Цитоскелет рисунок
Глава 6. Цитоскелет.
Микротрубочки ! Механизм образования пищеварительной вакуоли при фагоцитозе. Актомиозин. Клеточный цикл. Митоз. ! Механизм деления цитоплазмы в клетках растений
Любой из нас имеет скелет. Он состоит из твердых костей, гибких связок, соединяющих кости между собой, и мягких мышц, которые прикреплены к костям и, с силой меняя форму, изменяют взаимное расположение разных костей и мягких тканей тела относительно костей. В клетке имеются специальные белки, играющие роль костей и мышц. Всю систему таких белков называют цитоскелетом.
Микротрубочки
Рисунок 43
Микротрубочки (рис. 43) полностью соответствуют своему названию. Это прямые микроскопические трубочки (наружный диаметр 28 нм, внутренний — 14 нм), состоящие из двух похожих друг на друга белков a-тубулина (a — греческая буква альфа, все слово читается "альфа-тубулин") и в-тубулина ("бета-тубулин").
а конца микротрубочки отличаются друг от друга некоторыми важными свойствами (их называют "+" и "-"-концы). В ДНК клетки имеются два разных гена, содержащие информацию о последовательностях аминокислот а-тубулина и в-тубулина. После синтеза на рибосомах в цитоплазме молекулы а- и в-тубулина объединяются в димеры ("ди" — "два", "мерос" — "часть"). Димеры тубулина при определенных условиях могут присоединяться к "+"-концу микротрубочки, микротрубочка при этом удлиняется. С "-"-конца микротрубочки могут разбираться (то есть от него отделяются димеры тубулина, и микротрубочка при этом укорачивается). Изменяя условия в разных частях цитоплазмы, клетка имеет возможность делать сеть микротрубочек в ней более или, наоборот, менее густой. Кроме того, есть белки, способные присоединяться к "+"-концам микротрубочек, прекращая тем самым их сборку, и другие белки, способные присоединяться к "-"-концам и прекращать разборку микротрубочек (вместе они называются “кэпирующие белки”).
Известны специальные транспортные белки, способные перетаскивать по микротрубочкам различные органоиды клетки. Один из них, кинезин, переносит их в направлении от "-"- к "+"-концу.
! Механизм образования пищеварительной вакуоли при фагоцитозе
В большинстве клеток работают два независимых механизма.
Первый из них — простое следствие механизма прилипания пищевой частицы к мембране. За счет теплового движения молекул воды и пищевая частица, и рецепторы мембраны все время слегка вибрируют. Поэтому близко расположенные, но еще не соединившиеся друг с другом рецепторы и лиганды через короткое время сталкиваются и слипаются. Получается, что мембрана все больше и больше налипает на пищевую частицу со всех сторон (рис. 13а)).
Рисунок 44
Второй механизм обеспечивается работой специальных белков, одним концом присоединяющихся к рецепторам мембраны, уже прилипшим к лигандам на пищевой частице, а другим — к расположенным под мембраной микротрубочкам. Эти белки способны двигаться по микротрубочкам вглубь цитоплазмы, "волоча за собой" рецепторы, закрепленные в мембране. В результате работы многих таких белков весь кусок мембраны, прилипший к пищевой частице, погружается внутрь клетки, "на ходу" замыкаясь в пузырек (рис. 44).
Актомиозин.
Рисунок 45
Актомиозин — комплекс из молекул 4-х разных белков (а именно актина, тропонина, тропомиозина и миозина) в виде нитей в цитоплазме, способных с силой укорачиваться.
