Хлоропласты строение и функции кратко
10
Федеральное Агентство науки и образования.
Сибирский Федеральный Университет.
Институт Фундаментальной Биологии и Биотехнологии.
Кафедра биотехнологии.
РЕФЕРАТ
На тему: Строение и функции хлоропластов.
Геном пластид. Пропластиды.
Выполнила: студентка
31гр. Осипова И.В.
Проверила:
доцент кафедры
биотехнологии
д.б.н. Голованова Т.И.
Красноярск,2008г.
Содержание.
Содержание. 2
Введение. 3
Хлоропласты 4
Функции хлоропластов. 6
Геном пластид 9
Пропластиды 13
Заключение. 15
Литература. 16
Введение.
Пластиды – это мембранные органоиды, встречающиеся у фотосинтезирующих
эукариотических организмов (высшие растения, низшие водоросли, некоторые
одноклеточные организмы). Пластиды окружены двумя мембранами, в их
матриксе имеется собственная геномная система, функции пластид связаны с
энергообеспечением клетки, идущим на нужды фотосинтеза.
Всем пластидам свойственен ряд общих черт. Они имеют собственный геном,
одинаковый у всех представителей одного вида растений, собственную
белоксинтезирующую систему; от цитозоля пластиды отделены двумя
мембранами – наружной и внутренней. Для некоторых фототрофных организмов
число пластидных мембран может быть больше. Например, пластиды эвглен и
динфлагеллят окружены тремя, а у золотистых, бурых, желто-зелёных и
диатомовых водорослей они имеют четыре мембраны. Это связано с
происхождением пластид. Считается, что симбиотический процесс,
результатом которого стало формирование пластид, в процессе эволюции
происходило неоднократно (как минимум, трижды).
У высших растений найден целый набор различных пластид (хлоропласт,
лейкопласт, амилопласт, хромопласт), представляющих собой ряд взаимных
превращений одного вида пластиды в другой. Основной структурой, которая
осуществляет фотосинтетические процессы, является хлоропласт.
Хлоропласты.
Хлоропласты – это структуры, в которых происходят фотосинтетические
процессы, приводящие в конечном итоге к связыванию углекислоты, к
выделению кислорода и синтезу сахаров.Структуры удлиненной формы с
шириной 2-4 мкм и протяженностью 5-10 мкм. У зеленых водорослей
встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50
мкм.
у зеленых водорослей может быть по одному хлоропласту на клетку. Обычно
на клетку высших растений приходится в среднем 10-30 хлоропластов.
Встречаются клетки с огромным количеством хлоропластов. Например, в
гигантских клетках палисадной ткани махорки обнаружено около 1000
хлоропластов.
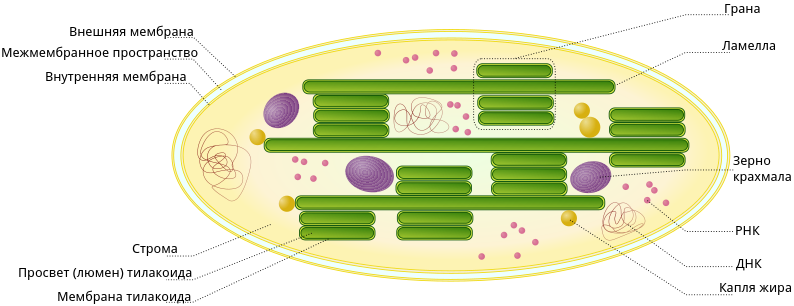
Хлоропласты представляют собой структуры, ограниченные двумя мембранами
– внутренней и внешней. Внешняя мембрана, как и внутренняя, имеет
толщину около 7 мкм, они отделены друг от друга межмембранным
пространством около 20-30 нм. Внутренняя мембрана хлоропластов отделяет
строму пластиды, аналогичную матриксу митохондрий. В строме зрелого
хлоропласта высших растений видны два типа внутренних мембран. Это –
мембраны, образующие плоские, протяженные ламеллы стромы, и мембраны
тилакоидов, плоских дисковидных вакуолей или мешков.
Ламеллы стромы (толщиной около 20 мкм) представляют собой плоские полые
мешки или же имеют вид сети из разветвленных и связанных друг с другом
каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри
хлоропласта лежат параллельно друг другу и не образуют связей между
собой.
Кроме мембран стромы в хлоропластах обнаруживаются мембранные тилакоиды.
Это плоские замкнутые мембранные мешки, имеющие форму диска. Величина
межмембранного пространства у них также около 20-30 нм. Такие тилакоиды
образуют стопки наподобие столбика монет, называемые гранами.
Число тилакоидов на одну грану очень варьирует: от нескольких штук до 50
и более. Размер таких стопок может достигать 0,5 мкм, поэтому граны
видны в некоторых объектах в световом микроскопе. Количество гран в
хлоропластах высших растений может достигать 40-60. Тилакоиды в гране
сближены друг с другом так, что внешние слои их мембран тесно
соединяются; в месте соединения мембран тилакоидов образуется плотный
слой толщиной около 2 нм. В состав граны кроме замкнутых камер
тилакоидов обычно входят и участки ламелл, которые в местах контакта их
мембран с мембранами тилакоидов тоже образуют плотные 2-нм слои. Ламеллы
стромы, таким образом, как бы связывают между собой отдельные граны
хлоропласта. Однако полости камер тилакоидов всезда замкнуты и не
переходят в камеры межмембранного пространства ламелл стромы. Ламеллы
стромы и мембраны тилакоидов образуются путем отделения от внутренней
мембраны при начальных этапах развития пластид.
