Цепи выедания пастбищные начинаются с
Такие трофические цепи, которые начинаются с зелёных растений, а далее идут к растительноядным животным, которые в свою очередь поедаются хищниками, называются цепями выедания (или пастбищные). Но если растения не съедены фитофагами или животное не съедено хищником, а отмирая (умирая) используется детритофагами, а те тоже могут служить пищей хищникам, то такие цепи называются детритными (или цепями разложения), в которых мёртвое вещество возвращается в кругооборот, минуя полное разложение (рис.4.6). Приведём примеры: листовой опад, мёртвые растения, трупы животных или их экскрименты могут быть потреблены детритофагами – дождевыми червями (листовой опад), жуками мертвоедами, жуками навозниками; а те не будут разрушены редуцентами грибами и бактериями, а будут съедены птицами или другими хищниками (барсук), и питательные элементы, которые содержатся в детрите будут повторно использованы, минуя стадию разложения.
Труп животного ® личинки мух-падалиц ® травяная лягушка ® уж
Такие сложные детритные цепи широко распространены в природе. Бактерии и грибы – редуценты и детритофаги – тоже могут в свою очередь служить пищей таким организмам, как простейшие одноклеточные микроорганизмы, клещи, насекомые и черви, живущие в почве и в воде. А когда грибы и другие редуценты отмирают, они сами превращаются в детрит и служат источником энергии и питательных веществ для других детритофагов и редуцентов.
Приведённые примеры упрощают действительную картину, так как одно и тоже растение может быть съедено тем или другим травоядным животным, а они, в свою очередь, стать жертвой разных хищников. Лист растения может съесть гусеница или слизень; гусеница может стать жертвой жука или насекомоядной птицы, которая может заодно и склевать и самого жука. Но жук может стать жертвой и паука. Поэтому в реальной природе пищевые цепи не изолированы друг от друга, они тесно переплетаются, образуя так называемые пищевые сети. Это связано с тем, что живые организмы редко специализируются на питании только одним каким-либо видом пищи (т.е. бывают стенофагами), некоторые животные даже относятся к категории всеядных, потребляя и растительную и животную пищу и детрит (крысы, барсук, медведь).

экологические ниши
С понятием жизненных форм организмов, которое мы с вами разбирали на второй лекции, тесно связано понятие об экологических нишах (ЭН). Под экологической нишей вида понимают его «положение» в общей системе биоценоза. Причём не территориальное его размещение, а функциональное проявление вида в сообществе – отношение к пище, к врагам, к внешним условиям. Итак, экологическую нишу можно определить как совокупность всех факторов (условий) и ресурсов среды, в пределах которой может существовать вид в природе.
Важно осознать, что ЭН не есть нечто такое, что можно увидеть. ЭН – это отвлечённое абстрактное понятие, сводящее к общему показателю всё, в чём нуждаются организмы того или иного вида – т.е. все необходимые условия и ресурсы в нужных количествах.
Экологи пользуются термином «местообитание» для обозначения места, где живёт организм, а термин ЭН они относят к роли, которую играют организмы данного вида в экосистеме. Можно сказать, что ниша – «профессия», «роль», «функция» вида; тогда как местообитание – его «адрес». Так кенгуру, бизон и корова – разные виды, но занимают сходную ЭН в экосистеме степей.
Ясно, что существование вида в сообществе определяется сочетанием и действием многих факторов, но в определении принадлежности вида к той или иной ЭН исходят, прежде всего, из характера питания этого вида, из способности добывать пищу.
Одно и то же местообитание содержит множество ниш. Чем больше разнообразие условий и ресурсов в нём, тем больше количество различных организмов может здесь жить, используя эти ресурсы. В связи с этим различают «насыщенные» и «ненасыщенные» биоценозы. В насыщенных биоценозах все ЭН заселены, жизненные ресурсы используются наиболее полно. Ненасыщенные биоценозы характеризуются наличием свободных ЭН. Один вид может быть безболезненно для всей экосистемы заменён другим видом со сходными экологическими потребностями, т.е. со сходной ЭН.