Рисунок 46
В результате синтеза белка на актиновой иРНК от рибосом отделяются молекулы G-актина (рис. 45а)). В цитоплазме они слипаются друг с другом в нити F-актина. Молекулы тропомиозина тоже сначала слипаются друг с другом в нити, а затем такие нити присоединяются к двум желобкам каждой нити F-актина. На нить F-актина садятся также молекулы тропонина (рис. 45б)). Молекула тропонина состоит из трех субъединиц. Одна из них способна присоединяться к F-актину, вторая — к тропомиозину, а третья соединяет первые две, прикрепляясь одним концом к первой, а другим — ко второй. Нить, состоящую из этих трех белков, называют актиновым филаментом, или микрофиламентом. При появлении в растворе ионов кальция третья субъединица тропонина удлиняется, извлекая нити тропомиозина из желобков F-актина (рис. 45в)), при исчезновении кальция из раствора эта субъединица укорачивается, возвращая нити тропомиозина обратно в желобки.
Рисунок 47
Молекула миозина состоит из двух "головок" и "хвоста". Такие молекулы в цитоплазме могут слипаться друг с другом, образуя нити миозина (рис. 46). "Головки" молекул миозина образуют на поверхности нити миозина шесть продольных рядов. Отдельная молекула миозина в присутствии ионов кальция и АТФ перемещается по микрофиламенту в направлении от своего "хвоста"”, цепляясь “головками” за желобки F-актина.
ть миозина может присоединить максимум 12 актиновых филаментов (по 6 с каждого конца), и затем в присутствии ионов кальция и АТФ (подробно про ионы кальция рассказано в главе 7, а про АТФ — в главе 9) "тащить" их друг к другу до соприкосновения (рис. 47а)). Выяснилось, что в некоторых клетках миозин образует димеры (рис. 47б)). Димер миозина может перемещать один микрофиламент по другому.
Клеточный цикл. Митоз.
Доказано, что новые живые клетки могут возникать одним-единственным способом — в результате деления клеток. В ядре каждой клетки имеются молекулы ДНК, содержащие информацию об аминокислотном составе всех ее белков. Обе клетки, возникающие в результате деления, должны получить полноценные копии всех молекул ДНК материнской клетки. Для этого все молекулы ДНК материнской клетки должны быть сначала удвоены (период в жизни клетки, когда в ней происходит удвоение (репликация) ДНК, называется S-фазой клеточного цикла), а во время деления клетки — распределены по обеим дочерним клеткам.
Рисунок 48
Клеточный цикл — это последовательность событий, связанных с размножением клетки (рис. 48). Он состоит из собственно деления клетки (митоза), паузы до начала удвоения ДНК (G1-фаза), удвоения ДНК (S-фаза) и паузы от момента окончания S-фазы до начала митоза (G2-фазы). G1-, S- и G2-фазы вместе называются интерфазой.
Рисунок 49
Молекулы ДНК в G2-фазе перед началом митоза подвергаются тщательной упаковке с помощью специальных белков (рис. 49). Результат этой упаковки — митотическая хромосома. Перед началом митоза внутри ядра под микроскопом становятся видны хромосомы (упакованные молекулы ДНК, соединенные попарно центромерами с помощью специальных белковых “замков” — кинетохоров). Каждая такая пара молекул ДНК — "сестры", получившиеся при удвоении одной молекулы ДНК клетки. При митозе им предстоит разойтись по разным дочерним клеткам.
Сам митоз состоит из четырех фаз: профазы, метафазы, анафазы и телофазы.
Рисунок 50
В профазе (рис. 50а)) происходит удвоение центриолей (каждая из двух центриолей материнской клетки строит около себя дочернюю центриоль) и две пары центриолей расходятся в разные концы (принято говорить: на разные полюса) делящейся клетки. После этого около каждой пары центриолей начинается сборка микротрубочек (при этом их "+"-концы обращены от центриолей в цитоплазму). В результате образуется веретено деления, состоящее из двух половинок (полуверетен) с парой центриолей в вершине каждой из них. В конце профазы оболочка ядра распадается на мелкие мембранные пузырьки (в конце митоза из них будут собраны два новых ядра), и хромосомы оказываются в цитоплазме.