В матриксе (строме) хлоропластов обнаруживаются молекулы ДНК, рибосомы;
там же происходит первичное отложение запасного полисахарида, крахмала,
в виде крахмальных зерен.
Характерным для хлоропластов является наличие в них пигментов,
хлорофиллов, которые и придают окраску зеленым растениям. При помощи
хлорофилла зеленые растения поглощают энергию солнечного света и
превращают ее в химическую.
В хлоропластах содержатся различные пигменты. В зависимости от вида
растений это:
хлорофилл:
– хлорофилл А (сине-зеленый) – 70 % (у высших растений и зеленых
водорослей);
– хлорофилл В (желто-зеленый) – 30 % (там же);
– хлорофилл С, D и E встречается реже – у других групп водорослей;
каротиноиды:
– оранжево-красные каротины (углеводороды);
– желтые (реже красные) ксантофиллы (окисленные каротины). Благодаря
ксантофиллу фикоксантину хлоропласты бурых водорослей (феопласты)
окрашены в коричневый цвет;
фикобилипротеиды, содержащиеся в родопластах (хлоропластах красных и
сине-зеленых водорослей):
– голубой фикоцианин;
– красный фикоэритрин.
Функции хлоропластов.
Хлоропласты- это структуры, в которых осуществляются фотосинтетические
процессы, приводящие в конечном итоге к связыванию углекислоты, к
выделению кислорода и синтезу сахаров.
Характерным для хлоропластов является наличие в них пигментов
хлорофиллов, которые и придают окраску зеленым растениям. При помощи
хлорофилла зеленые растения поглощают энергию солнечного света и
превращают её в химическую. Поглощение света с определённой длиной волны
приводит к изменению в структуре молекулы хлорофилла, при этом она
переходит в возбуждённое, активированное состояние. Освобождающаяся
энергия активированного хлорофилла через ряд промежуточных этапов
передаётся определённым синтетическим процессам, приводящим к синтезу
АТФ и к восстановлению акцептора электронов НАДФН
(никотинамидадениндинуклеотид-фосфат) до НАДФ*Н, которые тратятся в
реакции связывания СО2 и синтезе сахаров.
Суммарная реакция фотосинтеза может быть выражена следующим образом:
nCO2+ nH2O—(CH2O)n+nO2
Таким образом, главный итоговый процесс здесь- связывание двуокиси
углерода с использованием воды для образования различных углеводов и для
выделения кислорода. Молекула кислорода, который выделяется в процессе
фотосинтеза у растений, образуется за счёт гидролиза молекулы воды.
Следовательно, процесс включает в себя процесс гидролиза воды, которая
служит одним из источников электронов или атомов водорода. Биохимические
исследования показали, что процесс фотосинтеза представляет собой
сложную цепь событий, заключающую в себе 2 стадии: световую и темновую.
Первая, протекающая только на свету, связанная с поглощением света
хлорофиллами и с проведением фотохимической реакции (реакция Хилла). Во
второй фазе, которая может идти в темноте, происходят фиксация и
восстановление СО2, приводящие к синтезу углеводов.
В результате световой фазы осуществляются фотофосфорилирование, синтез
АТФ из АДФ и фосфата с использованием цепи переноса электронов, а также
восстановление кофермента НАДФ в НАДФН, происходящее при гидролизе и
ионизации воды.
этой фазе фотосинтеза энергия солнечного света
возбуждает электроны в молекулах хлорофилла, которые расположены в
мембранах тилакоидов. Эти возбуждённые электроны переносятся по
компонентам окислительной цепи в тилакоидной мембране, подобно тому как
электроны транспортируются по дыхательной цепи в мембране митохондрий.
Энергия, освобождающаяся при таком переносе электронов, используется для
перекачивания протонов через тилакоидную мембрану внутрь тилакоидов, что
приводит к возрастанию разности потенциалов между стромой и
пространством внутри тилакоида. Как и в мембранах крист митохондрий, в
мембранах тилакоидов встроены молекулярные комплексы АТФ-синтетазы,
которые начинают затем транспортировать протоны обратно в матрикс
хлоропласта, или строму, и параллельно этому фосфорилировать АДФ, т. е.
синтезировать АТФ.
Таким образом, в результате световой фазы происходят синтез АТФ и
восстановление НАДФ, которые затем используются при восстановлении СО2,
в синтезе углеводов уже в темновой фазе фотосинтеза.
В темновой (не зависящей от потока фотонов) стадии фотосинтеза за счет
восстановленного НАДФ и энергии АТФ осуществляется связывание
атмосферного СО2, что приводит к образованию углеводов. Процесс фиксации
СО2 и образования углеводов состоит из многих этапов, в которых
участвует большое число ферментов (цикл Кальвина). Биохимическими
исследованиями показано, что ферменты, участвующие в темновых реакциях,
содержатся в водорастворимой фракции хлоропластов, содержащей компоненты
матрикса-стромы этих пластид.
Процесс восстановления СО2 начинается с его присоединения к
рибулозодифосфату-углеводу, состоящему из пяти атомов углерода, с
образованием короткоживущего С6-соединения, которое сразу распадается на
два С3-соединения, на две молекулы глицерид-3-фосфата.
