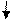
Схема строения плазматической мембраны
СТРОЕНИЕ И ФУНКЦИИ БИОЛОГИЧЕСКИХ МЕМБРАН. ЛИПИДЫ БИОМЕМБРАН. МЕХАНИЗМЫ ТРАНСПОРТА ВЕЩЕСТВ ЧЕРЕЗ БИОМЕМБРАНЫ
1. Многообразие мембранных структур и выполняемых ими функций.
2. Строение липидов, входящих в состав клеточных мембран: формулы фосфолипидов, гликолипидов, холестерола. Амфипатические свойства липидов мембран.
3. Белки мембран (интегральные, периферические): особенности структуры, свойства, функции. Взаимодействия белков и липидов в биологических мембранах.
4. Асимметрия мембран (примеры). Способность белков и липидов мембран к латеральной диффузии. Ограниченная возможность поперечной диффузии в мембранах.
5. Транспорт веществ через мембраны: простая и облегчённая диффузия, активный транспорт, экзо- и эндоцитоз, их особенности.
6. Na+, K+-АТФ-аза, механизм действия, роль в поддержании трансмембранного потенциала и возбудимости мембраны.
Ссылки по теме:
Березов Т.Т., Коровкин Б.Ф. Биологическая химия. Глава 9. Биомембраны и биоэнергетика. Основные принципы организации биомембран.
| Раздел 9.1 |
Классификация липидов и их роль в организме. |
|
|
|
|
9.1.1. Из курса биоорганической химии известно, что липиды – это большая группа соединений, которые существенно различаются по своему химическому строению и биологической роли. Общими признаками липидов являются:
Основные классы липидов представлены на рисунке 9.1.
Рисунок 9.1. Основные классы липидов. 9.1.2. Липиды выполняют в организме ряд функций. Главными из них являются следующие:
|
|
| Раздел 9.2 |
Классификация и функции биологических мембран. |
|
|
|
|
9.2.1. Биологические мембраны – сложные надмолекулярные структуры, окружающие все живые клетки и образующие в них замкнутые, специализированные компартменты – органеллы. Основные разновидности биологических мембран представлены на рисунке 9.2. Мембрану, ограничивающую цитоплазму клетки снаружи, называют цитоплазматической или плазматической мембраной (1). Название внутриклеточных мембран обычно происходит от названия ограничиваемых или образуемых ими субклеточных структур. Различают ядерную (2), митохондриальную (3), лизосомальную (4) мембраны, мембраны комплекса Гольджи (5), эндоплазматический ретикулум (6) и другие.
Рисунок 9.2. Схематическое изображение органелл животной клетки (объяснения в тексте). 9.2.2. Некоторые примеры функций биологических мембран. Плазматическая мембрана – ограничивает содержимое клетки от внешней среды; осуществляет контакт с другими клетками, получение, обработку и передачу информации внутрь клетки, поддержание постоянства внутренней среды. Ядерные мембраны (внешняя и внутренняя) – образуют ядерную оболочку, которая отделяет хромосомный материал от цитоплазматических органелл; через поры ядерной оболочки происходит транспорт белков и нуклеиновых кислот в ядро и из ядра. Митохондриальные мембраны – осуществляют преобразование энергии в ходе окислительного фосфорилирования, синтез АТФ. Лизосомальные мембраны – ограничивают гидролитические ферменты от цитоплазмы клетки, препятствуют самоперевариванию (аутолизу) клеток, способствуют поддержанию постоянства рН среды в лизосомах. Мембраны эндоплазматического ретикулума – принимают участие в образовании новых мембран, осуществляют синтез белков, липидов, полисахаридов, окисление гидрофобных метаболитов и ксенобиотиков. |
|
| Раздел 9.3 |
Состав и строение мембран. |
|
|
|
|
9.3.1. Все мембраны по своей организации и составу обнаруживают ряд общих свойств. Они:
9.3.2. Схема строения биологической мембраны, представлена на рисунке 9.3. Основу мембраны составляет липидный бислой – двойной слой молекул липидов, которые обладают свойствомамфифильности (содержат как гидрофильные, так и гидрофобные функциональные группы). В липидном бислое гидрофобные участки молекул взаимодействуют между собой, а гидрофильные участки обращены в окружающую водную среду. Рисунок 9.3. Схема строения биологической мембраны (по Сингеру и Николсону). Мембранные липиды выполняют роль растворителя мембранных белков, создавая жидкую среду, в которой они могут функционировать. По степени влияния на структуру бислоя и по силе взаимодействия с ним мембранные белки делят на интегральные и периферические. Важнейшие особенности интегральных и периферических белков представлены в таблице 9.1. Таблица 9.1 Характеристика мембранных белков По выполняемым функциям белки в составе мембран делятся на