В метафазе (рис. 50б)) "+"-концы микротрубочек прикрепляются к кинетохорам хромосом. Первый из этих "+"-концов может прикрепиться к кинетохору с любой стороны. Далее возможны два варианта развития событий. Если "+"-конец второй микротрубочки прикрепится к кинетохору с той же стороны, что и первый, то в следующий момент кинетохор отделяется от обеих микротрубочек, и все начинается сначала. Если же "+"-конец второй микротрубочки прикрепится к кинетохору со стороны другого полюса клетки, то кинетохор прочно прикрепляется к обеим микротрубочкам. Что происходит дальше, не вполне понятно. Почему-то сборка и разборка прикрепившихся к кинетохорам хромосом микротрубочек происходят так, что все хромосомы выстраиваются в плоскости экватора делящейся клетки. Известно, что если с помощью тонкой стеклянной иглы помешать одной хромосоме добраться до этой плоскости, митоз приостановится до тех пор, пока эта хромосома не займет свое место.
Рисунок 51
Когда все хромосомы выстраиваются в экваториальной плоскости, специальные белки разрезают кинетохоры пополам, так, что "сестринские" молекулы ДНК (с момента разрезания кинетохора каждую из них можно называть отдельной хромосомой) отделяются друг от друга и начинают расходиться к разным полюсам клетки. Это — момент начала анафазы (рис. 50в)). Полуверетена в анафазе расходятся в разные стороны, причем каждое из них двигается как единое целое. Расхождение происходит за счет работы молекул белков, похожих на кинезин. Каждая такая молекула, прикрепившись к микротрубочке одного полуверетена, тащит ее по микротрубочке второго полуверетена в направлении к "+"-концу (рис. 51).
В телофазе (рис. 50г)) происходит разборка микротрубочек веретена деления и образование двух ядер из мембранных пузырьков вокруг двух групп хромосом на полюсах клетки. Если стеклянной иглой отделить одну из хромосом от группы, то вокруг нее образуется отдельное маленькое ядро.
Последний этап митоза — деление цитоплазмы. У животных под мембраной клетки в районе ее экватора формируется кольцевой пучок актомиозина. Поочередно сокращаясь и перестраиваясь, он постепенно пережимает цитоплазму пополам, увлекая за собой мембрану.
! Механизм деления цитоплазмы в клетках растений
Рисунок 52
У растений экваториальная плоскость заполняется мембранными пузырьками, затем они сливаются друг с другом, разделяя цитоплазму на две части (рис. 52).
? Какие выводы можно сделать из опытов, описанных в рассказе про деление клетки? Предложите гипотезы:
- о том, что мешает белкам, разрезающим кинетохоры хромосом, начать это делать до того, как все хромосомы окажутся в экваториальной плоскости клетки;
- о том, что заставляет мембранные пузырьки в телофазе митоза собираться вокруг хромосом.
Источник: lasch.narod.ru
Цитоскелет выполняет три главные функции.
1. Служит клетке механическим каркасом, который придаёт клетке типичную форму и обеспечивает связь между мембранной и органеллами. Каркас представляет собой динамичную структуру, которая постоянно обновляется по мере изменения внешних условий и состояния клетки.
2. Действует как «мотор» для клеточного движения. Двигательные (сократительные) белки содержатся не только в мышечных клетках, но и в других тканях. Компоненты цитоскелета определяют направление и координируют движение, деление, изменение формы клеток в процессе роста, перемещение органелл, движение цитоплазмы.
3. Служит в качестве «рельсов» для транспорта органелл и других крупных комплексов внутри клетки.
Микрофиламенты и промежуточные волокна.
Микрофиламенты построенные из F-актина пронизывают микроворсинки, образуя узлы. Эти микроволокна удерживаются вместе с помощью актинсвязывающих белков, наиболее важными из которых являются фимбрин и виллин. Кальмодулин и миозиноподобная АТФ – аза соединяют крайние микроволокна с плазматической мембраной. .
