Особенности строения хроматина
Хроматином называют сложную смесь веществ, из которых построены хромосомы эукариот. Основными компонентами хроматина являются ДНК, гистоны и негистоновые белки, образующие высокоупорядоченные в пространстве структуры. Соотношение ДНК и белка в хроматине составляет ~1:1, а основная масса белка хроматина представлена гистонами. Гистоны образуют семейство высококонсервативных основных белков, которые разделяются на пять больших классов, названных H1, H2A, H2B, H3 и H4. Размер полипептидных цепей гистонов лежит в пределах ~220 (H1) и 102 (H4) аминокислотных остатков. Гистон H1 сильно обогащен остатками Lys, для гистонов H2A и H2B характерно умеренное содержание Lys, полипептидные цепи гистонов H3 и H4 богаты Arg. Внутри каждого класса гистонов (за исключением H4) на основании аминокислотных последовательностей различают несколько субтипов этих белков. Такая множественность особенно характерна для гистонов класса H1 млекопитающих. В этом случае различают семь субтипов, названных H1.1–H1.5, H1o и H1t.

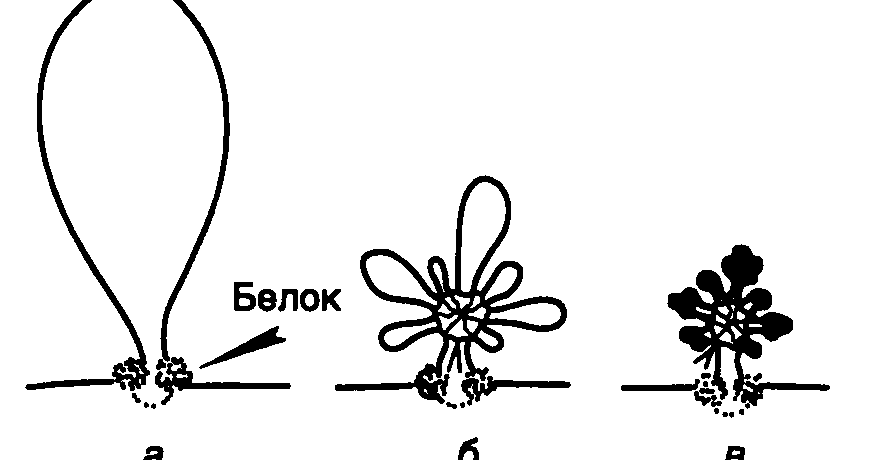
Рис. I.2. Схематическое изображение петельно-доменного уровня компактизации хроматина
а– фиксация петли хромомера на ядерном матриксе с помощью MAR/SAR-последовательностей и белков;б– «розетки», образованные из петли хромомера;в– конденсация петель «розеток» с участием нуклеосом и нуклеомеров
Важным результатом взаимодействия ДНК с белками в составе хроматина является ее компактизация. Суммарная длина ДНК, заключенной в ядре клеток человека, приближается к 1 м, тогда как средний диаметр ядра составляет 10 мкм. Длина молекулы ДНК, заключенной в одной хромосоме человека, в среднем равняется ~4 см. В то же время длина метафазной хромосомы составляет ~4 мкм. Следовательно, ДНК метафазных хромосом человека компактизована по длине, по крайней мере, в 104 раз. Степень компактизации ДНК в интерфазных ядрах значительно ниже и неравномерна в отдельных генетических локусах. С функциональной точки зрения различают эухроматин и гетерохроматин. Эухроматин характеризуется меньшей по сравнению с гетерохроматином компактизацией ДНК, и в нем главным образом локализуются активно экспрессирующиеся гены. В настоящее время широко распространено мнение о генетической инертности гетерохроматина. Поскольку его истинные функции сегодня нельзя считать установленными, эта точка зрения по мере накопления знаний о гетерохроматине может измениться. Уже сейчас в нем находят активно экспрессирующиеся гены.