Количество белков в мембранах могут существенно отличаться. Например, в миелиновой мембране, предназначенной для изоляции нервных волокон, белки составляют только 25% массы мембраны, а в мембранах митохондрий, связанных с процессами окислительного фосфорилирования, на долю белков приходится около 75% массы. В плазматической мембране доля белков и липидов примерно одинаковы. Углеводы в составе мембран не представлены самостоятельными соединениями, а обнаруживаются только в соединении с белками (гликопротеины) или липидами (гликолипиды). Длина углеводных цепей колеблется от двух до восемнадцати остатков моносахаридов. Большая часть углеводов расположена на наружной поверхности плазматической мембраны. Функции углеводов в биомембранах – контроль за межклеточными взаимодействиями, поддержание иммунного статуса, рецепция, обеспечение стабильности белковых молекул в мембране. |
|
| Раздел 9.4 |
Липидный состав мембран |
|
|
|
|
9.4.1. Как уже упоминалось (9.1), компонентами липидов являются остатки жирных кислот и одно- или многоатомных спиртов. Примеры жирных кислот, встречающихся в составе липидов мембран, представлены на рисунке 9.4. Выучите эти формулы.
Рисунок 9.4. Наиболее часто встречающиеся природные жирные кислоты. Основные особенности строения жирных кислот, входящих в состав природных жиров:
9.4.2. Большинство липидов в мембранах млекопитающих представлены фосфолипидами, гликосфинголипидами и холестеролом. Фосфолипиды в составе мембран подразделяются на две группы: глицерофосфолипиды и сфингомиелины. Глицерофосфолипиды – представляют собой сложные эфиры трёхатомного спирта глицерола, двух остатков жирных кислот и фосфорилированного аминоспирта. Общая формула глицерофосфолипида представлена на рисунке 9.5. Наиболее распространённым глицерофосфолипидом мембран является фосфатидилхолин:
В глицерофосфолипидах у второго углеродного атома глицерола обязательно находится остаток ненасыщенной жирной кислоты (в данном случае линолевой).
Рисунок 9.5. Общая формула глицерофосфолипидов. Сфингофосфолипиды (сфингомиелины) являются производными аминоспирта сфингозина (рисунок 9.6). Соединение сфингозина и жирной кислоты получило название церамид. Рисунок 9.6. Структурные формулы сфингозина и его производных. В сфингомиелинах водород гидроксильной группы у первого углеродного атома в церамиде замещён на фосфохолин. Пример сфингомиелина, содержащего остаток олеиновой кислоты:
Гликолипиды также являются производными церамида, содержащими один или несколько остатков моносахаридов. Например, цереброзиды содержат в первом положении остаток глюкозы или галактозы:
а ганглиозиды содержат цепочку из нескольких остатков сахаров, одним из которых обязательно является сиаловая кислота. Холестерол (рисунок 9.7) – одноатомный циклический спирт. Это один из главных компонентов плазматической мембраны клеток млекопитающих, в меньшем количестве может присутствовать также в митохондриях, мембранах комплекса Гольджи, ядерных мембранах. Особенно много его в нервной ткани. Рисунок 9.7. Структурные формулы холестерола и его эфира. 9.4.3. Как уже было сказано, характерной особенностью мембранных липидов является их амфифильность – наличие в молекуле одновременно гидрофобных и гидрофильных участков. Гидрофобная часть молекулы представлена остатками жирных кислот и боковой цепью сфингозина. Гидрофильные участки представлены в фосфолипидах фосфорилированным спиртом, а в гликолипидах – остатками сахаров. Амфифильность холестерола выражена слабо – циклическая структура и боковой радикал гидрофобны, и только гидроксильная группа гидрофильна. Амфифильность мембранных липидов определяет характер их поведения в водной среде. Слипание гидрофобных участков молекул приводит к образованию упорядоченных замкнутых структур – мицелл, в которых гидрофобные области защищены от воды, а гидрофильные обращены в водную среду. Молекулы холестерола встраиваются между радикалами жирных кислот гидрофобной части бислоя, а его гидроксильная группа примыкает к гидрофильным головкам фосфолипидов. Такая структура, стабилизированная нековалентными гидрофобными взаимодействиями, термодинамически очень устойчива и лежит в основе формирования биологических мембран. 