Клетка может менять набор синтезируемых белков цитоскелета в зависимости от условий, но процесс этот медленный. Конструкция цитоскелета способна быстро меняться даже без синтеза новых молекул, за счет полимеризации и деполимеризации нитей. В клетке все время идет обмен между нитями и раствором белков-мономеров в цитоплазме. Во многих клетках примерно половина молекул актина и тубулина находится в виде мономеров в цитоплазме и половина входит в состав нитей микрофиламентов. Клетка регулирует стабильность нитей цитоскелета, присоединяя к ним специальные белки, изменяющие скорость полимеризации. Общий принцип функционирования цитоскелета – динамическая нестабильность. Например, форму эритроцита в виде двояковогнутого диска поддерживает примембранный цитоскелет из волокон, образованных белком спектрином. Спектрин связан с белком анкерином (anchor – якорь), который соединяется с белком цитоплазматической мембраны, ответственным за транспорт анионов (Cl— , HCO—3). Дефекты белков спектрина и анкирина вызывают необычную форму эритроцитов. Такие эритроциты очень быстро разрушаются в селезенке. Болезни, вызываемые такими нарушениями, называют наследственным сфероцитозом или наследственным эллиптоцитозом.

Рис. Цитоскелет эукариот. Актиновые микрофиламенты окрашены в красный, микротрубочки — в зеленый, ядра клеток — в голубой цвет.
Кератиновые промежуточные филаменты в клетке.
Таким образом, эукариотические клетки обладают своего рода каркасом, который с одной стороны придает им определенную форму, а с другой допускает возможность её изменения, позволяя клеткам двигаться и перемещать свои органеллы с одной части клетки в другую. Кроме основных компонентов цитоскелета важную роль в его организации и функциональной интеграции играют вспомогательные белки. Эти белки отвечают за прикрепление органелл к цитоскелету, обеспечение направленного движения органелл, координацию функций цитоскелета.
Нарушения цитоскелета. Цитоскелет не является пассивной клеточной структурой, обеспечивающей только клеточную морфологию. Доказана роль цитоскелета в двигательной функции клеток, в структуре плазматической мембраны и, что очень важно, в рецепторной функции клеток. Отмечено, что изменения цитоскелета нарушают процесс высвобождения активного вещества (гормона, медиатора и т.д.), а также изменяют рецепторную функцию клеток-мишеней. В результате нарушается рецепция клетками (в частности, нервными) различных стимулирующих веществ. Кроме того, отмечается нарушение двигательной активности клеток (например, бета-клеток поджелудочной железы), в результате возникает недостаточность инсулина. Поэтому проявления диабета довольно постоянны при хромосомных синдромах (Тернера, Клайнфельтера, Дауна и т.п.). Другим примером заболеваний с нарушением цитоскелета являются мышечная дистрофия Дюшенна и мышечная дистрофия Беккера. Обе формы являются результатом мутаций гена, кодирующего белок дистрофин. Дистрофин, в свою очередь, входит в состав цитоскелета. В результате при биопсии мышц выявляют характерные изменения – перерождение мышц и некроз волокон.
Органеллы, содержащие триплеты микротрубочек
Центриоли. Центриоль имеет цилиндрическую форму, диаметр 150 нм и длину 500 нм; стенка образована 9 триплетами (триплетный – состоящий из трёх) микротрубочек. Центриоль – центр организации митотического веретена – участвует в делении клетки. В ходе фазы S клеточного цикла центриоли удваиваются. Образовавшаяся новая центриоль расположена под прямым углом к первоначальной центриоли. При митозе пары центриолей, каждая из которых состоит из первоначальной и вновь образованной, расходятся к полюсам клетки и участвуют в образовании митотического веретена.
Базальное тельце состоит из 9 триплетов микротрубочек, расположенных в основании реснички или жгутика; служит матрицей при организации аксонемы.
Аксонема состоит из 9 периферических пар микротрубочек и двух расположенных центрально одиночных микротрубочек. В каждой периферической паре микротрубочек различают субфибриллу А и субфибриллу В. С субфибриллой А связаны так называемые наружные и внутренние ручки. В их состав входит белок динеин, обладающий способностью расщеплять АТФ. Аксонема – основной структурный элемент реснички и жгутика.