Гетерохроматизация определенных участков хромосом часто сопровождается подавлением транскрипции имеющихся в них генов. В процесс гетерохроматизации могут быть вовлечены протяженные участки хромосом и даже целые хромосомы. В соответствии с этим считается, что регуляция транскрипции генов эукариот в основном происходит на двух уровнях. На первом из них компактизация или декомпактизация ДНК в хроматине может приводить к длительной инактивации или активации протяженных участков хромосом или даже целых хромосом в онтогенезе организма. Более тонкая регуляция транскрипции активированных участков хромосом достигается на втором уровне при участии негистоновых белков, включающих многочисленные факторы транскрипции.
Структурная организация хроматина и хромосом эукариот. Вопрос о структурной организации хроматина в интерфазных ядрах в настоящее время далек от своего разрешения. Это связано, прежде всего, со сложностью и динамичностью его структуры, которая легко меняется даже при незначительных экзогенных воздействиях. Большинство знаний о структуре хроматина было получено in vitro на препаратах фрагментированного хроматина, структура которого значительно отличается от таковой в нативных ядрах. В соответствии с распространенной точкой зрения различают три уровня структурной организации хроматина у эукариот: 1) нуклеосомная фибрилла; 2) соленоид, или нуклеомер; 3) петельно-доменная структура, включающая хромомеры.
Нуклеосомные фибриллы. В определенных условиях (при низкой ионной силе и в присутствии двухвалентных ионов металлов) в изолированном хроматине удается наблюдать регулярные структуры в виде протяженных фибрилл диаметром 10 нм, состоящих из нуклеосом. Эти фибриллярные структуры, в которых нуклеосомы расположены как бусы на нитке, рассматриваются в качестве низшего уровня упаковки ДНК эукариот в хроматине. Нуклеосомы, входящие в состав фибрилл, расположены более или менее равномерно вдоль молекулы ДНК на расстоянии 10–20 нм друг от друга. В состав нуклеосом входят четыре пары молекул гистонов: H2a, H2b, H3 и H4, а также одна молекула гистона H1. Данные по структуре нуклеосом в основном получены с использованием трех методов: рентгеноструктурного анализа низкого и высокого разрешения кристаллов нуклеосом, межмолекулярных сшивок белок–ДНК и расщепления ДНК в составе нуклеосом с помощью нуклеаз или радикалов гидроксила. На основании таких данных А. Клугом была построена модель нуклеосомы, в соответствии с которой ДНК (146 п.о.) в B-форме (правозакрученная спираль с шагом 10 п.о.) намотана на гистоновый октамер, в центральной части которого расположены гистоны Н3 и Н4, а на периферии – Н2а и Н2b.
аметр такого нуклеосомного диска составляет 11 нм, а его толщина – 5,5 нм. Структура, состоящая из гистонового октамера и намотанной на него ДНК, получила название нуклеосомной кóровой частицы. Кóровые частицы отделены друг от друга сегментами линкерной ДНК. Общая длина участка ДНК, включенного в нуклеосому животных, составляет 200 (15) п.о.
Полипептидные цепи гистонов содержат структурные домены нескольких типов. Центральный глобулярный домен и гибкие выступающие N- и С-концевые участки, обогащенные основными аминокислотами, получили название плеч (arm). С-концевые домены полипептидных цепей, участвующие в гистон–гистоновых взаимодействиях внутри кóровой частицы, находятся преимущественно в виде -спирали с протяженным центральным спиральным участком, вдоль которого с двух сторон уложено по одной более короткой спирали. Все известные места обратимых посттрансляционных модификаций гистонов, происходящих на протяжении клеточного цикла или во время дифференцировки клеток, локализованы в гибких основных доменах их полипептидных цепей (табл. I.2). При этом N-концевые плечи гистонов H3 и H4 являются самыми консервативными участками молекул, а гистоны в целом – одними из наиболее эволюционно консервативных белков. С помощью генетических исследований дрожжей S. cerevisiae было установлено, что небольшие делеции и точковые мутации в N-концевых частях генов гистонов сопровождаются глубокими и разнообразными изменениями фенотипа дрожжевых клеток. Это указывает на чрезвычайную важность целостности молекул гистонов в обеспечении правильного функционирования эукариотических генов.
В растворе гистоны Н3 и Н4 могут существовать в виде стабильных тетрамеров (Н3)2(Н4)2, а гистоны Н2А и Н2В – в виде стабильных димеров. Постепенное повышение ионной силы в растворах, содержащих нативный хроматин, приводит к освобождению сначала димеров Н2А/Н2В, а затем тетрамеров Н3/Н4.