Ниши видов, проживающих в одном и том же местообитании, обычно различаются. Процесс разделения видами или популяциями видов пространства и ресурсов называется дифференциацией ЭН. Более наглядно этот процесс выражен у животных. У представителей разных видов рацион питания неодинаков: например, в озере одни виды живут и питаются в толще воды, другие ведут придонный образ жизни, третьи – на грани суши и воды. У разных видов и планктона и бентоса пища неодинакова. В лесу каждый ярус имеет свои условия. Растения, казалось бы, имеют одинаковый рацион питания: минеральные вещества, СО2, солнечная энергии и вода. Тем не менее, и у них ниши дифференцированы. Есть, как мы уже говорили, растения светолюбивые и теневыносливые; их корневые системы расположены на разной глубине, им требуются различные элементы минерального питания и в разных количествах, неодинаковое количество воды, в разное время они цветут и плодоносят, имеют собственных опылителей.
Справедливо также и то, что один и тот же вид может занимать различные ЭН в разных местообитаниях или географических районах. Хорошим примером является сам Человек. Где-то он плотояден (север), где-то растительнояден (юг), в большинстве же случаев человек является существом всеядным. Роль Человека и весь его образ жизни меняются в зависимости от того, на какой источник пищи он рассчитывает.
Характер занимаемой ЭН определяется как экологическими возможностями вида – фундаментальная ниша, так и тем, насколько эти возможности могут быть реализованы в конкретном биоценозе — реализованная ниша. Большинство видов не обитает в своей потенциальной фундаментальной нише, а вследствие взаимодействия с другими организмами занимает меньшую по размерам реализованную нишу. Вот, что пишет Дажо Р.: «К первой относится вся совокупность необходимых для существования и даже процветания вида условий среды при отсутствии какого бы то нибыло давления (конкуренции) со стороны других видов. Она соответствует максимальным экологическим возможностям вида. Вторая представляет собой часть потенциальной ниши, фактически занимаемой видом в биоценозе», которую он способен отстоять от конкурентов при их наличии. Если защитить растение от конкуренции, то оно освоит свою фундаментальную нишу. Вот почему в бот.садах растут растения из разных районов Земли и чувствуют себя неплохо, пока их охраняет человек. Практически в условиях фундаментальной ниши оказываются все культурные растения, которые человек защищает от сорняков.
Специализация вида по питанию, использованию пространства, времени активности и т.д. характеризуется как сужение его ЭН, обратный процесс – как её расширение.
Если ЭН, например, двух видов, представить в виде кружочков, то возможны 4 случая (рис.4.9.). Учёными было доказано, что два вида с совершенно одинаковыми потребностями не могут существовать вместе: один из них через какое-то время обязательно будет вытеснен. Т.о. вместе с концепцией ЭН развивалась концепция конкурентного исключения – закон «конкурентного вытеснения».
Рис.4.9. Возможные варианты «соотношений» экологических ниш (ЭН)
Важное следствие из этой концепции: если два вида со сходным образом жизни всё же существуют радом, то между ними должно быть какое-то экологическое различие – они либо используют разные ресурсы, либо активны в разное время, либо делят территорию – т.е каждый из них занимает всё же свою экологическую нишу (чему примеры мы уже приводили). Главный результат дифференциации (т.е. расхождения) ниш – снижение конкуренции. В данном случае речь должна идти о межвидовой конкуренции.
С другой стороны, ослабление межвидовой конкуренции приводит к расширению ЭН вида, напротив – усиление конкуренции сужает нишу, не давая проявиться всем его потенциям. В то же время внутривидовая конкуренция способствует расширению ЭН, т.к. при росте численности вида особи начинают использовать дополнительные корма, осваивать новые местообитания, вступают в новые биоценотические связи.