Ресничка – вырост клетки длиной 5-10мкм и толщиной 0,2 мкм, содержащий аксонему. Реснички присутствуют в эпителиальных клетках воздухопроводящих и половых путей; перемещают слизь с инородными частицами и остатками отмерших клеток и создают ток жидкости около клеточной поверхности. Под влиянием табачного дыма реснички воздухоносных путей разрушаются, что способствует задержке секрета в бронхах.
Рис. Схема поперечного сечения реснички. (Из кн. Б. Албертс и др. «Молекулярная биология клетки», том 3.)
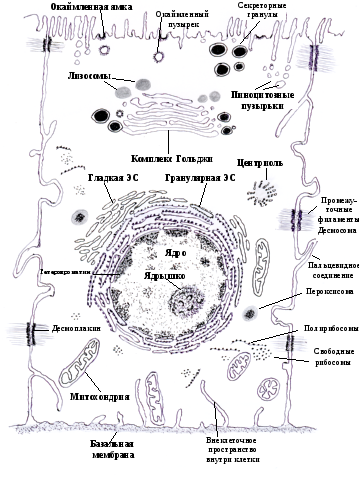
Схема строения эукариотической эпителиальной клетки
Рисунок В.П. Андреева
Внутриклеточное пространство внутри клетки – это зона цитозоля неструктурированного мембранами внутриклеточного содержимого. Цитозоль является жидкой частью цитоплазмы и составляет около половины объема клетки. Здесь синтезируются белки, часть которых собирается на полисомах и остается в цитозоле. Цитозоль непосредственно сообщается через крупные ядерные поры с содержимым ядра. В ядре идут процессы транскрипции РНК с ДНК, причем синтезируются как нормальные клеточные, так и вирусные при вирусных инфекциях клеток. РНК из ядра транспортируется для синтеза белка в цитозоль на полирибосомы. Синтезированные белки под контролем шаперонов («катализаторов» принятия полипептидной цепью биологически значимой конформации) направляются в специальные участки эндоплазматического ретикулума. Лишние, испорченные, а также вирусные белки расщепляются в цитозоле так называемыми протеасомами. «Протеасомы» представляют собой мультипротеазные комплексы, состоящие из 28 субъединиц. Протеасомы расщепляют вирусные белки до пептидов- антигенов. Образовавшиеся пептиды- антигены вступают в связь с молекулами главного комплекса гистосовместимости (ГКГ – I), и направляются для экспрессии на клеточную мембрану. Комплексы антиген – ГКГ- I, расположенные на клеточной мембране, узнаются СД8+ Т- лимфоцитами, которые при этом активируются и обеспечивают противовирусную защиту, а также защиту от цитозольных внутриклеточных инфекций.
Внеклеточное пространство внутри клетки – это пространство (зона, компартмент) связанное с внешней внеклеточной средой и ограниченное мембранами структур и везикул, включающее в себя аппарат Гольджи, эндоплазматический ретикулум, лизосомы, эндосомы, фагосомы и фаголизосомы. Особое значение эта зона имеет в структуре антигенпредставляющих клеток, к которым относятся макрофаги и дендритные клетки (вариант лимфоцитов). На рибосомах эндоплазматической сети этих клеток синтезируются цепи молекул главного комплекса гистосовместимости (ГКГ- III). Конформация этих молекул произойдет только в том случае, если они соединятся с пептидами, образующимися в результате протеолиза (расщепления) белков – антигенов, захваченных клеткой посредством эндоцитоза или фагоцитоза. Это происходит тогда, когда фаголизосомы сливаются с везикулами, содержащими несконформированные молекулы ГКГ- II. С участием пептида молекула ГКГ- II принимает правильную конформацию, продвигается к мембране и экспрессируется на ней. Комплексы антигенов-пептидов с молекулами ГКГ- II распознают СД4+ Т – лимфоциты, которые играют главную роль в защитных реакциях от внеклеточных инфекций.