9.4.4. Замкнутый липидный бислой определяет основные свойства мембран: 1) текучесть – зависит от соотношения насыщенных и ненасыщенных жирных кислот в составе мембранных липидов. Гидрофобные цепочки насыщенных жирных кислот ориентированы параллельно друг другу и образуют жёсткую кристаллическую структуру (рисунок 9.8, а). Ненасыщенные жирные кислоты, имеющие изогнутую углеводородную цепь, нарушают компактность упаковки и придают мембране бóльшую жидкостность (рисунок 9.8, б). Холестерол, встраиваясь между жирными кислотами, уплотняет их и повышает жёсткость мембран.
Рисунок 9.8. Влияние жирнокислотного состава фосфолипидов на текучесть липидного бислоя. 2) латеральная диффузия – свободное перемещение молекул относительно друг друга в плоскости мембран (рисунок 9.9,а).
Рисунок 9.9. Виды перемещений фосфолипидных молекул в липидном бислое. 3) ограниченная способность к поперечной диффузии (переходу молекул из наружного слоя во внутренний и наоборот, см. рисунок 9.9, б), что способствует сохранению асимметрии – структурно-функциональных различий наружного и внутреннего слоёв мембраны. 4) непроницаемость замкнутого бислоя для большинства водорастворимых молекул. |
|
| Раздел 9.5 |
Трансмембранный перенос веществ. |
|
|
|
|
9.5.1. Одна из главных функций мембран – участие в переносе веществ. Этот процесс обеспечивается при помощи трёх основных механизмов: простой диффузией, облегчённой диффузией и активным транспортом (рисунок 9.10). Запомните важнейшие особенности этих механизмов и примеры транспортируемых веществ в каждом случае.
Рисунок 9.10. Механизмы транспорта молекул через мембрану Простая диффузия — перенос веществ через мембрану без участия специальных механизмов. Транспорт происходит по градиенту концентрации без затраты энергии. Путём простой диффузии транспортируются малые биомолекулы – Н2О, СО2, О2, мочевина, гидрофобные низкомолекулярные вещества. Скорость простой диффузии пропорциональна градиенту концентрации. Облегчённая диффузия — перенос веществ через мембрану при помощи белковых каналов или специальных белков-переносчиков. Осуществляется по градиенту концентрации без затраты энергии. Транспортируются моносахариды, аминокислоты, нуклеотиды, глицерол, некоторые ионы. Характерна кинетика насыщения – при определённой (насыщающей) концентрации переносимого вещества в переносе принимают участие все молекулы переносчика и скорость транспорта достигает предельной величины. Активный транспорт – также требует участия специальных белков-переносчиков, но перенос происходит против градиента концентрации и поэтому требует затраты энергии. При помощи этого механизма через клеточную мембрану транспортируются ионы Na+, K+, Ca2+, Mg2+, через митохондриальную – протоны. Для активного транспорта веществ характерна кинетика насыщения. 9.5.2. Примером транспортной системы, осуществляющей активный транспорт ионов, является Na+,K+-аденозинтрифосфатаза (Na+,K+-АТФаза или Na+,K+-насос). Этот белок находится в толще плазматической мембраны и способен катализировать реакцию гидролиза АТФ. Энергия, выделяемая при гидролизе 1 молекулы АТФ, используется для переноса 3 ионов Na+ из клетки во внеклеточное пространство и 2 ионов К+ в обратном направлении (рисунок 9.11). В результате действия Na+,K+-АТФазы создаётся разность концентраций между цитозолем клетки и внеклеточной жидкостью. Поскольку перенос ионов неэквивалентен, то возникает разность электрических потенциалов. Таким образом, возникает электрохимический потенциал, который складывается из энергии разности электрических потенциалов Δφ и энергии разности концентраций веществ ΔС по обе стороны мембраны.