Дальнейшее уточнение тонкой структуры нуклеосом в кристаллах было проведено недавно в работе К. Люгера с соавт. (1997 г.) с помощью рентгеноструктурного анализа высокого разрешения. Было установлено, что выпуклая поверхность каждого гистонового гетеродимера в составе октамера огибается сегментами ДНК длиной 27–28 п.о., расположенными по отношению друг к другу под углом 140о, которые разделены линкерными участками длиной в 4 п.о.
В соответствии с современными данными пространственная структура ДНК в составе кóровых частиц несколько отличается от B-формы: двойная спираль ДНК перекручена на 0,25–0,35 п.о./виток двойной спирали, что приводит к образованию шага спирали, равному 10,2 п.о./виток (у В-формы в растворе – 10,5 п.о./виток).
абильность комплекса гистонов в составе кóровой частицы определяется взаимодействием их глобулярных частей, поэтому удаление гибких плеч в условиях мягкого протеолиза не сопровождается разрушением комплекса. N-концевые плечи гистонов, по-видимому, обеспечивают их взаимодействие со специфическими участками ДНК. Так, N-концевые домены гистона Н3 контактируют с участками ДНК на входе в кóровую частицу и выходе из нее, тогда как соответствующий домен гистона Н4 связывается с внутренней частью ДНК нуклеосомы.
Упомянутые выше исследования структуры нуклеосом высокого разрешения показывают, что центральная часть сегмента ДНК длиной в 121 п.о. в составе нуклеосомы образует дополнительные контакты с гистоном H3. При этом N-концевые части полипептидных цепей гистонов H3 и H2B проходят через каналы, образуемые малыми бороздками соседних супервитков ДНК нуклеосомы, а N-концевая часть гистона H2A контактирует с малой бороздкой внешней части супервитка ДНК. В совокупности данные высокого разрешения показывают, что ДНК в составе коровых частиц нуклеосом огибает гистоновые октамеры неравномерно. Кривизна нарушается в местах взаимодействия ДНК с поверхностью гистонов, и такие изломы наиболее заметны на расстоянии 10–15 и 40 п.о. от центра супервитка ДНК.
Таблица I.2
Источник: studfile.net
Определение понятия
Хроматином называется содержащий наследственную информацию материал клеточного ядра, представляющий собой сложный функциональный комплекс ДНК со структурными белками и другими элементами, обеспечивающими упаковку, хранение и реализацию кариотического генома. В упрощенной трактовке это вещество, из которого состоят хромосомы. Термин происходит от греческого «хрома» – цвет, краска.
Понятие было введено Флемингом еще в 1880 году, но до сих пор идут споры о том, что такое хроматин, с точки зрения биохимического состава. Неопределенность касается небольшой части компонентов, не участвующих в структурировании генетических молекул (некоторые ферменты и рибонуклеиновые кислоты).
На электронной фотографии интерфазного ядра хроматин визуализируется как многочисленные участки темной материи, которые могут быть мелкими и разрозненными или объединяться в крупные плотные скопления.
Конденсация хроматина во время клеточного деления приводит к образованию хромосом, которые видны даже в обычном световом микроскопе.
Структурные и функциональные компоненты хроматина
С целью определить, что такое хроматин на биохимическом уровне, ученые экстрагировали это вещество из клеток, переводили в раствор и в таком виде изучали компонентный состав и структуру. При этом использовались как химические, так и физические методы, включая технологии электронной микроскопии. Выяснилось, что химический состав хроматина на 40% представлен длинными молекулами ДНК и почти на 60% – различными белками. Последние подразделяются на две группы: гистоны и негистоновые.
Гистонами называют большое семейство основных ядерных белков, которые прочно связываются с ДНК, формируя структурный скелет хроматина. Их количество примерно равно процентному содержанию генетических молекул.
Остальная часть (до 20%) протеиновой фракции приходится на ДНК-связывающие и пространственно-модифицирующие белки, а также ферменты, принимающие участие в процессах считывания и копирования генетической информации.
Помимо основных элементов, в составе хроматина в небольшом количестве обнаруживаются рибонуклеиновые кислоты (РНК), гликопротеиды, углеводы и липиды, однако вопрос об их ассоциации с ДНК-упаковочным комплексом до сих пор открыт.
