Способы размножения патогенных микроорганизмов

Микроорганизмы – довольно быстрые живые существа по своей реакции на изменяющиеся условия окружающей среды и жизненному циклу. Так, при идеальных благоприятных условиях рост и размножение микроорганизмов происходит чрезвычайно быстро. Большинство бактерий делятся каждые полчаса, но этот процесс не постоянный, так как зависит от изменяющейся окружающей среды и внутренних процессов самих микроорганизмов. В идеальных же условиях бактерии могли размножаться бесконечно долго до исчерпания всех полезных веществ на планете.
Зачастую размножение микроорганизмов происходит в рамках одних закономерностей. После того, как микроорганизмы прошли стадию засева питательной среды, они будто бы замирают, находятся в состоянии покоя. После данной стадии, на которой они вообще не развиваются, наступает стремительное и быстрое их размножение. После достижения микроорганизмами максимально возможной концентрации в данной питательной среде происходит такой же быстрый процесс их отмирания. В конце цикла микроорганизм полностью аннигилирует питательную среду и вынуждено умирает до последней клетки.
Также полное отмирание микроорганизма происходит, если благоприятная среда становится неблагоприятной. К этим факторам относятся показатели влаги, которые должны быть на достаточно высоком уровне, а также постоянная температура. Скачки температуры, резкое охлаждение могут заставить микроорганизм перейти в замирание, анабиоз или же полностью умереть. Нередки те случаи, когда после быстрого замораживания и правильного и последовательного размораживания микроорганизм продолжал свою жизнедеятельность и процесс размножения.
Существуют различные способы размножения микроорганизмов. Выделяют следующие способы: простое, множественное деление, почкование, спорообразование. Также это может быть половой способ и параполовой. Основным способом является простое деление. Простое или бинарное деление предполагает образование из одной клетки микроорганизма двух новых клеток. Эти дочерние клетки являются равноценными, и между ними не происходит обмена генетической информацией.
Бинарное деление включает фазу роста клетки, фазу ее удвоения, фазу разделения, а также расхождения дочерних клеток. Дочерние клетки по достижению соответствующего размера также могут делиться, разрастаясь таким образом. Дочерние клетки также могут не раздваиваться полностью, образуя между собой цепочки, пучки или другие образования нескольких клеток.
Размножение и рост микроорганизмов находятся под контролем генома, эти процессы являются сбалансированными и последовательными. Если же внешняя среда изменяется или происходит какое-либо влияние на микроорганизм, на его генном уровне могут произойти определенные изменения, которые приводят к неконтролируемому росту и замиранию размножения. То есть, клетка начинает сильно расти, но размножение не наступает, что приводит к образованию инволюционной формы микроорганизма.
Источник: mikrobiki.ru
Среди микроорганизмов, ставших объектами генетических исследований, известны многие грибы, водоросли, бактерии и вирусы.
Грибы и большинство водорослей имеют обычную организацию ядра, свойственную растительной клетке, хотя у разных представителей этих организмов детали строения ядра могут различаться. Для бактерий характерно особое строение ядра: в нем имеется, как правило, одна очень маленькая хромосома, лишенная видимых под световым микроскопом морфологических особенностей; под электронным микроскопом видна связь этой хромосомы с клеточной мембраной. Ядро бактерий не отделено от толщи цитоплазмы мембраной. При делении ядро не претерпевает реорганизации, подобной той, которая происходит во время митотического цикла в клетках высших организмов, и веретено деления не образуется.
Организмы, имеющие ядро, сходное с бактериальным, называют прокариотами в отличие от высших организмов — эукариотов.
ро бактерий и актиномицетов содержит ДНК в виде нитей диаметром 25—30 Å. Такое ядро называют нуклеоидом. Оно окрашивается по Фельгену. Число нуклеоидов в бактериальной клетке различно: в одной клетке их может быть один или несколько, причем разной формы. Дочерние нуклеоиды воспроизводятся прямым делением. С помощью радиоактивной метки установлено, что репликация ДНК в нуклеоиде осуществляется полуконсервативным путем. Генетически нуклеоид соответствует хромосоме (геному), что дает нам известное право в дальнейшем называть геном бактериальной клетки хромосомой.