Концепции современной цитологии
Для разных клеточных типов у различных организмов характерны универсальные процессы. Это передача сигналов внутри клетки, регуляция клеточного цикла, апоптоз, тепловой шок, деградация внутриклеточных белков.
Апоптоз – биологический механизм гибели клетки по тому или иному сигналу извне или изнутри, который активирует внутри клетки определенные системы ферментов, обеспечивающих повреждение митохондрий, фрагментацию ДНК и затем фрагментацию ядра и цитоплазмы клетки. В результате клетка распадается на окруженные мембраной апоптозные тельца, которые могут фагоцитироваться соседними эпителиальными клетками и макрофагами. Содержимое погибающей клетки не попадает во внеклеточную среду. В ткани не развивается воспаление. Жизнь многоклеточных организмов невозможна без запрограммированной клеточной гибели, которая регулирует развитие, тканевый гомеостаз, клеточный ответ на повреждение ДНК и старение.
Тепловой шок
Тепловой шок может вызываться не только слишком высокой, но и слишком низкой температурой, ядами и множеством других воздействий, например, сбоем цикла суточной активности. Под воздействием этих факторов в клетке появляются белки с «неправильной» третичной структурой. Многие белки теплового шока как раз и помогают переводить в раствор и вновь сворачивать денатурированные или неправильно свернутые белки.
Реакция теплового шока сопровождается прекращением синтеза обычных для клетки белков и ускоренным синтезом различных защитных белков. Эти белки защищают от повреждений ДНК, матричные РНК, предшественники рибосом, и прочие важные для клетки структуры. Реакция теплового шока необычайно древняя и консервативная. Некоторые белки теплового шока обнаруживают гомологию у бактерий и человека.
К N-концу поврежденных, изношенных, недостроенных и функционально неактивных белков присоединяются молекулы белка-убиквитина, делая их мишенью для ферментов класса протеаз. Ассоциированный с убиквитином белок разрушается в особых мультикомпонентных комплексах, называемых протеасомами. Убиквитин – пример белка теплового шока, функционирующий в клетке и в нормальных условиях. В некоторых клетках, синтезируется до 30% аномальных белков. За открытие роли убиквитина в деградации белков была присуждена в 2004 году Нобелевская премия по химии.
Шапероны (от англ. букв.- пожилая дама, сопровождающая молодую девушку на балах) – семейство специализированных внутриклеточных белков, обеспечивающих быстрое и правильное сворачивание (фолдинг) вновь синтезированных молекул белка.
Кроме этого известны и другие белки шапероны. Например, шаперон HSP 70. Его синтез активируется при многих стрессах, в частности при тепловом шоке (отсюда и название Heart shook protein 70 – белок теплового шока). Цифра 70 означает молекулярную массу в килодальтонах. Основная функция этого белка – предотвращение денатурации других белков при повышении температуры. Шапероны – одни из самых жизненно важных белков всех живых существ. Они возникли на самых ранних стадиях эволюции, возможно еще до разделения организмов на прокариоты и эукариоты
Передача внешнего сигнала в клетку
Клетки не могут сами принять решение о том, что нужно организму. Они должны получить сигнал извне и лишь после этого внутриклеточная регуляция включится в поддержание необходимых процессов. Известные биохимики Вильям Эллиот и Дафна Эллиот приводят аналогию с мореплаванием. «Каждый корабль представляет собой организационную единицу «клетку», где поддерживается порядок и дисциплина, упорядоченно работают все механизмы и т.д. Вместе с тем, цели и маршруты плавания для кораблей определяются внешними сигналами (гормонами) высшего руководства (эндокринные железы и мозг).
Клетка обычно принимает сигнал о «состоянии дел» вокруг нее с помощью рецепторов. Н.Н. Мушкамбаров и С.Л. Кузнецов выделяют несколько механизмов действия сигнальных веществ.