Гистоны и нуклеосомы
Молекулярная масса гистонов варьирует в пределах от 11 до 21 кДа. Большое количество остатков основных аминокислот лизина и аргинина придают этим белкам положительный заряд, способствуя формированию ионных связей с противоположно заряженными фосфатными группами двойной спирали ДНК.
Выделяют 5 разновидностей гистонов: H2A, H2B, H3, H4 и H1. Первые четыре типа участвуют в формировании основной структурной единицы хроматина – нуклеосомы, которая состоит из кора (белковой сердцевины) и обмотанной вокруг него ДНК.
Нуклеосомный кор представлен октамерным комплексом из восьми молекул гистонов, в который входят тетрамер H3-H4 и димер Н2A-H2B. Участок ДНК протяженностью около 146 нуклеотидных пар накручивается на поверхность белковой частицы, образуя 1,75 витка, и переходит в линкерную последовательность (примерно 60 н. п.), соединяющую нуклеосомы друг с другом. Молекула H1 связывается с линкерной ДНК, защищая ее от действия нуклеаз.
Гистоны могут подвергаться различным модификациям, таким как ацетилирование, метилирование, фосфорилирование, ADP-рибозилирование и взаимодействие с убивиктиновым белком. Эти процессы влияют на пространственную конфигурацию и плотность упаковки ДНК.
Негистоновые белки
Существует несколько сотен разновидностей негистоновых белков с различными свойствами и функциями. Их молекулярная масса варьирует от 5 до 200 кДа. Особую группу составляют сайт-специфические белки, каждый из которых комплементарен определенному участку ДНК. В эту группу входят 2 семейства:
- «цинковые пальцы» – узнают фрагменты длиной в 5 нуклеотидных пар;
- гомодимеры – характеризуются структурой «спираль-поворот-спираль» во фрагменте, связанном с ДНК.
Лучше всего изучены так называемые белки высокой подвижности (HGM-белки), постоянно ассоциированые с хроматином. Такое наименование семейство получило из-за высокой скорости перемещения белковых молекул в электрофорезном геле. Эта группа занимает большую часть негистоновой фракции и включает в себя четыре основных типа HGM-белков: HGM-1, HGM-14, HGM-17 и HMO-2. Они выполняют структурную и регуляторную функции.
К негистоновым белкам относят также ферменты, обеспечивающие транскрипцию (процесс синтеза матричной РНК), репликацию (удвоение ДНК) и репарацию (устранение повреждений в генетической молекуле).
Уровни компактизации ДНК
Особенность структуры хроматина такая, что позволяет нитям ДНК с суммарной длиной более метра поместиться в ядро диаметром около 10 мкм. Такое возможно благодаря многоступенчатой системе упаковки генетических молекул. Общая схема компактизации включает пять уровней:
- нуклеосомная нить диаметром 10–11 нм;
- фибрилла 25–30 нм;
- петлевые домены (300 нм);
- волокно толщиной 700 нм;
- хромосомы (1200 нм).
Такая форма организации обеспечивает уменьшение длины исходной молекулы ДНК в 10 тысяч раз.
Нить диаметром 11 нм образована рядом нуклеосом, связанных линкерными участками ДНК. На электронной микрофотографии такая структура напоминает нанизанные на леску бусы. Нуклеосомная нить сворачивается в спираль по типу соленоида, образуя фибриллу толщиной 30 нм. В ее формировании участвует гистон H1.
Соленоидная фибрилла складывается в петли (иначе – домены), которые закрепляются на поддерживающем внутриядерном матриксе. Каждый домен содержит от 30 до 100 тысяч пар нуклеотидов. Такой уровень компактизации характерен для интерфазного хроматина.
Структура толщиной 700 нм образуется при спирализации доменной фибриллы и называется хроматидой. В свою очередь, две хроматиды формируют пятый уровень организации ДНК – хромосому диаметром 1400 нм, которая становится видна на стадии митоза или мейоза.
Таким образом, хроматин и хромосома – это формы упаковки генетического материала, зависящие от жизненного цикла клетки.