Рисунок 9.11. Схема Na+, K+-насоса. 9.5.3. Перенос через мембраны частиц и высокомолекулярных соединений Наряду с транспортом органических веществ и ионов, осуществляемым переносчиками, в клетке существует совершенно особый механизм, предназначенный для поглощения клеткой и выведения из неё высокомолекулярных соединений при помощи изменения формы биомембраны. Такой механизм называют везикулярным транспортом.
Рисунок 9.12. Типы везикулярного транспорта: 1 — эндоцитоз; 2 — экзоцитоз. При переносе макромолекул происходит последовательное образование и слияние окружённых мембраной пузырьков (везикул). По направлению транспорта и характеру переносимых веществ различают следующие типы везикулярного транспорта: Эндоцитоз (рисунок 9.12, 1) — перенос веществ в клетку. В зависимости от размера образующихся везикул различают: а) пиноцитоз — поглощение жидкости и растворённых макромолекул (белков, полисахаридов, нуклеиновых кислот) с помощью небольших пузырьков (150 нм в диаметре); б) фагоцитоз — поглощение крупных частиц, таких, как микроорганизмы или обломки клеток. В этом случае образуются крупные пузырьки, называемые фагосомами диаметром более 250 нм. Пиноцитоз характерен для большинства эукариотических клеток, в то время как крупные частицы поглощаются специализированными клетками — лейкоцитами и макрофагами. На первой стадии эндоцитоза вещества или частицы адсорбируются на поверхности мембраны, этот процесс происходит без затраты энергии. На следующей стадии мембрана с адсорбированным веществом углубляется в цитоплазму; образовавшиеся локальные впячивания плазматической мембраны отшнуровываются от поверхности клетки, образуя пузырьки, которые затем мигрируют внутрь клетки. Этот процесс связан системой микрофиламентов и является энергозависимым. Поступившие в клетку пузырьки и фагосомы могут сливаться с лизосомами. Содержащиеся в лизосомах ферменты расщепляют вещества, содержащиеся в пузырьках и фагосомах до низкомолекулярных продуктов (аминокислот, моносахаридов, нуклеотидов), которые транспортируются в цитозоль, где они могут быть использованы клеткой. Экзоцитоз (рисунок 9.12, 2) — перенос частиц и крупных соединений из клетки. Этот процесс, как и эндоцитоз, протекает с поглощением энергии. Основными разновидностями экзоцитоза являются: а) секреция — выведение из клетки водорастворимых соединений, которые используются или воздействуют на другие клетки организма. Может осуществляться как неспециализированными клетками, так и клетками эндокринных желёз, слизистой желудочно-кишечного тракта, приспособленными для секреции производимых ими веществ (гормонов, нейромедиаторов, проферментов) в зависимости от определённых потребностей организма. Секретируемые белки синтезируются на рибосомах, связанных с мембранами шероховатого эндоплазматического ретикулума. Затем эти белки транспортируются к аппарату Гольджи, где они модифицируются, концентрируются, сортируются, и затем упаковываются в пузырьки, которые отщепляются в цитозоль и в дальнейшем сливаются с плазматической мембраной, так что содержимое пузырьков оказывается вне клетки. В отличие от макромолекул, секретируемые частицы малых размеров, например, протоны, транспортируются из клетки при помощи механизмов облегчённой диффузии и активного транспорта. б) экскреция — удаление из клетки веществ, которые не могут быть использованы (например, удаление в ходе эритропоэза из ретикулоцитов сетчатой субстанции, представляющей собой агрегированные остатки органелл). Механизм экскреции, по-видимому, состоит в том, что вначале выделяемые частицы оказываются в цитоплазматическом пузырьке, который затем сливается с плазматической мембраной. |
|
Источник: dendrit.ru
Строение клеточной мембраны
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.