Именно на этом этапе при карбоксилировании рибулозодифосфата и роисходит
связывание СО2. Дальнейшие реакции превращения глицерид-3-фосфата
приводят к синтезу различных гесоз и пентоз, к регенерации
рибулозодифосфата и к его новому вовлечению в цикл реакций связывания
СО2. В конечном счёте в хлоропласте из шести молекул СО2 образуется одна
молекула гексозы. Для этого процесса требуется 12 молекул НАДФН и 18
молекул АТФ, поступающих из световых реакций фотосинтеза. Образовавшийся
в результате темновой реакции фруктоза-6-фосфат даёт начало сахарам,
полисахаридам (крахмал) и галактолипидам. В строме хлоропластов, кроме
того, из части глицерид-3-фосфата образуются жирные кислоты,
аминокислоты и крахмал. Синтез сахарозы завершается в цитоплазме.
В строме хлоропластов происходит восстановление нитрттов до аммиака за
счёт энергии электронов, активированных светом; в растениях этот аммиак
служит источником азота при синтезе аминокислот и нуклеотидов.
Геном пластид.
Подобно митохондриям, хлоропласты имеют собственную генетическую
систему, обеспечивающую синтез ряда белков внутри самих пластид. В
матриксе хлоропластов обнаруживаются ДНК, разные РНК и рибосомы.
Оказалось, что ДНК хлоропластов резко отличается от ДНК ядра. Она
представлена циклическими молекулами длиной до 40-60 мкм, имеющими
молекулярный вес 0,8-1,3х108 дальтон. В одном хлоропласте может быть
множество копий ДНК. Так, в индивидуальном хлоропласте кукурузы
присутствует 20-40 копий молекул ДНК. Длительность цикла и скорость
репликации ядерной и хлоропластной ДНК, как было показано на клетках
зеленых водорослей, не совпадают. ДНК хлоропластов не состоит в
комплексе с гистонами. Все эти характеристики ДНК хлоропластов близки к
характеристикам ДНК прокариотических клеток. Более того, сходство ДНК
хлоропластов и бактерий подкрепляется еще и тем, что основные
регуляторные последовательности транскрипции (промоторы, терминаторы) у
них одинаковы. На ДНК хлоропластов синтезируются все виды РНК
(информационная, трансферная, рибосомная). ДНК хлоропластов кодирует
рРНК, входящую в состав рибосом этих пластид, которые относятся к
прокариотическому 70S типу (содержат 16S и 23S рРНК). Рибосомы
хлоропластов чувствительны к антибиотику хлорамфениколу, подавляющему
синтез белка у прокариотических клеток.
Так же как в случае хлоропластов мы вновь сталкиваемся с существованием
особой системы синтеза белка, отличной от таковой в клетке.
Эти открытия вновь пробудили интерес к теории симбиотического
происхождения хлоропластов. Идея о том, что хлоропласты возникли за счет
объединения клеток-гетеротрофов с прокариотическими синезелеными
водорослями, высказанная на рубеже XIX и XX вв. (А.С. Фоминцин,
К.С.Мережковский) вновь находит свое подтверждение. В пользу этой теории
говорит удивительное сходство в строении хлоропластов и синезеленых
водорослей, сходство с основными их функциональными особенностями, и в
первую очередь со способностью к фотосинтетическим процессам.
Известны многочисленные факты истинного эндосимбиоза синезеленых
водорослей с клетками низших растений и простейших, где они
функционируют и снабжают клетку-хозяина продуктами фотосинтеза.
Оказалось, что выделенные хлоропласты могут также отбираться некоторыми
клетками и использоваться ими как эндосимбионты. У многих беспозвоночных
(коловратки, моллюски), питающихся высшими водорослями, которые они
переваривают, интактные хлоропласты оказываются внутри клеток
пищеварительных желез. Так, у некоторых растительноядных моллюсков в
клетках найдены интактные хлоропласты с функционирующими
фотосинтетическими системами, за активностью которых следили по
включению С14О2.
Как оказалось, хлоропласты могут быть введены в цитоплазму клеток
культуры фибробластов мыши путем пиноцитоза.
нако они не подвергались
атаке гидролаз. Такие клетки, включившие зеленые хлоропласты, могли
делиться в течение пяти генераций, а хлоропласты при этом оставались
интактными и проводили фотосинтетические реакции. Были предприняты
попытки культивировать хлоропласты в искусственных средах: хлоропласты
могли фотосинтезировать, в них шел синтез РНК, они оставались интактными
100 ч, у них даже в течение 24 ч наблюдались деления. Но затем
происходило падение активности хлоропластов, и они погибали.
Эти наблюдения и целый ряд биохимических работ показали, что те черты
автономии, которыми обладают хлоропласты, еще недостаточны для
длительного поддержания их функций и тем более для их воспроизведения.
В последнее время удалось полностью расшифровать всю последовательность
нуклеотидов в составе циклической молекулы ДНК хлоропластов высших
растений. Эта ДНК может кодировать до 120 генов, среди них: гены 4
рибосомных РНК, 20 рибосомных белков хлоропластов, гены некоторых
субъединиц РНК-полимеразы хлоропластов, несколько белков I и II
фотосистем, 9 из 12 субъединиц АТФ-синтетазы, части белков комплексов
цепи переноса электронов, одной из субъединиц
рибулозодифосфат-карбоксилазы (ключевой фермент связывания СО2), 30
молекул тРНК и еще 40 пока неизвестных белков. Интересно, что сходный
набор генов в ДНК хлоропластов обнаружен у таких далеко отстоящих
представителей высших растений как табак и печеночный мох.