Размножение неспороносных бактерий состоит из повторяющихся делений вегетативных клеток (Salmonella, Escherichia, Schigella и др.). Установлено, что в процессе размножения культура микроорганизма проходит пять фаз:
- лаг-фазу
- фазу ускорения
- логарифмическую (экспоненциальную)
- фазу замедления
- стационарную
Размножение клеток, помещенных в свежую питательную среду, на некоторое время задерживается. Такое состояние культуры называют задержкой размножения (лаг-фазой). В следующей фазе происходит ускорение роста, и частота делений клеток за единицу времени постепенно нарастает (фаза ускорения). Затем следует фаза, когда скорость размножения клеток за единицу времени постоянна (фаза логарифмического роста). Истощение питательной среды вызывает фазу торможения скорости деления, и затем наступает так называемая стационарная фаза, когда число жизнеспособных бактерий в культуре не меняется. Следующая, последняя, фаза связана с отмиранием клеток.
Все эти фазы проходит одна культура без дополнения питательной среды. При смене фаз меняются численность клеток в культуре, морфология клеток и число нуклеоидов в клетке.
Типичный половой процесс обнаружен не у всех микроорганизмов. Однако многие грибы и водоросли имеют половой процесс, который в принципе сходен с таковым у высших растений. У этих организмов гаплоидные продукты мейоза некоторое время сохраняются вместе, что дает возможность применять тетрадный анализ и непосредственно изучать результаты расщепления в мейозе. Гаплоидное состояние клеток позволяет легко устанавливать частоту и характер возникающих рецессивных мутаций, поскольку отсутствие соответствующих доминантных аллелей исключает необходимость перевода этих мутаций в гомозиготное состояние.
У некоторых грибов, наряду с нормальным половым процессом, имеется парасексуальный (дословно — возле полового) цикл, который также обеспечивает рекомбинацию наследственных факторов.
Термин «парасексуальный» был предложен для обозначения процесса рекомбинации наследственных факторов, осуществляющегося в митозе и не связанного с оплодотворением и мейозом. Например, у плесневого гриба аспергилла его можно выявить следующим образом. Гифы мицелия многоядерны; большинство ядер находится в гаплоидном состоянии. При совместном выращивании двух разных мутантных мицелиев между их гифами возникают цитоплазматические анастомозы, через которые происходит обмен ядрами. В результате такого обмена образуется гетерокарион, % е. мицелий с гаплоидными ядрами разных генотипов.
При образовании одноядерных конидий гетерокарион распадается на два исходных мутантных генома. Редко, но с определенной частотой при вегетативном росте гетерокариона может происходить слияние двух гаплоидных ядер с мутантными геномами и образование диплоидного гетерозиготного ядра. Это явление называют диплоидизацией.
Клетка с диплоидным ядром путем ряда делений дает диплоидные гетерозиготные гифы. В процессе деления диплоидных клеток независимо друг от друга могут протекать еще два различных процесса: во-первых, редкий митотический кроссинговер и, во-вторых, случайная (нерегулируемая), не связанная с мейозом, гаплоидизация ядер. И тот, и другой процессы приводят к расщеплению в потомстве гетерозиготного диплоида. При этом гаплоидные и диплоидные участки мицелия можно различать как по размеру конидий — у первых мельче, чем у вторых, так и генетически — у гаплоидных форм выявляются рецессивные гены всех групп сцепления, а у диплоидных форм эти гены проявляются лишь в участках хромосом, претерпевших митотический перекрест, т. е. там, где они оказываются в гомозиготном состоянии.
При гаплоидизации в ходе парасексуального цикла отсутствует синапсис хромосом, и поэтому гены каждой из хромосом обнаруживают полное сцепление. Гены негомологичных хромосом перекомбинируются независимо.