В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков. Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.
Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Большинство погруженных в клеточную мембрану белков выполняют ферментативную функцию (являются ферментами). Часто (особенно в мембранах органоидов клетки) ферменты располагаются в определенной последовательности так, что продукты реакции, катализируемые одним ферментом, переходят ко второму, затем третьему и т. д. Образуется конвейер, который стабилизируют поверхностные белки, т. к. не дают ферментам плавать вдоль липидного бислоя.
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции. Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).
При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO2, O2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану. Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример — натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами). Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз — это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа. При этом могут открываться каналы, и в клетку могут начать поступать определенные вещества или выводиться из нее.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) — синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
Источник: biology.su
Мембрана состоит из двух слоев липидных молекул (бислой), в который встроены белки. К некоторым липидным и белковым молекулам присоединены углеводы. Их немного. Толщина мембраны около 10 нм (0,00001 мм). Основную часть мембраны составляет непрерывный слой фосфолипидных молекул. В этот жидкийслой погружены молекулы различных по строению и функций белковых молекул. Белки не полностью покрывают липидный бислой, а располагаются в нём по отдельности или группами. В целом это напоминает мозаику (рис. 2. Б. В). В связи с этим принятая в настоящее время модель мембраны носит название жидкостно-мозаичная. Белки способны перемещаться по липидному слою. Перемещаются и молекулы липидного слоя. Понятно, что движение молекул мембраны меняет физико-химические характеристики последней, а это, в свою очередь, меняет функциональные характеристики мембраны. Необходимо отметить, что плазматическая мембрана большинства клеток не имеет форму идеального шара. Наоборот, она имеет множество выступов, углублений, которые постоянно меняют свою форму и величину. Полученные в последнее время результаты внесли некоторые коррективы в теорию строение мембран. Было показано, что не все мембранные белки способны к движению, а некоторые участки мембран отличаются по своей структуре от классического липидного бислоя.
Молекула фосфолипида имеет вид головки с двумя хвостиками (рис. 2 А). Головка (глицерин) растворима в воде, гидрофильна, хвостики (жирные кислоты) нерастворимы в воде, гидрофобны. Поэтому, находясь в воде, молекулы самопроизвольно занимают определённое положение по отношению к водной фазе. Отталкиваясь от молекул воды, хвостики располагаются в глубине липидного слоя, а водорастворимые головки обращены к внешней и внутренней водной среде (рис. 2. Б). Бислой липидов носит название матрикс. Особо следует отметить наличие в мембранах клеток липидов, хвостик которых содержит жирные кислоты, имеющие в своей структуре двойные связи, расположенные через CH2-группу (– СН = СН – СН = СН – СН –) . Такие жирные кислоты называют ненасыщенными. Эти кислоты наиболее всего подвержены воздействию активных форм кислорода (АФК), которые постоянно присутствуют в организме всех живых существ. Их количество особенно возрастает при различных заболеваниях. Это может привести к неблагоприятным последствиям, о чём мы скажем ниже.
Вкрапленные в матрикс белки (рис. 2 В) располагаются по-разному. Одни на внутренней и наружной поверхности липидного слоя – примембранные или поверхностные белки, другие полупогружены в мембрану – полуинтегральные белки, третьи пронизывают всю мембрану – интегральные белки. Обычно полуинтегральные и интегральные белки объединяют одним термином – внутренние белки, поскольку их трудно отличить друг от друга. Чаще всего в мембранах встречаются интегральные белки. Они могут быть представлены, как одной молекулой и выполнять какую-либо одну функцию, так и группой (ансамблем) белков. Каждый участник ансамбля выполняет строго определённую роль. Эти комплексы также выполняют одну или несколько конечных функций.