Основная же масса белков хлоропластов контролируется ядерным геномом.
Оказалось, что ряд важнейших белков, ферментов, а соответственно и
метаболические процессы хлоропластов находятся под генетическим
контролем ядра. Так, клеточное ядро контролирует отдельные этапы синтеза
хлорофилла, каротиноидов, липидов, крахмала. Под ядерным контролем
находятся многие энзимы темновой стадии фотосинтеза и другие ферменты, в
том числе некоторые компоненты цепи транспорта электронов. Ядерные гены
кодируют ДНК-полимеразу и аминоацил-тРНК-синтетазу хлоропластов. Под
контролем ядерных генов находится большая часть рибосомных белков. Все
эти данные заставляют говорить о хлоропластах, так же как и о
митохондриях, как о структурах с ограниченной автономией.
Транспорт белков из цитоплазмы в пластиды происходит в принципе сходно с
таковым у митохондрий. Здесь также в местах сближения внешней и
внутренней мембран хлоропласта располагаются каналообразующие
интегральные белки, которые узнают сигнальные последовательности
хлоропластных белков, синтезированных в цитоплазме, и транспортируют их
в матрикс-строму. Из стромы импортируемые белки согласно дополнительным
сигнальным последовательностям могут включаться в мембраны пластиды
(тилакоиды, ламеллы стромы, внешняя и внутренняя мембраны) или
локализоваться в строме, входя в состав рибосом, ферментных комплексов
цикла Кальвина и др.
Удивительное сходство структуры и энергетических процессов у бактерий и
митохондрий, с одной стороны, и у синезеленых водорослей и хлоропластов
– с другой, служит веским аргументом в пользу теории симбиотического
происхождения этих органелл. Согласно этой теории, возникновение
эукариотической клетки прошло через несколько этапов симбиоза с другими
клетками. На первой стадии клетки типа анаэробных гетеротрофных бактерий
включили в себя аэробные бактерии, превратившиеся в митохондрии.
Параллельно этому в клетке-хозяине прокариотический генофор формируется
в обособленное от цитоплазмы ядро. Так могли возникнуть гетеротрофные
эукариотические клетки. Повторные эндосимбиотические взаимоотношения
между первичными эукариотическими клетками и синезелеными водорослями
привели к появлению в них структур типа хлоропластов, позволяющих
клеткам осуществлять автосинтетические процессы и не зависеть от наличия
органических субстратов. В процессе становления такой составной живой
системы часть генетической информации митохондрий и пластид могла
изменяться, перенестись в ядро. Так, например две трети из 60 рибосомных
белков хлоропластов кодируется в ядре и синтезируются в цитоплазме, а
потом встраивается в рибосомы хлоропластов, имеющие все свойства
прокариотических рибосом. Такое перемещение большой части
прокариотических генов в ядро привело к тому, что эти клеточные
органеллы, сохранив часть былой автономии, попали под контроль
клеточного ядра, определяющего в большей степени все главные клеточные
функции.
Пропластиды.
При нормальном освещении пропластиды превращаются в хлоропласты. Сначала
они растут, при этом происходит образование продольно расположенных
мембранных складок от внутренней мембраны. Одни из них простираются по
всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы
тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых
хлоропластов. Несколько иначе развитие пластид происходит в темноте. У
этиолированных проростков происходит в начале увеличение объема пластид,
этиопластов, но система внутренних мембран не строит ламеллярные
структуры, а образует массу мелких пузырьков, которые скапливаютсяя в
отдельные зоны и даже могут формировать сложные решетчатые структуры
(проламеллярные тела). В мембранах этиопластов содержится
протохлорофилл, предшественник хлорофилла желтого цвета. Под действие
света из этиопластов образуются хлоропласты, протохлорофилл превращается
в хлорофилл, происходит синтез новых мембран, фотосинтетических
ферментов и компонентов цепи переноса электронов.
При освещении клеток мембранные пузырьки и трубочки быстро
реорганизуются, из них развивается полная система ламелл и тилакоидов,
характерная для нормального хлоропласта.
Лейкопласты отличаются от хлоропластов отсутствием развитой ламеллярной
системы. Встречаются они в клетках запасающих тканей. Из-за их
неопределенной морфологии лейкопласты трудно отличить от пропластид, а
иногда и от митохондрий. Они, как и пропластиды, бедны ламеллами, но тем
не менее способны к образованию под влиянием света нормальных
тилакоидных структур и к приобретению зеленой окраски. В темноте
лейкопласты могут накапливать в проламеллярных телах различные запасные
вещества, а в строме лейкопластов откладываются зерна вторичного
крахмала. Если в хлоропластах происходит отложение так называемого
транзиторного крахмала, который присутствует здесь лишь во время
ассимиляции СО2, то в лейкопластах может происходить истинное запасание
крахмала. В некоторых тканях (эндосперм злаков, корневища и клубни)
накопление крахмала в лейкопластах приводит к образованию амилопластов,
сплошь заполненных гранулами запасного крахмала, расположенных в строме
пластиды.