1) Вещество взаимодействует с рецептором плазмолеммы, что индуцирует передачу сигнала внутрь клетки и при этом происходит химическая модификация (фосфорилирование, дефосфорилирование) определенных белков. (Фосфорильная группа несет сильный отрицательный заряд, что способствует изменению конформации белковой молекулы).
2) Вещество взаимодействует с рецептором плазмолеммы, который является одновременно и ионным каналом, открывающимся при связывании регулятора.
3) Внеклеточный регулятор проникает внутрь клетки мишени, связывается с цитоплазматическим или ядерным белком-рецептором и, выступая после этого как транскрипционный фактор, влияет на экспрессию определенных генов. Так действуют гормоны стероидной природы (например, мужские и женские половые гормоны).
В качестве сигнальных молекул иногда выступают простагландины и NO (оксид азота). Они проникают в клетку-мишень и влияют на активность регуляторных ферментов. Конечный результат – модификация определенных белков.
Наиболее часто используемым является механизм первого типа. При этом конкретные способы его реализации весьма разнообразны.
Передача сигналов внутри клетки
Водорастворимые сигнальные молекулы, в том числе известные нейромедиаторы, пептидные гормоны и факторы роста, присоединяются к специфическим белковым рецепторам на поверхности клеток-мишеней. Поверхностные рецепторы связывают сигнальную молекулу (лиганд), проявляя большое сродство к ней, и это внеклеточное событие порождает внутриклеточный сигнал, изменяющий поведение клетки.
Рецепторы являются интегральными мембранными белками.
Существует множество сигнальных путей, начинающихся от мембранного рецептора.
(Изменение мембранных рецепторов сопровождается возникновением различных болезней. Так, например, дефект в рецепторе мужского полового гормона тестостерона приводит к тому, что особи с мужским генотипом (2А+ХУ) выглядят как самки; все млекопитающие, не подвергнувшиеся в эмбриональный период воздействию тестостерона, развиваются по женскому пути. Мутантные самцы имеют нормальные семенники, вырабатывающие тестостерон, но ткани этих самцов не реагируют на гормон из-за дефектности соответствующих рецепторов. В результате у таких самцов развиваются все вторичные половые признаки самок и их семенники не опускаются в мошонку, а остаются в брюшной полости. Этот синдром (тестикулярной феминизации или сидром Морриса) встречается у мышей, крыс, крупного рогатого скота, а также у человека. Хотя изменен только ген, кодирующий рецептор тестостерона, затронутыми оказываются все разнообразные типы клеток, в норме реагирующие на этот гормон. Таким образом, один внешний сигнал может включать различные наборы генов в клетках разного типа.
Подавляющее большинство поверхностных рецепторов для гидрофильных сигнальных молекул, связав лиганд на внешней стороне мембраны, претерпевает конформационное изменение. Это изменение создает внутриклеточный сигнал, изменяющий поведение клетки-мишени. Внутриклеточные сигнальные молекулы часто называют вторыми посредниками (мессенджерами, англ. messenger – посыльный), считая «первым посредником» внеклеточный лиганд. К вторичным (внутриклеточным) посредникам относят циклический аденозинмонофосфат (цАМФ), циклический гуанозин 3΄,5΄ — монофосфат (цГМФ), катионы кальция, инозит-1,4,5-трифосфат, диацилглицерин. Кроме этого, известны сигнальные пути опосредованные белками, липидами, в том числе свободными жирными кислотами, оксидом азота (NO), а также пути не содержащие вторичного посредника. Примером последнего варианта является влияние γ-интерферона на транскрипцию определенных генов, с антивирусной направленностью. Внутриклеточные сигнальные пути регуляции клеточной активности очень сложны, до конца не изучены и многие открытия еще впереди. Достаточно сказать, что внутриклеточный сигнальный путь с участием инсулина, несмотря на многолетние исследования, еще не расшифрован.