Хромосомы
Хромосома состоит из двух идентичных друг другу сестринских хроматид, каждая из которых образована одной суперспирализованной молекулой ДНК. Половинки соединяются особым фибриллярным тельцем, называемым центромерой. Одновременно эта структура является перетяжкой, разделяющей каждую хроматиду на плечи.
В отличие хроматина, представляющего собой структурный материал, хромосома – это дискретная функциональная единица, характеризующаяся не только структурой и составом, но и уникальным генетическим набором, а также определенной ролью в реализации механизмов наследственности и изменчивости на клеточном уровне.
Эухроматин и гетерохроматин
Хроматин в ядре существует в двух формах: менее спирализованной (эухроматин) и более компактной (гетерохроматин). Первая форма соответствует транскрипционно-активным участкам ДНК и поэтому структурирована не так плотно. Гетерохроматин подразделяется на факультативный (может переходить из активной формы в плотную неактивную в зависимости от стадии жизненного цикла клетки и необходимости реализовать те или иные гены) и конститутивный (постоянно уплотнен). Во время митотического или мейотического деления весь хроматин неактивен.
Конститутивный гетерохроматин обнаружен возле центромер и в концевых участках хромосомы. Результаты электронной микроскопии показывают, что такой хроматин сохраняет высокую степень конденсации не только на стадии деления клетки, но и во время интерфазы.
Биологическая роль хроматина
Основная функция хроматина заключается в плотной упаковке большого количества генетического материала. Однако просто уместить ДНК в ядре для жизнедеятельности клетки недостаточно. Необходимо, чтобы эти молекулы должным образом «работали», то есть, могли передавать заключенную в них информацию по системе ДНК-РНК-белок. Кроме этого, клетке нужно распределять генетический материал во время деления.
Устройство хроматина полностью отвечает этим задачам. Белковая часть содержит все необходимые ферменты, а особенности структуры позволяют им взаимодействовать с определенными участками ДНК. Поэтому, второй важной функцией хроматина является обеспечение всех процессов, связанных с реализацией ядерного генома.
Источник: www.syl.ru
У всех ли есть ядро?
Все живые существа, от самых мелких до гигантских, снабжены генетической информацией в виде дезоксирибонуклеиновой кислоты. Существуют две принципиально отличные формы представления ее в клетках:
- Прокариотические организмы (доядерные) имеют не разделенные на компартменты клетки. Хранилище их единственной кольцевой ДНК, не соединенной с белками, это просто участок цитоплазмы, называемый нуклеоидом. Репликация нуклеиновой кислоты и синтез белков проходят у прокариот в едином пространстве клеток. Мы не увидим их невооруженным глазом, ведь представителями этой группы организмов являются микроскопические, размером до 3 мкм, бактерии.
- Эукариотические организмы характеризуются более сложным устройством клетки, где наследственная информация защищена двойной мембраной ядра. Линейные молекулы ДНК совместно с белками-гистонами образуют хроматин, активно продуцирующий РНК при помощи полиферментных комплексов. Синтез белка происходит в цитоплазме на рибосомах.
Оформленное ядро в клетках эукариот можно увидеть в период интерфазы. В кариоплазме содержится белковый остов (матрикс), ядрышки и нуклеопротеиновые комплексы, состоящие из участков гетерохроматина и эухроматина. Это состояние ядра сохраняется до момента начала деления клетки, когда оболочка и ядрышки исчезают, а хромосомы приобретают компактную палочковидную форму.
Главный в ядре
Основной компонент содержимого ядра, хроматин, является его смысловой частью. Его функции включают хранение, реализацию и передачу генетической информации о клетке или организме. Непосредственно реплицируемая часть хроматина — это эухроматин, несущий данные о структуре белков и различных типов РНК.
Остальные части ядра выполняют вспомогательные функции, обеспечивают надлежащие условия для реализации генетической информации:
- ядрышки — уплотненные участки ядерного содержимого, определяющие места синтеза рибонуклеиновых кислот для рибосом;
- белковый матрикс упорядочивает расположение хромосом и всего содержимого ядра, поддерживает его форму;
- полужидкая внутренняя среда ядра, кариоплазма, обеспечивает транспорт молекул и протекание различных биохимических процессов;
- двухслойная оболочка ядра, кариолемма, защищает генетический материал, обеспечивает избирательное двустороннее проведение молекул и молекулярных комплексов за счет сложно устроенных ядерных пор.