Другой формой пластид у высших растений является хромопласт,
окрашивающийся обычно в желтый свет в результате накопления в нем
каротиноидов. Хромопласты образуются из хлоропластов и значительно реже
их лейкопластов (например, в корне моркови). Процесс обесцвечивания и
изменения хлоропластов легко наблюдать при развитии лепестков или при
созревании плодов. При этом в пластидах могут накапливаться окрашенные в
желтый цвет капельки (глобулы) или в них появляются тела в форме
кристаллов. Эти процессы сопряжены с постепенным уменьшением числа
мембран в пластиде, с исчезновением хлорофилла и крахмала. Процесс
образования окрашенных глобул объясняется тем, что при разрушении ламелл
хлоропластов выделяются липидные капли, в которых хорошо растворяются
различные пигменты (например, каротиноиды). Таким образом, хромопласты
представляют собой дегенерирующие формы пластид, подвернутые
липофанерозу – распаду липопротедных комплексов.
Заключение.
Пластиды. Пластиды – особые органоиды растительных клеток, в которых
осуществляется синтез различных веществ, и в первую очередь фотосинтез.
В цитоплазме клеток высших растений имеется три основных типа пластид:
1) зеленые пластиды – хлоропласты; 2) окрашенные в красный, оранжевый и
другие цвета хромопласты; 3) бесцветные пластиды – лейкопласты. Все эти
типы пластид могут переходить один в другой. У низших растений, например
у водорослей, известен один тип пластид – хроматофоры. Процесс
фотосинтеза у
высших растений протекает в хлоропластах, которые, как правило,
развиваются только на свету.
Снаружи хлоропласты ограничены двумя мембранами: наружной и внутренней.
В состав хлоропластов высших растений, по данным электронной
микроскопии, входит большое количество гран, расположенных группами.
Каждая
грана состоит из многочисленных круглых пластин, имеющих форму плоских
мешочков, образованных двойной мембраной и сложенных друг с другом
наподобие столбика монет. Граны соединяются между собой посредством
особых пластин или трубочек, расположенных в строме хлоропласта и
образующих
единую систему. Зеленый пигмент хлоропластов содержат только граны;
строма их бесцветна.
Хлоропласты одних растений содержат лишь несколько гран, других – до
пятидесяти и больше.
У зеленых водорослей процессы фотосинтеза осуществляются в хроматофорах,
которые не содержат гран, и продукты первичного синтеза –различные
углеводы – часто откладываются вокруг особых клеточных структур,
называемых пиреноидами.
Окраска хлоропластов зависит не только от хлорофилла, в них могут
содержаться и другие пигменты, например каротин и каротиноиды,
окрашенные в разные цвета – от желтого до красного и коричневого, а
также фикобилины. К последним относится фикоцианин и фикоэритрин красных
и сине-зеленых водорослей.Пластиды развиваются из особых клеточных
структур, носящих название пропластид. Пропластиды – это бесцветные
образования, внешне похожие на митохондрии, но отличающиеся от них более
крупными размерами и тем, что всегда имеют удлиненную форму. Снаружи
пластиды ограничены двойной мембраной, небольшое количество мембран
находится также в их внутренней части. Пластиды размножаются путем
деления, и контроль над этим процессом осуществляется, по-видимому, ДНК,
содержащейся в них же. При делении происходит перетяжка пластиды, но
разделение пластид может происходить и путем образования перегородки.
Способность пластид к делению обеспечивает их непрерывность в ряду
клеточных поколений. При половом и бесполом размножении растений
происходит передача пластид дочерним организмам.
Подобно митохондриям, хлоропласты имеют собственную генетическую
систему, обеспечивающую синтез ряда белков внутри самих пластид. В
матриксе хлоропластов обнаруживаются ДНК, разные РНК и рибосомы. ДНК
хлоропластов резко отличается от ДНК ядра.
Литература.
1) Ю.С.Ченцов. Введение в клеточную биологию./Ю.С. Ченцов.-М.:ИКЦ
«Академкнига»,2005-495с.:ил.
2) Физиология растений: Учебник для студ.вузов/ Н.Д.Алёхина,
Ю.В.Балнокин, В.Ф. Гавриленко, Т.В.Жигалова, Н.Р. Мейчик, А.М.Носов,
О.Г.Полесская, Е.В.харитонашвили; Под ред. И.П.Ермакова.-М.:изд.центр
«Академия»,2005.-640с.
Источник: ukrreferat.com
Описание клеточных элементов
Какие компоненты клеток именуются пластидами. Это структурные органоиды клетки, имеющие сложное строение и функции, важные для жизни растительных организмов.
 Какое имеют пластиды строение под микроскопом рассмотреть сложно, благодаря плотной оболочке они не просвечиваются.
Какое имеют пластиды строение под микроскопом рассмотреть сложно, благодаря плотной оболочке они не просвечиваются.
Однако, ученым удалось выяснить, что этот органоид имеет две мембраны, внутри заполнен стромой, аналогичной цитоплазме жидкостью.
Складки внутренней мембраны, уложенные стопочками, образуют граны, которые могут соединяться между собой.
Также внутри присутствуют рибосомы, липидные капли, зерна крахмала. Еще у пластид, особенно у хлоропластов, имеются свои молекулы ДНК.
Это интересно! Сходство, отличия и признаки: голосеменные и покрытосеменные растения
Классификация
Разделяются на три группы по цвету и выполняемым функциям:
- хлоропласты,
- хромопласты,
- лейкопласты.