Механизмы выхода из конкурентных отношений и разграничения ЭН могут быть различны. В частности, размерная дифференциация – когда разные по размерам виды организмов используют сходную, но соразмерную себе пищу (крупные виды акул – крупных рыб-жертв, мелкие – мелких). Поведенческие различия проявляются в разной стратегии добывания пищи (пищуха обследует ствол дерева в поисках насекомых снизу вверх, а поползень – сверху вниз). Пространственная дифференциация позволяет разным видам в приделах, например, одного яруса сбора пищи использовать разные его участки (корни разных видов растений на разной глубине собирают влагу). Различия во времени активности относятся к использованию видами одного и того же ресурса в разное время суток (соколы и ястребы – дневные хищники, совы – ночные; растения цветут по очереди в течении лета). При наличии конкуренции с близкородственными или экологически сходными видами зона местообитания сокращается до оптимальных границ. Т.е. вид сохраняется в наиболее благоприятных для него зонах, где он обладает преимуществами по сравнению со своими конкурентами.
Принцип дифференциации ЭН используется в сельском хозяйстве: выращивание совместно нескольких сортов одного вида или даже нескольких видов растений (так называемые сортосмеси и поликультуры) позволяет за счёт снижения конкуренции между ними и более полного использования ресурсов почвы и света повышать урожай. При этом отсутствие свободных ниш снижает засорённость посевов и можно обходиться без гербицидов.
Дифференциация ниш у животных может сопровождаться сигналами, извещающими о том, что участок занят (мочевые метки зверей, пение птиц и т.п.). У растений сигнальных механизмов нет, хотя некоторые растения могут выделять в атмосферу или почву вещества, препятствующие росту соседей (аллелопатия, про которую мы уже говорили).
Источник: studopedia.ru
Пищевые цепи можно разделить на два основных типа: пастбищные цепи, которые начинаются с зеленого растения и идут дальше к пасущимся фитофагам, а затем к хищникам; детритные цепи, которые от мертвого органического вещества идут к детрито-фагам, а затем к хищникам. Пастбищные цепи, в свою очередь, объединяют пищевые цепи хищников и пищевые цепи паразитов.[ …]
Пищевая (трофическая) цепь — это перенос энергии от ее источника — продуцентов — через ряд организмов. Пищевые цепи можно разделить на два основных типа: пастбищная цепь, которая начинается с зеленого растения и идет далее к пасущимся растительноядным животным и к хищникам, и детритная цепь (от латинского истертый), которая начинается от продуктов распада мертвого органического вещества.
формировании этой цепи решающую роль играют различные микроорганизмы, которые питаются мертвым органическим веществом и минерализуют его, вновь превращая в простейшие неорганические соединения. Пищевые цепи не изолированы одна от другой, а тесно переплетаются друг с другом. Часто животное, потребляющее живое органическое вещество, поедает и микробов, потребляющих в пищу неживое органическое вещество. Таким образом, пути потребления пищи разветвляются, образуя так называемые пищевые сети.[ …]
Пастбищные пищевые цепи, или цепи эксплуататоров начинаются с продуцентов; для таких цепей при переходе с одного трофического уровня на другой характерно увеличение размеров особей при одновременном уменьшении плотности популяций, скорости размножения и продуктивности по биомассе (например, трава ■-» полевки —> лисица).[ …]
В случае пастбищных пищевых цепей леса, когда продуцентами служат деревья, а первичными консументами — насекомые, уровень первичных консументов численно богаче особями уровня продуцентов. Таким образом, пирамиды чисел могут быть обращенными. Для примера на рис. 9.7 приведены пирамиды чисел экосистем степи и леса умеренной зоны.[ …]
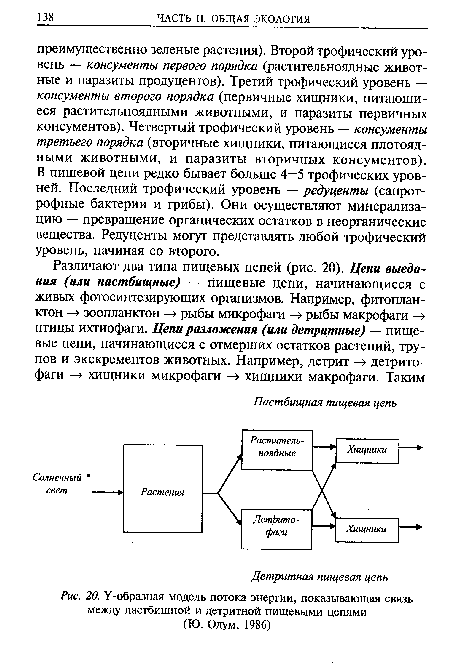
На рис. 3.10 пастбищные и детритные пищевые цепи показаны в виде Y-образной, или двухканальной, диаграммы потоков энергии.[ …]