Следовательно, гаплоидизация в парасексуальном цикле облегчает разбивку генов по группам сцепления. Механизм гаплоидизации пока точно не выяснен. Ясно лишь, что при этом происходит постепенное уменьшение числа хромосом в ряду митотических делений вплоть до их гаплоидного числа.
Определение местоположения гена, т. е. его локализации в пределах группы сцепления, в случае парасексуального процесса производят на диплоидной стадии на основе митотического кроссинговера. И хотя митотический кроссинговер осуществляется лишь при случайной и очень редкой конъюгации гомологичных хромосом у гетерозиготного диплоида, тем не менее его можно использовать для локализации. Очевидно, частота выхода гена в гомозиготное состояние характеризует его расстояние от центромеры. На этом принципе были построены карты хромосом аспергилла и других грибов.
Если у аспергилла парасексуальный процесс существует наряду с обычным половым процессом, то для ряда других грибов он является единственным способом рекомбинации генов. К таким грибам относится, например, пеницилл. Обнаружение у него парасексуального цикла позволило использовать процесс рекомбинации в селекции этого важнейшего продуцента антибиотиков. При изучении парасексуального процесса удалось также провести генетический анализ свойства вырабатывать антибиотик.
Гибридизация у микроорганизмов может осуществляться несколькими путями:
- копуляцией
- конъюгацией
- трансдукцией
- трансформацией
Копуляция известна у грибов и водорослей. При копуляции происходит слияние гамет и образование зиготы, совмещающей в себе ядро и цитоплазму обеих гамет.
Гибридизация путем конъюгации, трансдукции и трансформации характерна для бактерий. При данных способах гибридизации наследственные факторы передаются односторонне — от донора к реципиенту и только частично в виде отдельных фрагментов ДНК. Клетка-реципиент, получающая лишь часть наследственной информации от клетки-донора и возникшая в результате любого из этих трех типов гибридизации, называется мерозиготой.
Вирусы размножаются и гибридизируются иным путем, отличным от всех других организмов: они размножаются только внутри клеток.
Вирусы являются паразитами клеток животных, растений и бактерий. Вирусы, паразитирующие в бактериях, называют фагами (бактериофагами). Фаги поражают определенный для каждого из них вид и даже определенный штамм бактерий. Они имеют характерную форму и размеры, отличающие их от других вирусов. Например, фаги серии Т (Т1, Т2, Т4 и т. д.) кишечной палочки по форме на поминают сперматозоиды млекопитающих и состоят из головки и хвоста. Размер частиц фагов колеблется от 200 до 500 ммк, размер головки от 45 до 80 ммк, длина хвоста от 100 до 170 ммк.
Фаг состоит из белковой оболочки и внутреннего содержимого – ДНК. Нуклеиновая кислота находится в головной части фага. Конец хвоста фага морфологически довольно сложен. Им фаг прикрепляется к поверхности поражаемой клетки. На кончике хвоста фага имеется фермент лизоцим. Вслед за прикреплением к бактериальной клетке, фаг локально разрушает (лизирует) оболочку, и содержащаяся в нем ДНК впрыскивается внутрь клетки. Так осуществляется заражение клетки. При этом белковая оболочка фага остается на поверхности клетки, а внутрь ее проникает, по-видимому, лишь ДНК фага.
Внутри бактерии ДНК фага начинает воспроизводиться, реплицироваться, используя ферментные системы и материалы клетки-хозяина. В первый период репродукции фага внутри бактерии фаговый белок не обнаруживается. Он выявляется позднее, когда уже накапливается в достаточном количестве ДНК фага. Затем частицы ДНК фага окружаются специфическим белком, в результате чего образуются зрелые частицы фага. Одна частица фага, попавшая в клетку, способна дать начало сотне и более новых частиц. Через 10—45 мин после заражения бактериальная клетка лизируется, и из нее в среду выходят зрелые частицы фага (от 100 до 300 частиц), которые способны вновь заражать здоровые бактерии.