Хлоропласты
Наиболее глубоко изучены, имеют зеленую окраску. Содержаться в листьях растений, иногда в стеблях, плодах и даже корнях. По внешнему виду похожи на округлые зернышки размером 4-10 микрометров. Малый размер и большое количество значительно увеличивает площадь рабочей поверхности.
Могут отличаться по цвету, это зависит от вида и концентрации содержащегося в них пигмента. Основной пигмент- хлорофилл, также присутствуют ксантофилл и каротин. В природе существует 4 вида хлорофилла, обозначаемых латинскими буквами: а, b, с, е. Первые два типа содержат клетки высших растений и зеленых водорослей, у диатомовых присутствуют только разновидности — а и с.
Хлоропласты выполняют важную функцию: внутри них происходит процесс фотосинтеза — преобразование солнечного света в энергию химических связей формирующихся углеводов. При этом они могут двигаться вместе с током цитоплазмы или активно передвигаться сами. Так, при слабом освещении они скапливаются у стенок клетки с большим количеством света и поворачиваются к нему большей площадью, а при очень активном освещении, наоборот, встают ребром.
Хромопласты
Приходят на смену разрушенным хлоропластам, бывают желтого, красного и оранжевого оттенков. Цветная окраска формируется благодаря содержанию каротиноидов.
Данные органоиды содержаться в листья, цветах и плодах растений. По форме могут быть округлыми, прямоугольными или даже игольчатыми. Строение аналогично хлоропластам.
Основная функция – придание окраски цветам и плодам, что позволяет привлечь насекомых- опылителей и животных, которые поедают плоды и тем самым способствуют распространению семян растения.
Лейкопласты
Данные пластиды имеют отличия в строении и функциях. Основная задача – запасать питательные вещества впрок, поэтому находятся они преимущественно в плодах, но также могут быть в утолщенных и мясистых частях растения:
- клубнях,
- корневищах,
- корнеплодах,
- луковицах и других.
Бесцветная окраска не позволяет выделить их в структуре клетки, однако лейкопласты легко разглядеть при добавлении небольшого количества йода, который, взаимодействуя с крахмалом, окрашивает их в синий цвет.
Форма близка к округлой, при этом внутри плохо развита система мембран. Отсутствие складок мембран помогает органоиду при запасании веществ.
Крахмальные зерна увеличиваются в размерах и легко разрушают внутренние мембраны пластиды, как-бы растягивая ее. Это позволяет накопить больше углеводов.
В отличие от других пластид, содержат молекулу ДНК в оформленном ядре. При этом, накапливая хлорофилл, лейкопласты могут превращаться в хлоропласты.
Определяя, какую функцию выполняют лейкопласты, нужно отметить их специализацию, поскольку существует несколько типов, запасающих определенные вид органического вещества:
- амилопласты накапливают крахмал;
- олеопласты производят и запасают жиры, при этом последние могут запасаться и в других частях клеток;
- протеинопласты «берегут» белки.
Помимо накопления, могут выполнять функцию расщепления веществ, для чего существуют ферменты, которые активизируются, когда возникает дефицит энергии или строительного материала.
В такой ситуации ферменты начинают расщеплять запасенные жиры и углеводы до мономеров, чтобы клетка получила необходимую энергию.
Все разновидности пластид, не смотря на особенности строения, обладают способностью превращаться друг в друга. Так, лейкопласты могут преобразоваться в хлоропласты, этот процесс мы видим при позеленении клубней картофеля.
В то же время, по осени хлоропласты превращаются в хромопласты, в результате чего листья желтеют. Каждая клетка содержит только один вид пластид.
Это интересно! Кто такие эукариоты и прокариоты: сравнительная характеристика клеток разных царств
Происхождение
Теорий происхождения множество, наиболее обоснованными среди них являются две:
- симбиоза,
- поглощения.
Первая рассматривает образование клетки как процесс симбиоза, происходящего в несколько ступеней. В его ходе гетеротрофные и автотрофные бактерии объединяются, получая взаимную выгоду.
Вторая теория рассматривает образование клетки через поглощение более крупными организмами мелких. Однако, при этом не происходит их переваривание, они встраиваются в структуру бактерии, выполняя свою функцию внутри нее. Такое строение оказалось удобным и дало организмам преимущество перед другими.
Виды пластидов в растительной клетке
Пластиды — их функции в клетке и типы
Вывод
Пластиды в растительных клетках – это своеобразная «фабрика», где осуществляется производство, связанное с токсичными промежуточными веществами, высокой энергией и процессами преобразования свободных радикалов.
Источник: uchim.guru
фотосинтез протекает в специализированных органеллах клеток — хлоропластах. Хлоропласты высших растений имеют форму двояковыпуклой линзы (диска), которая наиболее удобна для поглощения солнечных лучей. Их размеры, количество, расположение полностью отвечают назначению: как можно эффективнее поглощать солнечную энергию, как можно полнее усваивать углерод. Установлено, что количество хлоропластов в клетке измеряется десятками. Это обеспечивает высокое содержание этих органелл на единицу поверхности листа. Так, на 1 мм2 листа фасоли приходится 283 тыс. хлоропластов, у подсолнечника — 465 тыс. Диаметр хлоропластов в среднем 0,5-2 мкм.