На первом пищевом уровне растениями поглощается около 70 % падающего света, а превращается в энергию биомассы не более 1 % поглощенной энергии. Биомасса, синтезированная растениями, расходуется по трем направлениям: а) используется самими растениями на дыхание: б) потребляется животными, питающимися зелеными растениями, и начинает пастбищную пищевую цепь; в) оставшиеся растения отмирают, опадают на землю и начинают детритную пищевую цепь. Вторичная продуктивность пищевой цепи на уровне травоядных составляет около 10 % от поступающей первичной продуктивности.[ …]
Концепция пищевых цепей позволяет в дальнейшем проследить круговорот химических элементов в природе, хотя простые пищевые цепи, подобные изображенным ранее, где каждый организм представлен как питающийся организмами только какого-то одного типа, в природе встречаются редко. Реальные пищевые связи намного сложнее, ибо животное может питаться организмами разных типов, входящих в одну и ту же пищевую цепь или в различные цепи, что особенно характерно для хищников (консументов) высших трофических уровней. Связь между пастбищной и детритной пищевыми цепями иллюстрирует предложенная Ю. Одумом модель потока энергии (рис. 5.2).[ …]
| Схема пастбищной и детритной пищевых цепей (по 10. Одуму) | |
В первом типе пищевой сети поток энергии идет от растений к растительноядным животным, а далее к консументам более высокого порядка. Это сеть выедания, или пастбищная сеть. Вне зависимости от величины биоценоза и места обитания растительноядные животные (наземные, водные, почвенные) пасутся, выедают зеленые растения и передают энергию на следующие уровни (рис. 96).[ …]
Приведенные типы пищевых цепей начинаются с фотосинтезирующих организмов и носят название пастбищных (или цепи выедания, или цепи потребления).[ …]
Пути I и II соответствуют пастбищной и детритной пищевым цепям. По мнению некоторых специалистов, существует, кроме того, более прямой путь III. Путь автолиза, показанный на схеме, можно рассматривать как путь IV (Иоханнес, 1968).[ …]
Воду широко используют в пастбищных и ферменных биогеоценозах для поения животных, в антропогеоценозах — для удовлетворения разнообразных потребностей людей. Из водных экосистем в наземные мигрируют вещества по биогеохимической пищевой цепи вода -»ил -> растения (водные) -> животные (водные, земноводные, сухопутные). Масштабы миграции веществ по трофическому пути, по-видимому, невелики, но они играют существенную биогеоценотическую роль.[ …]
Существуют два главных типа пищевых цепей — пастбищные (или «выедания») и детритные (или «разложения»).[ …]
Ранние архантропы занимали нишу собирателей пастбищной пищевой цепи с относительно малой долей животной пищи. Они были вынуждены постоянно выполнять большую работу по добыче пищи и осваивать большую кормовую территорию (порядка п • 102 га/чел). Начало использования огня и увеличение добычи и потребления животной пищи расширило экологическое пространство человека до ниши первобытных охотников и рыболовов (с площадью п • 103 га/чел). К этому же периоду относится и применение огня для выжигания лесов, сначала для целей загонной охоты, а затем и для подсечноогневого земледелия. М. Ичас (1994) даже назвал это первой экологической революцией, но вряд ли саму по себе утрату локальных лесных массивов можно так квалифицировать. Гораздо существеннее изменение характера «землепользования» в эту эпоху, которую обычно называют неолитической (первой сельскохозяйственной) революцией.[ …]
В сельскохозяйственных экосистемах (агробиогеоценозах, пастбищных и ферменных БГЦ) пищевые цепи вовлечены в сферу деятельности человека. В них изменена экологическая пирамида. На вершине экологической пирамиды встал человек. Своеобразие экологической пирамиды, на вершине которой находится человек, — специфический признак любой сельскохозяйственной экосистемы.[ …]
| У-образная модель потока энергии, показывающая связь между пастбищной и детритной пищевыми цепями (Ю. Одум, 1986) | |