Размножение фага в жидкой бактериальной культуре выявляется просветлением суспензии из-за лизиса клеток. На агаровой среде размножение фагов выявляется в виде пятна лизиса на сплошном газоне культуры в чашках Петри. Эти зоны лизиса называются негативными колониями фага, так как каждая такая колония является результатом размножения одной исходной фаговой частицы.
Фаг, способный вызвать лизис зараженной бактерии, называют вирулентным. Наряду с вирулентными фагами существуют так называемые, умеренные. Последние, проникая в клетку, не разрушают ее, а остаются в ней в качестве «жильцов» — симбионтов. При этом умеренный фаг пребывает в ней в форме профага, который размножается синхронно с делением самой клетки. Профаг, таким образом, представляет собой невирулентное состояние фага.
Присутствие профага в бактерии делает ее иммунной по отношению к заражению частицами генетически родственного фага. К частицам фага другой генетической природы бактерия не имеет иммунитета, и она может быть ими заражена. Поэтому в клецку, могут проникать частицы фагов разной генетической природы.
Нормально размножающаяся бактерия с находящимся в ней профагом называется лизогенной бактерией. Явление симбиотических отношений фага с бактерией называют лизогенией. Отношение бактериальных клеток к фагам, лизирование бактерии, или лизогенизация ее, определяются генотипом бактерии и фага. Вопрос о том, каким образом воспроизводится наследственная основа фага в лизогенных бактериях, до сих пор остается загадкой. Но очевидно, что профаг тесно связан с ядерным веществом бактерий.
Состояние лизогении не абсолютно устойчиво. Иногда происходит нарушение лизогенного состояния бактерий (индукция фага), при этом начинается синтез фаговых частиц, не контролируемый клетками, и клетки разрушаются. Затем из бактерии выходят зрелые частицы фага, способные заражать другие бактерии Лак же, как и в случае вирулентности фага. Индукция синтеза фаговых частиц в бактерии и выход фага из нее вызываются рядом внешних агентов, многие из которых являются мутагенными.
Фаги, как правило, имеют двунитчатую спиральную структуру ДНК и весьма разнообразную по химическому составу белковых Молекул оболочку. Лишь некоторые фаги (фХ-174) содержат однонитчатую структуру ДНК, однако в момент репликации у них молекула ДНК двунитчатая.
Среди животных вирусов известны такие, которые имеют ДНК, и такие, которые имеют РНК (например, вирусы, вызывающие заболевание полиомиелитом).
Наиболее изученным растительным вирусом является вирус табачной мозаики (ВТМ), поражающий растения рода табак (Nicosia). Особенностью этого вируса является то, что он имеет однонитчатую молекулу РНК, свернутую в виде спирали. Она находится внутри полого цилиндра, оболочка которого состоит из одинаковых белковых молекул — субъединиц. Установлено, что в такую белковую субъединицу входит 158 аминокислот и найдена последовательность расположения их в этих субъединицах. Размножение ВТМ происходит внутри растительной клетки лишь в том случае, если вирусная частица попадает через поврежденную клеточную оболочку. Чистая РНК ВТМ способна заражать клетку и воспроизводить белок для каждой вирусной частицы за счет аминокислот хозяина.
Источник: www.activestudy.info
Бактерии, как и все живые организмы, размножаются. Происходит это чаще всего путем простого поперечного деления в различных плоскостях. При этом образуются разнообразные сочетания клеток: парные соединения, одиночные клетки, гроздья, цепочки, пакеты и др.
Некоторые микроорганизмы размножаются спорообразованием (актиномицеты и грибы) и почкованием (дрожжи), у некоторых микроорганизмов наблюдается половое размножение, но большинство из них размножается бесполым (вегетативным) путем. При благоприятных условиях размножение протекает с необыкновенной быстротой — каждые 20-30 минут материнская бактериальная клетка делится на две дочерние. Дочерняя клетка со временем становится материнской и также делится. Таким образом, деление бактерий идет в геометрической прогрессии. Если бы такое деление шло беспрепятственно, то через 48 часов одна бактерия могла бы дать потомство в сотни биллионов клеток, а через пять дней такую массу, которая заполнила бы собой бассейны всех морей и океанов. Однако этого не происходит, поскольку на микроорганизмы действуют различные факторы окружающей среды.