Строение хлоропласта весьма сложное. Подобно ядру и митохондриям хлоропласт окружен оболочкой, состоящей из двух липопротеидных мембран. Внутреннюю среду представляет относительно однородная субстанция — матрикс или строма, которую пронизывают мембраны — ламеллы (рис.). Ламеллы, соединенные друг с другом, образуют пузырьки — тилакоиды. Плотно прилегая друг к другу, тилакоиды образуют граны, которые различают даже под световым микроскопом. В свою очередь, граны в одном или нескольких местах объединены друг с другом с помощью межгранных тяжей — тилакоидов стромы.
Свойства хлоропластов: способны измененять ориентацию и перемещаться. Например, под влиянием яркого света хлоропласты поворачиваются узкой стороной диска к падающим лучам и перемещаются на боковые стенки клеток. Хлоропласты передвигаются в направлении более высокой концентрации СО2 в клетке. Днем они обычно выстраиваются вдоль стенок, ночью опускаются на дно клетки.
Химический состав хлоропластов: воды — 75 %; 75-80 % общего количества сухих веществ составляют орг. соединения, 20-25 % -минеральные.
Структурной основой хлоропластов являются белки (50-55 % сухой массы), половина из них составляют водорастворимые белки. Такое высокое содержание белков объясняется их многообразными функциями в составе хлоропластов (структурные белки мембран, белки-ферменты, транспортные белки, сократительные белки, реценторные).
Важнейшей составной частью хлоропластов являются липиды, (30-40% сух. м.). Липиды хлоропластов представлены тремя группами соединений.
-
Структурные компоненты мембран, которые представлены амфипатическими липоидами и отличаются высоким содержанием (более 50%) галактолипидов и сульфолипидов. Фосфолипидный состав характеризуется отсутствием фосфатидилэтаноламина и высоким содержанием фосфатидилглицерина (более 20 %). Свыше 60 % состава ЖК приходится на линолевую кислоту.
-
Фотосинтетическне пигменты хлоропластов — гидрофобные вв-а, относящиеся к липоидам (в клеточном соке — водорастворимые пигменты). Высшие растения содержат 2 формы зеленых пигментов: хлорофилл а и хлорофилл b и 2 формы желтых пигментов: каротины и ксантофиллы (каротиноиды). Хлорофиллы выполняет роль фотосенсибилизаторов, другие пигменты расширяют спектр действия фотосинтеза за счет более полного поглощения ФАР. Каротиноиды защищают хлорофилл от фотоокисления, участвуют в транспорте водорода, образующегося при фотолизе воды.
-
Жирорастворимые витамины — эргостерол (провитамин Д), витамины Е, К — сосредоточены практически целиком в хлоропластах, где участвуют в преобразовании световой энергии в химическую. В цитозоле клеток листа в основном находятся водорастворимые витамины. Так, у шпината содержание аскорбиновой кислоты в хлоропластах в 4-5 раз меньше, чем в листьях.
В хлоропластах листьев присутствует значительное количество РНК и ДНК. НК составляют примерно 1 % сухой массы хлоропластов (РНК — 0.75 %, ДНК — 0,01-0,02 %). Геном хлоропластов представлен кольцевой молекулой ДНК длиной 40 мкм с молекулярной массой 108, кодирующей 100-150 белков средних размеров. Рибосомы хлоропластов составляют от 20 до 50 % общей популяции рибосом клетки. Т.о., хлоропласты имеют собственную белоксинтезирующую систему. Однако для нормального функционирования хлоропластов необходимо взаимодеЯствие ядерного и хлоропластного геномов. Ключевой фермент фотосинтеза РДФ-карбоксилаза синтезируется под двойным контролем-ДНК ядра и хлоропласта.
Углеводы не являются конституционными веществами хлоропласта. Представлены фосфорными эфирами сахаров и продуктами фотосинтеза. Поэтому содержание углеводов в хлоропластах значительно колеблется (от 5 до 50 %). В активно функционирующих хлоропластах углеводы обычно не накапливаются, происходит их быстрый отток. При уменьшении потребности в продуктах фотосинтеза в хлоропластах образуются крупные крахмальные зерна. В этом случае содержание крахмала может возрасти до 50 % сухой массы и активность хлоропластов снизится.
Минеральные вещества. Сами хлоропласты составляют 25-30 % массы листа, но в них сосредоточено до 80 % Fe, 70-72 — Mg и Zn, 50 — Cu, 60 % Ca, содержащихся в тканях листа. Это объясняется высокой и разнообразной ферментативной активностью хлоропластов (входят с состав простетических групп и кофакторов). Mg входит в состав хлорофилла. Ca стабилизирует мембранные структуры хлоропластов.
Возникновение и развитие хлоропластов. Хлоропласты образуются в меристематических клетках из инициальных частиц или зачаточных пластид (рис.). Инициальная частица состоит из амебоидной стремы, окруженной двухмембранной оболочкой. По мере роста клетки инициалььные частицы увеличиваются в размере и приобретают форму двояковыпуклой линзы, в стреме появляются небольшие крахмальные зерна. Одновременно внутренняя мембрана начинает разрастаться, образуя складки (впячивания), от которых отшнуровываются пузырьки и трубочки. Такие образования называют пропластидами. Для дальнейшего их развития необходим свет. В темноте же формируются этиопласты, в которых образуется мембранная решетчатая структура — проламеллярное тело. На свету внутренние мембраны пропластид и этиопластов образуют гранильную систему. Одновременно с этим также на свету в граны встраиваются вновь образованные молекулы хлорофилла и других пигментов. Таким образом, структуры, которые подготавливаются к функционированию на свету, появляются и развиваются только при его наличии.