А Б В
| Фосфолипид 1-головка, 2- хвостик | Липидный бислой (матрикс) | Расположение молекул белка относительно липидного слоя: 1- примембранные белки, 2 – полупогружённый белок, 3 — интегральный белок |
Рис. 2. Схематическое строение плазматической мембраны.
Следует отметить, что интегральные и поверхностные белки-ферменты, функционирующие в мембране, достаточно часто меняют своё положение. В некоторых случаях сложно определить к какому типу (поверхностные или интегральные) относится тот или иной белок мембраны. Например, фермент фосфолипаза А, осаждаясь на мембрану, является поверхностным белком, но затем она активируется, становится интегральным белком и, взаимодействуя с липидами бислоя, образует из них арахидоновую кислоту (рис.3). Последняя покидает мембрану и превращается в другие активные соединения, которые участвуют в развитии различных патологических процессов.
Na+, K+— АТФ-аза Адренорецептор Аденилатциклаза
Ca2+-АТФ-аза
Фосфолипаза
|
|||||||||
|
|||||||||
|
|
||||||||
|
|||||||||
цАМФ
Na+, K+ Сa2+ Арахидоновая
кислота G-белок Гликоген
|
Глюкоза
Рис. 3. Гипотетическая схема локализации некоторых мембранных белков
Напротив, белки, участвующие в перемещении веществ через мембрану – например, белки, принимающие участие в облегчённой диффузии Na+, K+— АТФ-аза или Ca2+-АТФ-аза, как правило, не меняют своё положение, функционируя как интегральные белки (рис. 3). И, наконец, как мы уже говорили, в мембране могут находиться сложные комплексы из нескольких белков, связанных в единый ансамбль одной задачей. К таким комплексам относятся белки, принимающие участие в проведении информационного сигнала через мембрану (рис.3). К последним относится комплекс, содержащий три белка — адренорецептор, G-белок и аденилатциклазу. Все эти белки имеют существенное значение в нормальной жизнедеятельности клетки и при патологии. Об этом мы расскажем ниже.
Кроме липидов и белков в мембране имеются углеводы, но они располагаются не как самостоятельные компоненты, а как составные части липидов (гликолипиды) или белков (гликопротеинов). Углеводы располагаются на наружной поверхности плазмалеммы.
Источник: studopedia.su
Плазматическая мембрана, строение, функции.
Цитолемма – биологическая мембрана, окружающая клетку снаружи. Это самая толстая (10 нм) и сложно организованная мембрана клетки. В её основе лежит универсальная биологическая мембрана, покрытая снаружи гликокаликсом, а изнутри, со стороны цитоплазмы, подмембранным слоем (рис.2-1Б). Гликокаликс (3-4 нм толщины) представлен наружными, углеводными участками сложных белков – гликопротеинов и гликолипидов, входящих в состав мембраны. Эти углеводные цепочки играют роль рецепторов, обеспечивающих распознавание клеткой соседних клеток и межклеточого вещества и взаимодействие с ними. В этот слой также входят поверхностные и полуинтегральные белки, функциональные участки которых находятся в надмембранной зоне (например, иммуноглобулины). В гликокаликсе находятся рецепторы гистосовместимости, рецепторы многих гормонов и нейромедиаторов.
Подмембранный, кортикальный слой образован микротрубочками, микрофибриллами и сократимыми микрофиламентами, которые являются частью цитоскелета клетки. Подмембранный слой обеспечивает поддержание формы клетки, создание её упругости, обеспечивает изменения клеточной поверхности. За счёт этого клетка участвует в эндо- и экзоцитозе, секреции, движении.