Делению клетки предшествует равномерное увеличение общего азота, РНК и белка в цитоплазме. Затем происходит репликация (удвоение) ДНК. В делящейся клетке между спиралями ДНК разрываются водородные связи и образуются одиночные дочерние спирали ДНК (рис.25).
Рис. 25. Процесс бинарного деления палочковидных прокариот
1 — образование одиночных спиралей ДНК;
2 — удвоение (репликация) ДНК;
3 -вытягивание клетки;
— формирование перегородки;
4 — окончание формирования перегородки и образование выпуклой клеточной стенки;
5 -разделение клеток.
Сразу после репликации ДНК начинается вытяжение клетки и образование поперечной перегородки за счет двух слоев цитоплазматической мембраны, выпячивающейся навстречу друг другу. Чаще всего перегородка образуется посередине материнской клетки, в результате чего дочерние клетки имеют примерно одинаковые размеры. Между слоями перегородки идет формирование клеточной стенки.
Одиночная спираль ДНК в новых клетках служит матрицей для создания второй спирали, в результате чего образуется двойная спираль ДНК с восстановленными водородными связями и формируется новый нуклеоид.
В процессе размножения одна из половин клетки постоянно сохраняет жгутики. На конечном этапе размножения бактерий жгутики вырастают и у другой половины.
Рост и размножение микроорганизмов зависит от различных факторов окружающей среды и видовых характеристик. Наблюдение за развитием микроорганизмов, культивируемых в жидкой питательной среде в замкнутых резервуарах, показывает, что для роста биомассы необходимы наличие источника энергии, присутствие компонентов, необходимых для синтеза биомассы, отсутствие в среде ингибиторов, подавляющих рост клеток, поддержание в среде необходимых физико-химических условий. В этих условиях рост микроорганизмов условно можно подразделить на несколько последовательных фаз или периодов (рис. 26):
Рис. 26. Типичная кривая роста популяции микроорганизмов 1 — лаг-фаза;
2 — фаза ускоренного роста; 3 — фаза логарифмического (экспоненциального) роста;
4 — фаза замедления роста; 5 — фаза стационарного роста; 6 — фаза старения и отмирания.
1. лаг-фаза (англ. lag — запаздывание) — период между посевом бактерий и началом размножения. В этот период происходит адаптация бактериальной культуры к питательной среде. Она проявляется в накоплении оптимального количества необходимых ферментов, в инактивации некоторого ингибитора, присутствующего в среде, в прорастании спор и др. При благоприятных условиях бактерии увеличиваются в размерах и готовятся к делению. Лаг-фаза может длиться от 10 минут до нескольких часов, но в среднем она составляет 4-5 часов.
2. Фаза ускоренного роста наблюдается после лаг-фазы и характеризуется нарастанием темпов деления микроорганизмов и накопления биомассы.
3. Фаза логарифмического или экспоненциального роста является периодом наиболее интенсивного деления бактерий. Бактерии делятся каждые 20-40 минут. Во время этой фазы бактерии особенно ранимы, что объясняется высокой чувствительностью растущих клеток к факторам окружающей среды. Продолжительность экспоненциального роста зависит от концентрации питательных веществ в субстрате и в среднем составляет 5-6 часов.
4. Фаза замедления роста является переходным периодом от экспоненциального роста к фазе стационарного роста. Во время этой фазы наблюдается истощение питательных веществ субстрата и накопление в нем продуктов метаболизма, что снижает интенсивность размножения микроорганизмов.