Что значит хроматин
Свое название хроматин получил в 1880 году благодаря опытам Флемминга по наблюдению клеток. Дело в том, что во время фиксации и окрашивания некоторые части клетки особенно хорошо проявляются («хроматин» значит «окрашиваемый»). Позже выяснилось, что этот компонент представлен ДНК с белками, которая, благодаря кислотным свойствам, активно воспринимает щелочные красители.
В центральной части клетки на фото видны окрашенные хромосомы, образующие метафазную пластинку.
Формы существования ДНК
В клетках эукариотических организмов нуклеопротеиновые комплексы хроматина могут находиться в двух состояниях.
- В процессе деления клетки ДНК достигает максимальной скрученности и представлена митотическими хромосомами. Каждая нить образует отдельную хромосому.
- В период интерфазы, когда ДНК клеток наиболее деконденсирована, хроматин равномерно заполняет пространство ядра или образует видимые в световой микроскоп сгустки. Такие хромоцентры чаще выявляются близ ядерной мембраны.
Данные состояния альтернативны друг другу, в интерфазе не сохраняется полностью компактизированных хромосом.
Эухроматин и гетерохроматин
Интерфазный хроматин представляет собой хромосомы, утратившие компактную форму. Их петли разрыхляются, заполняя объем ядра. Существует прямая зависимость между степенью деконденсации и функциональной активностью хроматина.
Его участки, полностью «распутанные», называются диффузным или активным хроматином. Он практически не виден в световой микроскоп после окрашивания. Это объясняется тем, что толщина спирали ДНК составляет всего 2 нм. Другое его название — эухроматин.
Это состояние обеспечивает доступ ферментных комплексов к смысловым фрагментам ДНК, их свободное присоединение и функционирование. С диффузных участков считывается РНК-полимеразами структура информационных РНК (транскрибирование) либо происходит копирование самой ДНК (репликация). Чем выше синтетическая активность клетки в данный момент, тем больше доля эухроматина в ядре.
Диффузные участки хроматина чередуются с компактными, скрученными в разной степени зонами гетерохроматина. Из-за большей плотности, окрашенный гетерохроматин хорошо заметен в интерфазных ядрах.
На рисунке изображен хроматин различной степени компактизации:
- 1 — двухцепочечная молекула ДНК;
- 2 — гистоновые белки;
- 3 — ДНК, обернутая вокруг гистонного комплекса на 1,67 оборота, формирует нуклеосому;
- 4 — соленоид;
- 5 — интерфазная хромосома.
Тонкости определения
Эухроматин в конкретный момент времени может быть не задействован в синтетических процессах. В таком случае он временно находится в более компактном состоянии и может быть принят за гетерохроматин.
Настоящий гетерохроматин, его еще называют конститутивным, не несет смысловой нагрузки и деконденсируется только в процессе репликации. ДНК в этих местах содержит короткие повторяющиеся последовательности, не кодирующие аминокислот. В митотических хромосомах они оказываются в области первичной перетяжки и теломерных окончаний. Также они разделяют участки танскрибируемой ДНК, образуя вставочные (интеркалярные) фрагменты.
Как «работает» эухроматин
В состав эухроматина входят гены, определяющие в конечном счете структуру белков (структурные гены). Дешифровка нуклеотидной последовательности в белок происходит при помощи посредника, способного, в отличие от хромосом, покинуть ядро — информационной РНК.
В процессе транскрипции РНК синтезируется на матрице ДНК из свободных адениловых, уридиловых, цитидиловых и гуаниловых нуклеотидов. Транскрипцию осуществляет ферментный комплекс РНК-полимераза.
Некоторые гены определяют последовательность других видов РНК (транспортной и рибосомной), необходимых для завершения в цитоплазме процессов синтеза белка из аминокислот.
Гетерохроматин отдельной хромосомы часто бывает собран в хорошо заметный хромоценр. Вокруг него располагаются петли деспирализованного эухроматина. Благодаря такой конфигурации ДНК ядра, к смысловым частям легко подходят ферментные комплексы и свободные нуклеотиды, необходимые для реализации функций эухроматина.
Источник: FB.ru