Источник: StudFiles.net
На фото, сделанном с помощью конфокального микроскопа, — миобласт C2C12 с помеченным ядром (синее), актиновыми филаментами (красные) и микротрубочками (зеленые).
Многие из нас в школе на уроках биологии рассматривали под микроскопом строение клеток кожицы лука — классического школьного препарата. Если помните, клетки на этом препарате были прозрачными. Таково большинство клеток, и без специальной окраски увидеть можно немного: в основном, клеточную оболочку и ядро. Но существует множество различных способов окраски клеток, которые открывают перед нами невероятный внутренний мир клетки: от классического окрашивания гематоксилин-эозином до методик, позволяющих наблюдать за клетками в сверхвысоком разрешении.
Чаще всего для окрашивания и изучения клеток используются флуоресцентные молекулы, которыми помечают нужные компоненты клетки. После этого клетки наблюдают с помощью конфокального или флуоресцентного микроскопа.
На фото представлено довольно простое окрашивание. Синим помечено ядро клетки, где хранится ДНК. Длина ДНК человека, по разным данным, составляет от 1,5 до 3 метров, но вся она упакована в небольшом объеме (диаметр ядра клетки на фотографии — около 20 микрометров). Это становится возможным благодаря особой многоуровневой компактной упаковке хроматина.
Окрашивать ядра клеток очень просто: флуоресцентные молекулы красителя DAPI прочно прикрепляются к ДНК, и ядро становится видимым.
Тот же принцип иcпользуется и для окраски актиновых филаментов (на фото красные). Токсин фаллоидин из яда бледной поганки (Amanita phalloides), меченый флуоресцентным красителем, охотно прикрепляется к актиновым филаментам. Белок актин, из которого состоят филаменты, — это каркас и «мышцы» клетки. Вместе с белком миозином он образует микрофиламенты, которые могут сокращаться, и с их помощью клетка движется (см. Actin, Myosin, and Cell Movement).
В отличие от большинства растительных клеток, окруженных плотной клеточной стенкой и неподвижных, животные клетки ползают и даже «бегают». Способность к миграции очень важна. Например, она играет роль в функционировании иммунных клеток, в восстановлении поврежденных мышечных волокон, но также и в патологических процессах: метастазы образуются как раз за счет способности раковых клеток к миграции.
Зеленое на фото — микротрубочки, образованные димерами белка тубулина. Это транспортная система клетки: по ним, как по рельсам, переносятся питательные вещества и строительные материалы. В роли «грузчиков» выступают специальные моторные белки — динеины и кинезины. Динеины транспортируют грузы в направлении от клеточной мембраны к ядру, кинезины — наоборот, от центра клетки к периферии. Кроме транспортной функции микротрубочки участвуют в делении и миграции клеток, а также играют важнейшую роль в функционировании нервных клеток.
Окраска микротрубочек отличается от окраски ДНК или актина — для нее используется техника иммунофлуоресценции. Первичное антитело «узнает» белок тубулин, из которого состоят микротрубочки, а вторичное, несущее флуоресцентную молекулу, прикрепляется к первичному. С помощью данной техники можно визуализировать самые разные белки в клетке.
В знаменитой анимации The Inner Life of the Cell («Внутренняя жизнь клетки»), созданной в 2006 году командой из Гарвардского университета совместно с XVIVO, можно посмотреть на актиновые филаменты, на микротрубочки, на гордо шагающий по ним кинезин и на многое другое. Вот это видео с комментариями на русском языке:
Снимок цитосклета сделан с помощью конфокального микроскопа Zeiss LSM 700. Фото © Варвары Грибовой.
См. также об окрашивании клеток:
1) Разноцветные нейроны, «Элементы», 5.05.2016.
2) Микроглия через конфокальный микроскоп, «Элементы», 15.06.2016.
3) Светящиеся данио-рерио, «Элементы», 26.07.2016.
Варвара Грибова
Источник: elementy.ru