5. Фаза стационарного роста вызывается постепенным истощением среды, накоплением в ней литических ферментов, химическим ингибированием роста микробной клетки продуктами метаболизма. Эта фаза отличается от предыдущей повышенной сопротивляемостью бактерий многим химическим и физическим факторам. К началу этой фазы количество жизнеспособных клеток достигает максимального уровня и остается на этом максимуме в течение нескольких часов в зависимости от вида микроорганизмов и особенностей их культивирования. В конце этой фазы у некоторых микроорганизмов наблюдается процесс спорообразования.
6. Завершающая фаза процесса размножения — фаза старения и гибели — характеризуется отмиранием бактерий из-за истощения питательной среды и накопления в ней продуктов метаболизма. Наблюдается автолиз микроорганизмов как экстремальное проявление нестабильности клетки после прекращения роста. Продолжительность этой фазы может составлять от нескольких часов до нескольких недель.
Источник: studopedia.org
Рост бактерий – увеличение бактериальной клетки в размерах без увеличения числа особей в популяции.
Размножение бактерий – процесс, обеспечивающий увеличение числа особей в популяции. Бактерии характеризуются высокой скоростью размножения.
Рост всегда предшествует размножению. Бактерии размножаются поперечным бинарным делением, при котором из одной материнской клетки образуются две одинаковые дочерние.
Процесс деления бактериальной клетки начинается с репликации хромосомной ДНК. В точке прикрепления хромосомы к цитоплазматической мембране (точке-репликаторе) действует белок-инициатор, который вызывает разрыв кольца хромосомы, и далее идет деспирализация ее нитей. Нити раскручиваются, и вторая нить прикрепляется к цитоплазматической мембране в точке-прорепликаторе, которая диаметрально противоположна точке-репликатору. За счет ДНК-полимераз по матрице каждой нити достраивается точная ее копия. Удвоение генетического материала – сигнал для удвоения числа органелл. В септальных мезосомах идет построение перегородки, делящей клетку пополам.
Двухнитевая ДНК спирализуется, скручивается в кольцо в точке прикрепления к цитоплазматической мембране. Это является сигналом для расхождения клеток по септе. Образуются две дочерние особи.
На плотных питательных средах бактерии образуют скопления клеток – колонии, различные по размерам, форме, поверхности, окраске и т. д. На жидких средах рост бактерий характеризуется образованием пленки на поверхности питательной среды, равномерного помутнения или осадка.
Размножение бактерий определяется временем генерации. Это период, в течение которого осуществляется деление клетки. Продолжительность генерации зависит от вида бактерий, возраста, состава питательной среды, температуры и др.
Фазы размножение бактериальной клетки на жидкой питательной среде:
1) начальная стационарная фаза; то количество бактерий, которое попало в питательную среду и в ней находится;
2) лаг-фаза (фаза покоя); продолжительность – 3–4 ч, происходит адаптация бактерий к питательной среде, начинается активный рост клеток, но активного размножения еще нет; в это время увеличивается количество белка, РНК;
3) фаза логарифмического размножения; активно идут процессы размножения клеток в популяции, размножение преобладает над гибелью;
4) максимальная стационарная фаза; бактерии достигают максимальной концентрации, т. е. максимального количества жизнеспособных особей в популяции; количество погибших бактерий равно количеству образующихся; дальнейшего увеличения числа особей не происходит;
5) фаза ускоренной гибели; процессы гибели преобладают над процессом размножения, так как истощаются питательные субстраты в среде. Накапливаются токсические продукты, продукты метаболизма. Этой фазы можно избежать, если использовать метод проточного культивирования: из питательной среды постоянно удаляются продукты метаболизма и восполняются питательные вещества.
Следующая глава >
Источник: bio.wikireading.ru
Способы размножения микроорганизмов
I. Половой способ размножения наблюдается только у эукариот.
II. Бесполые способы размножения.
-
Равновеликое бинарное поперечное деление (простое деление, изоморфное деление, митоз) наблюдается у большинства одноклеточных микроорганизмов (бактерий, риккетсий, простейших, дрожжей), в результате образуются две новые дочерние полноценные особи, наделенные генетической информацией материнской клетки, симметричные в отношении продольной и поперечной оси, сама материнская клетка исчезает.
