные эволюционные группы – гаплоидная и диплоидная. К первой ветви относятся мхи, у которых лучше развит гаметофит, а спорофит занимает подчиненное положение. К диплоидной ветви принадлежат папоротники, хвощи и плауны. У них гаметофит редуцирован, и выглядит как заросток.
Из спор, которые формируют особи бесполого поколения, вырастают особи полового поколения. Они имеют специальные мужские и женские половые органы, в которых развиваются мужские и женские половые клетки (гаметы) – подвижные сперматозоиды и неподвижные яйцеклетки. Для оплодотворения сперматозоид должен попасть во внешнюю среду и оплодотворить яйцеклетку, которая находится внутри женского полового органа. Для перемещения сперматозоида необходима вода. Из оплодотворенной яйцеклетки образуется зародыш. Он прорастает и превращается в особь бесполого поколения, которая размножается спорами.
Грибы, лишайники и водоросли размножаются подвижными зооспорами, спорангиеспорами, неподвижными апланоспорами. Для этих растений характерно половое размножение ооспорами, зигоспорами, вегетативное размножение также посредством спор. Из спор появляется такое же растение, как материнское. Тогда как у высших споровых растений такое растение, как то, что сформировало споры, появляется после сложного процесса оплодотворения.
Споры у определенного вида растений могут быть одинаковыми по размерам или разными. Микроспоры – мелкие, макроспоры – крупные. Микроспоры обычно дают начало мужским заросткам, а макроспоры – женским.
Высшие споровые растения распространены в разных климатических условиях, но большинство из них обитает на влажных участках суши, так как для полового размножения им нужна вода. Однако некоторые виды этих растений встречаются и в пустынях.
Сотрудники университета Ноттингема установили, что споры древних растений содержат в себе информацию об «озоновых дырах».
Дело в том, что растения крайне чувствительны к ультрафиолету, поэтому по ним можно буквально изучать изменения состояния озонового слоя Земли.
Судить о силе поступающего на землю ультрафиолетового излучения учёные смогли благодаря наблюдениям за толщиной образуемого на стенках спор растений в различные периоды защитных веществ. А, как известно, количество поступающего на нашу планету ультрафиолета напрямую связано с состоянием озонового слоя.
Исследователи утверждают, что теперь стало возможным получить обширные сведения о защитном слое Земли за период равный миллионам лет. Кроме того, специалисты рассчитывают определить, насколько изменения в озоновом слое соответствуют естественным историческим колебаниям.
Споры растений, как, оказалось, могут оказать людям непосильную помощь при сборе нефти из поверхности воды.
По убеждению ученых, если модифицировать микрокапсулы спор растения, то они могут абсорбировать количество разлившейся нефти, которое превышает собственный вес спор более чем в три раза.
Команда ученых из Университета Хала во главе с Грэхэмом Маккензи смогли экстрагировать внешний слой спор плауна и модифицировать его до состояния пустых модифицированных микрокапсул растения. Данные микрокапсулы ученые поместили в воду с нефтью, после этого емкость встряхивали в течение пятнадцати минут, после микрокапсулы были отфильтрованы. Как результат, после фильтрации вода была абсолютно чистой, и не содержала нефтепродуктов.
Как говорят исследователи, микрокапсулы могут быть использованы несколько раз подряд без утраты эффективности их нефтеочистительных свойств. Кроме этого, возможен и обратный процесс — нефть может быть выпущена из микрокапсул. Из этого следует, что микрокапсулы, могут пригодится в косметической и фармацевтической отрасли.
Источник: biofile.ru
Автор статьи — Л.В. Окольнова.
Это растения, которые первыми освоили сушу. Их относят в высшим, т.к. по сравнению с водорослями, у них уже появились ткани и органы — приспособления для жизни в наземной среде.
Водоросли тоже размножаются спорами, но это водные организмы, самые простые по организации.
Основная особенность споровых — размножение специальными клетками — спорами.
Прежде, чем мы разберем, что из себя представляют споры, давайте вспомним, какие бывают клетки.
Клетки:
- соматические (или вегетативные) — клетки тела организма. У них двойной (диплоидный) набор хромосом, обозначается как 2n
- половые ( или генеративные, или гаметы) — это “половинчатые” клетки — их набор хромосом в 2 раза меньше, чем у соматических — 1n
Получается, что споры — это такие половые клетки ( или группы клеток) у растений. Растение, которое их образует — спорофит — оно диплоидное (2n)
НО:
У животных гаметы образуются путем мейоза, у растений — путем митоза.
У животных гаметы тут же сливаются, происходит оплодотворение, новый организм получает двойной — диплоидный (2n) набор хромосом.
У споровых растений клетка споры прорастает в целый самостоятельный организм — гаметофит, который может опять производить половые клетки — гаметы.
Гаметы сливаются и образуют опять спорофит.
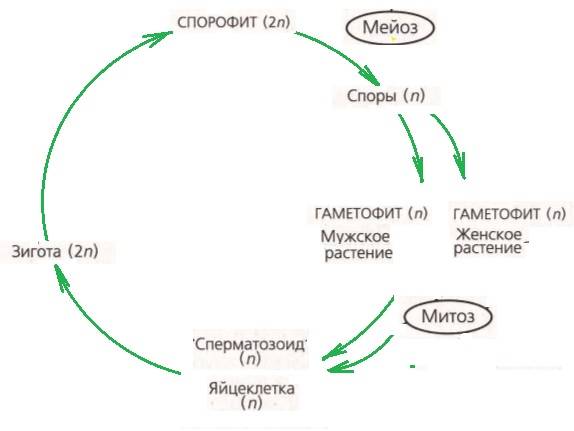
Получается, что жизнь споровых растений — своеобразный цикл — чередование спорофита и гаметофита.
У растений может преобладать какая-то фаза:
В целом, у всех споровых растений жизненный цикл выглядит следующим образом:
Зачем споровым растениям понадобились такие сложности, такая многостадийность в жизненном цикле?
Это были первые растения, освоившие молодую, далеко не комфортную сушу (мхи появились примерно 350 млн лет назад!)
Условия жизни были довольно сложными, поэтому каждая стадия старалась закрепиться, накопить побольше питательных веществ и уже потом развиваться дальше
Источник: ege-study.ru