По цели производства различают товарные рыбоводные хозяйства, дающие пищевую рыбную продукцию, и нерестово-выростные хозяйства и рыборазводные заводы, производящие рыбопосадочный материал (молодь рыбы) для заселения водоемов, отведенных под пастбищную аквакультуру.[ …]
По мере роста молоди у части ее группировок начинает прояв-ляться иной пастбищный тип нагульного поведения, заключающийся в частой смене индивидуальных кормовых участков, резком увеличении двигательной активности, подъемах в толщу воды, чередованиях дрейфа в потоках с опусканиями на дно для питания. У рыб, перешедших на пастбищный образ жизни, значительно (примерно на порядок) увеличивается нагульный ареал, расширяется пищевой спектр и, видимо, ускоряется рост. Однако у популяций полной смены стойлового типа поведения на пастбищный не происходитоба типа сохраняются и у взрослых рыб на протяжении всей их жизни. Как показали биотелеметрические наблюдения, большая часть внутристадных нагульных группировок бентофагов, попав в зону аккумуляции биомассы, сохраняет стойловый тип поведения, обитает на освоенном нагульном участке весь вегетационный период и, зимуя вблизи от него вдоль склонов подводных возвышений дна, где лучше кислородный режим, весной компактными группами совершает преднерестовую миграцию в прибрежье.[ …]
По мере развития экосистемы следует ожидать тонких изменений в структуре пищевых цепей. Относительно простые и линейные связи между организмами, входящими в состав данной пищевой цепи, существуют только на очень ранних стадиях сукцессии. Более того, гетеротрофное использование чистой продукции происходит преимущественно в пастбищных пищевых цепях, т. е. в последовательности растение— травоядное — хищник. На зрелых стадиях, напротив, пищевые цепи превращаются в сложные сети, и при этом большая часть биологического потока энергии следует по детритному пути (табл. 37, п. 5), как это было подробно описано в гл. 3. При ненарушаемом течении сукцессии имеется достаточно времени для развития все более тесных связей и взаимных адаптаций между растениями и животными, и это ведет к образованию множества механизмов, уменьшающих выедание растений, например к образованию плохо перевариваемых опорных тканей, к развитию механизмов обратной связи между растениями и животными (Пиментел, 1961) и к увеличению давления хищников на растительноядных животных. Жесткий стресс или быстрые изменения, вызываемые внешними факторами, могут, конечно, разрушить эти защитные механизмы и вызвать чрезмерный (как бы «злокачественный») рост некоторых видов, что человек на свое горе слишком часто и обнаруживает.[ …]
Характерной особенностью аграрных ландшафтов как экосистем является то, что в них пищевые (трофические) цепи и биотический круговорот вовлечены в сферу человеческой деятельности. Человек оказывает влияние на условия питания растений и животных. Он улучшает условия минерального питания растений при помощи внесения в почвы удобрений, обогащения воздуха С02 (в теплицах и т. д.). Разработаны специальные технологии подкормки сельскохозяйственных культур, садово-ягодных и фруктовых растений, луговых трав. Предложены режимы кормления животных в условиях их пастбищного и стойлового содержания.[ …]
Ярусное строение наземных биоценозов тесно связано с их функциональной активностью. Так, пастбищные пищевые цепи преобладают в надземной части биоценозов, а детритные — в подземной. В водных экосистемах крупномасштабная вертикальная структура задается в первую очередь абиотическими условиями. Определяющими факторами являются градиенты освещенности, температуры, концентрации биогенов и т. п. На значительных глубинах усиливается влияние гидростатического давления. В донных биоценозах важны состав грунтов, гидродинамика придонных течений. Особенности вертикальной структуры выражаются в видовом составе, смене доминирующих видов, показателях биомассы и продуктивности. Фотосинтезирующие водоросли преобладают в верхних, хорошо освещенных горизонтах, что формирует вертикальные потоки вещества и энергии в направлении глубоководных биоценозов, жизнь которых основывается на привнесенной органике.[ …]