При этом у большинства Грам+ бактерий деление происходит путем синтеза поперечной перегородки, идущей от периферии к центру (рис. 63А). Клетки большинства Грам- бактерий делятся путем перетяжки клетки (клетка истончается посередине) (рис. 63Б).
-
Почкование (неравновеликое бинарное деление) наблюдается у представителей родов Francisella и Mycoplasma и дрожжеподобных грибов. При почковании материнская клетка дает начало дочерней клетке: на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки, после чего отделяется. КС почки полностью синтезируется заново (рис. 63В). В процессе почкования симметрия наблюдается в отношении только продольной оси. Между материнской и дочерней клетками существуют морфологические и физиологические различия. Новая дочерняя клетка лучше приспосабливается к меняющимся условиям.
-
Фрагментация нитевидных форм характерна для рода Actinomyces и Mycoplasma.
-
Образование экзоспор характерно для Streptomycetes, дрожжеподобных и плесневых грибов.
-
Особый цикл развития наблюдается у Chlamydia.К делению в клетках макроорганизма способны лишь вегетативные формы хламидий (ретикулярные или инициальные тельца). Их цикл, состоящий из нескольких делений, завершается образованием промежуточных форм, из которых формируются элементарные тельца, дающие начало вегетативным формам. После разрушения стенки вакуоли и клетки-хозяина элементарные тельца высвобождаются, и цикл повторяется. Цикл длится 40–48 ч.
-
Множественное деление описано для одной группы одноклеточных цианобактерий. В основе множественного деления лежит принцип равновеликого бинарного деления. Отличие заключается в том, что в этом случае после бинарного деления не происходит роста образовавшихся дочерних клеток, а они снова подвергаются делению (рис. 63Г).
|
|
|
Рис. 63.Способы деления и синтез КС у прокариот: А — деление путем образования поперечной перегородки; Б —деление путем перетяжки; В —почкование; Г —множественное деление. 1 —КС (толстой линией обозначена КС материнской клетки, тонкой—заново синтезированная); 2 —ЦПМ; 3 — мембранная структура; 4 — цитоплазма, в центре которой расположен нуклеоид; 5 —дополнительный фибриллярный слой КС |
Множественное деление (шизогония) описано также у простейших (малярийных плазмодиев): ядерный материал делится на множество ядрышек, окружается участками цитоплазмы, в результате образуется множество дочерних клеток.
Механизм и фазы простого деления
А. Рост до определенной степени зрелости. Рост клетки не беспределен и после достижения определенных размеров бактериальная клетка начинает делиться. Во время деления рост клетки замедляется и начинается вновь после деления.
Б. Кариокинез (репликация ДНК и деление нуклеоида). Из созревшей цитоплазмы поступает сигнал, который активирует ген-инициатор на ДНК. Микроорганизмы под действием гена-инициатора синтезируют белок-инициатор, который действует на ген-репликатор — специальный участок ДНК, с которого начинается удвоение ДНК и деление на две нити.
Деление молекулы ДНК (репликация) происходит по полуконсервативному механизму и в норме всегда предшествует делению клетки. Репликация ДНК начинается в точке прикрепления кольцевой хромосомы к ЦПМ, где локализован ферментативный аппарат, ответственный за репликацию.
|
Контакт ДНК с ЦПМ осуществляется посредством мезосом. Репликация, начавшаяся в точке прикрепления, идет затем в двух противоположных направлениях. Возникающие дочерние хромосомы остаются прикрепленными к мембране (рис. 64).
|
|
Механизм репликации ДНК выражается в разрыве водородных связей между ее двумя полинуклеотидными цепями, раскручивании их и синтезе с помощью ДНК-полимеразы вдоль каждой старой цепи новых цепей с комплементарной последовательностью оснований. После расхождения в дочерние клетки по одной старой и одной новой полинуклеотидной цепи между ними восстанавливаются водородные связи и формируется полуконсервативная двухцепочечная ДНК.