Цитолемма выполняет множество функций:
1) разграничительная (цитолемма отделяет, отграничивает клетку от окружающей среды и обеспечивает её связь с внешней средой);
2) распознавание данной клеткой других клеток и прикрепление к ним;
3) распознавание клеткой межклеточного вещества и прикрепление к его элементам (волокнам, базальной мембране);
4) транспорт веществ и частиц в цитоплазму и из неё;
5) взаимодействие с сигнальными молекулами (гормонами, медиаторами, цитокинами) благодаря наличию на её поверхности специфических рецепторов к ним;
- обеспечивает движение клетки (образование псевдоподий) благодаря связи цитолеммы с сократимыми элементами цитоскелета.
В цитолемме расположены многочисленные рецепторы, через которые биологически активные вещества (лиганды, сигнальные молекулы, первые посредники: гормоны, медиаторы, факторы роста) действуют на клетку. Рецепторы представляют собой генетически детерминированные макромолекулярные сенсоры (белки, глико- и липопротеины) встроенные в цитолемму или расположенные внутри клетки и специализированные на восприятии специфических сигналов химической или физической природы. Биологически активные вещества при взаимодействии с рецептором вызывают каскад биохимических изменений в клетке, трансформируясь при этом в конкретный физиологический ответ (изменение функции клетки).
Все рецепторы имеют общий план строения и состоят из трёх частей: 1) надмебранной, осуществляющей взаимодействие с веществом (лигандом); 2) внутримембранной, осуществляющей перенос сигнала и 3) внутриклеточной, погружённой в цитоплазму.
Виды межклеточных контактов.
Цитолемма участвует также в образовании специальных структур – межклеточных соединений, контактов, которые обеспечивают тесное взаимодействие между рядом расположенными клетками. Различают простые и сложные межклеточные соединения. В простых межклеточных соединениях цитолеммы клеток сближаются на расстояние 15-20 нм и молекулы их гликокаликса взаимодействуют друг с другом (рис. 2-3). Иногда выпячивание цитолеммы одной клетки входит в углубление соседней клетки, образуя зубчатые и пальцевидные соединения (соединения «по типу замка»).
Сложные межклеточные соединения бывают нескольких видов: запирающие, сцепляющие и коммуникационные (рис. 2-3). К запирающим соединениям относят плотный контакт или запирающую зону. При этом интегральные белки гликокаликса соседних клеток образуют подобие ячеистой сети по периметру соседних эпителиальных клеток в их апикальных частях. Благодаря этому межклеточные щели запираются, отграничиваются от внешней среды (рис. 2-3).
Рис. 2-3. Различные типы межклеточных соединений.
- Простое соединение.
- Плотное соединение.
- Адгезивный поясок.
- Десмосома.
- Полудесмосома.
- Щелевое (коммуникационное) соединение.
- Микроворсинки.
(По Ю. И. Афанасьеву, Н. А. Юриной).
К сцепляющим, заякоревающим соединениям относят адгезивный поясок и десмосомы. Адгезивный поясок располагается вокруг апикальных частей клеток однослойного эпителия. В этой зоне интегральные гликопротеиды гликокаликса соседних клеток взаимодействуют между собой, а к ним со стороны цитоплазмы подходят подмембранные белки, включающие пучки актиновых микрофиламентов. Десмосомы (пятна сцепления) – парные структуры размером около 0,5 мкм. В них гликопротеиды цитолеммы соседних клеток тесно взаимодействуют, а со стороны клеток в этих участках в цитолемму вплетаются пучки промежуточных филаментов цитоскелета клеток (рис. 2-3).
К коммуникационным соединениям относят щелевидные соединения (нексусы) и синапсы. Нексусы имеют размер 0,5-3 мкм. В них цитолеммы соседних клеток сближаются до 2-3 нм и имеют многочисленные ионные каналы. Через них ионы могут переходить из одной клетки в другую, передавая возбуждение, например, между клетками миокарда. Синапсы характерны для нервной ткани и встречаются между нервными клетками, а также между нервными и эффекторными клетками (мышечными, железистыми). Они имеют синаптическую щель, куда при прохождении нервного импульса из пресинаптической части синапса выбрасывается нейромедиатор, передающий нервный импульс на другую клетку (подробнее см. в главе «Нервная ткань»).
Источник: alexmed.info