Наряду с хлоропластами имеется ряд других пластид, которые образуются либо непосредственно из пропластид, либо одна из другой путем взаимных превращений (рис.). К ним относятся накапливающие крахмал амилопласты (лейкопласты) и хромопласты, содержащие каротиноиды. В цветках и плодах хромопласты возникают на ранних стадиях развития пропластид. Хромопласты осенней листвы представляют собой продукты деградации хлоропластов, в которых в качестве структур — носителей каротнноидов выступают пластоглобулы.
Пигменты хлоропласта, участвующие в улавливании световой энергии, а также ферменты, необходимые для световой фазы фотосинтеза, вмонтированы в мембраны тилакоидов.
Ферменты, которые катализируют многочисленные реакции восстановительного цикла углеводов (темповой фазы фотосинтеза), а также разнообразные биосинтезы, в том числе биосинтезы белков, липидов, крахмала, присутствуют главным образом в строме, часть из них является периферическими белками ламелл.
Строение зрелых хлоропластов одинаково у всех высших растений, так же как в клетках разных органов одного растения (листьях, зеленеющих корнях, коре, плодах). В зависимости от функциональной нагрузки клеток, физиологического состояния хлоропластов, их возраста различают степень их внутренней структурированности: размеры, количество гран, связь между ними. Так, в замыкающих клетках устьиц основная функция хлоропластов — фоторегуляция устьичных движений. Хлоропласты не имеют строгой гранальной структуры, содержат крупные крахмальные зерна, набухшие тилакоиды, липофильные глобулы. Все это свидетельствует об их низкой энергетической нагрузке (эту функцию выполняют митохондрии). Другая картина наблюдается при изучении хлоропластов зеленых плодов томата. Наличие хорошо развитой гранулярной системы свидетельствует о высокой фукциональной нагрузке этих органелл и, вероятно, существенном вкладе фотосинтеза при формировании плодов.
Возрастные изменения: Молодые характеризуются ламеллярнои структурой, в таком состоянии хлоропласты способны размножаться путем деления. В зрелых хорошо выражена система гран. В стареющих происходит разрыв тилакоидов стромы, связь между гранами уменьшается, в дальнейшем наблюдаются распад хлорофилла и деструкция гран. В осенней листве деградация хлоропластов приводит к образованию хромопластов.
Структура хлоропластов лабильна и динамична, в ней отражаются все условия жизни растения. Большое влияние оказывает режим минерального питания растений. При недостатке N хлоропласты становятся в 1.5-2 раза мельче, дефицит P и S нарушает нормальную структуру ламелл и гран, одновременная нехватка N и Ca приводит к переполнению хлоропластов крахмалом из-за нарушения нормального оттока ассимилятов. При недостатке Ca нарушается структура наружной мембраны хлоропласта. Для поддержания структуры хлоропласта также необходим свет, в темноте идет постепенное разрушение тилакоидов гран и стремы.
Источник: StudFiles.net
Строение хлоропласта
В строении хлоропластов выделяют внешнюю и внутреннюю мембраны, межмембранное пространство, строму, тилакоиды, граны, ламеллы, люмен.
Тилакоид представляет собой ограниченное мембраной пространство в форме приплюснутого диска. Тилакоиды в хлоропластах объединяются в стопки, которые называют гранами. Граны связаны между собой удлиненными тилакоидами — ламеллами.
Полужидкое содержимое хлоропласта называется стромой. В ней находятся его ДНК и РНК, рибосомы, обеспечивающие полуавтономность органоида (см. Симбиогенез).
Также в строме находятся зерна крахмала. Они образуются при избытке углеводов, образовавшихся при фотосинтетической активности. Жировые капли обычно формируются из мембран разрушающихся тилакоидов.
Функции хлоропластов
Основная функция хлоропластов — это фотосинтез — синтез глюкозы из углекислого газа и воды за счет солнечной энергии, которая улавливается хлорофиллом. В качестве побочного продукта фотосинтеза выделяется кислород. Однако процесс этот сложный и многоступенчатый, при котором синтезируются и побочные продукты, использующиеся как в самом хлоропласте, так и в остальных частях клетки.
Основным фотосинтетическим пигментом является хлорофилл. Он существует в нескольких разных формах. Кроме хлорофилла в фотосинтезе принимают участие пигменты каротиноиды.
Пигменты локализованы в мембранах тилакоидов, здесь протекают световые реакции фотосинтеза. Кроме пигментов здесь присутствуют ферменты и переносчики электронов. Хлоропласты стараются расположиться в клетке так, чтобы их тилакоидные мембраны находились под прямым углом к солнечному свету.
Хлорофилл состоит из длинного углеводного кольца и порфириновой головки. Хвост гидрофобен и погружен в липидный слой мембран тилакоидов. Головка гидрофильна и обращена к строме. Энергия света поглощается именно головкой, что приводит к возбуждению электронов.
Электрон отделяется от молекулы хлорофилла, который после этого становится электроположительным, т. е. оказывается в окисленной форме. Электрон принимается переносчиком, которые передает его на другое вещество.
Разные виды хлорофилла отличаются между собой несколько различным спектром поглощения солнечного света. Больше всего в растениях хлорофилла А.
В строме хлоропласта происходят темновые реакции фотосинтеза. Здесь находятся ферменты цикла Кальвина и другие.
Источник: biology.su