В норме существует определенная временная связь между репликацией хромосомы и делением бактериальной клетки. Воздействия различными химическими веществами и физическими факторами, приводящие к подавлению репликации ДНК, останавливают и клеточное деление. Однако при некоторых условиях связь между обоими процессами может быть нарушена, и клетки способны делиться в отсутствие синтеза ДНК.
В. Цитокинез (деление клетки). Параллельно с репликацией молекул ДНК происходит синтез мембраны рядом с мезосомой, в области контакта ДНК с ЦПМ. Образование перегородки приводит к делению клетки. Моментом, инициирующим деление клетки, является окончание репликации ДНК. Это приводит к разделению дочерних молекул ДНК и оформлению обособленных хромосом. Вновь образованные дочерние клетки отделяются друг от друга.
Угнетение синтеза мембраны до окончания репликации приводит к нарушению процесса деления: клетка перестает делиться и растет в длину. У некоторых бактерий образование перегородки не приводит к разделению клеток: образуются многокамерные клетки.
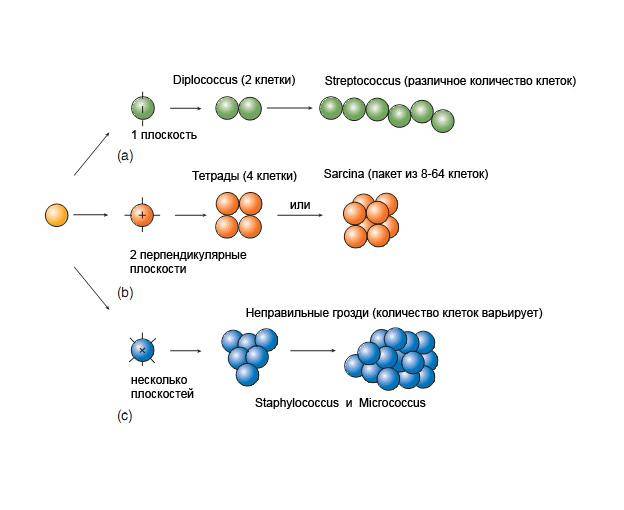
Г. Расхождение образовавшихся дочерних клеток происходит в результате лизиса среднего слоя КС. Если после многократного деления в одной плоскости клетки не расходятся, образуются цепочки палочковидных (Bacillus) или сферических (Streptococcus) клеток или парные клетки (Neisseria). Разъединение клеток возможно с обособлением одной из клеток путем движения по поверхности другой, в результате бактерии располагаются беспорядочно (Escherichia).Если при разъединении одна из дочерних клеток, не отрываясь от точки деления, передвигается по дуге, создается V-образная форма (Corynebacterium, Bifidobacterium). После бинарного деления и расхождения клеток в нескольких плоскостях образуются клеточные скопления разной формы: гроздья (Staphylococcus), пакеты (Sarcina) (рис. 65). Если деление нуклеоида предшествует клеточному делению, образуются многонуклеоидные микроорганизмы. Под влиянием неблагоприятных внешних факторов (соли желчных кислот, УФ-лучи, ПАВ, антибиотики) деление клетки может остановиться с сохранением ее роста. В таком случае возможно образование удлиненных нитевидных клеток.
Рис. 65.Деление кокков
Период генерации — интервал времени, в течение которого происходит удвоение количества бактерий Скорость размножения микроорганизмов и период генерации зависят от вида микроорганизма, величины и свойств инокулята, состава питательной среды, ее рН, аэрации, температуры инкубации, других факторов. При благоприятных условиях у многих микроорганизмов деление происходит через 15–30 мин (E. coli, S. typhi). У прихотливых микроорганизмов деление осуществляется через 45–90 мин (Streptococcus, Corynebacterium) и даже через 18 ч (M. tuberculosis).
Источник: StudFiles.net