Ярусное строение наземных биоценозов тесно связано с их функциональной активностью. Так, пастбищные пищевые цепи преобладают в надземной части биоценозов, а детритные — в подземной. В водных экосистемах крупномасштабная вертикальная структура задается в первую очередь абиотическими условиями. Определяющими факторами являются градиенты освещенности, температуры, концентрации биогенов и т. п. На значительных глубинах усиливается влияние гидростатического давления. В донных биоценозах важны состав грунтов, гидродинамика придонных течений. Особенности вертикальной структуры выражаются в видовом составе, смене доминирующих видов, показателях биомассы и продуктивности. Фотосинтезирующие водоросли преобладают в верхних, хорошо освещенных горизонтах, что формирует вертикальные потоки вещества и энергии в направлении глубоководных биоценозов, жизнь которых основывается на привнесенной органике.[ …]
Существуют три основных пути возвращения питательных веществ в новые циклы поглощения. Первый соответствует пищевой цепи пастбищного типа, второй путь характерен для степей, лесов умеренной зоны и других сообществ, в которых основной поток энергии идет через детритную пищевую цепь. Третий путь — прямая передача питательных веществ от растения к растению т.н. симбиотическими организмами.[ …]
Перенос энергии пищи от ее источника — растений —через ряд организмов, происходящий путем поедания одних организмов другими, называется пищевой цепью. При каждом очередном переносе большая часть (80—90%) потенциальной энергии теряется, переходя в тепло. Это ограничивает возможное число этапов, или «звеньев» цепи, обычно до четырех-пяти. Чем короче пищевая цепь (или чем ближе организм к ее началу), тем больше количество доступной энергии. Пищевые цепи можно разделить на два основных типа: пастбищные цепи, которые начинаются с зеленого растения и идут далее к пасущимся, растительноядным животным (т. е. к организмам, поедающим зеленые растения) и к хищникам (организмам, поедающим животных), и детритные цепи, которые начинаются от мертвого органического вещества, идут к микроорганизмам, которые им питаются, а затем детритофагам и к их хищникам. Пищевые цепи не изолированы одна от другой, а тесно переплетены. Их сплетение часто называют пищевой сетью. В сложном природном сообществе организмы, получающие свою пищу от растений через одинаковое число этапов, считаются принадлежащими к одному трофическому уровню. Так, зеленые растения занимают первый трофический уровень (уровень продуцентов), травоядные — второй (уровень первичных консументов), хищники, поедающие травоядных, — третий (уровень вторичных консументов), а вторичные хищники — четвертый уровень (уровень третичных консументов). Необходимо подчеркнуть, что эта трофическая классификация делит на группы не сами виды, а их типы жизнедеятельности; популяция одного вида может занимать один или более трофических уровней, смотря по тому, какие источники энергии она использует. Поток энергии через трофический уровень равен общей ассимиляции (Л) на этом уровне, а общая ассимиляция в свою очередь равна продукции биомассы (Р) плюс дыхание (/?) .[ …]
В состав каждого биоценоза входят следующие функциональные компоненты: продуценты, консументы I—III порядков, а также редуценты, образующие пищевые цепи разных типов (пастбищные и детритные). Такая структура экосистемы обеспечивает перенос энергии от звена (трофического уровня) к звену. В реальных условиях цепи питания могут иметь разное число звеньев, кроме того, трофические цепи могут перекрещиваться, образуя сети питания. Почти все виды животных, за исключением очень специализированных в пищевом отношении, используют не один какой-нибудь источник пищи, а несколько. Если один член биоценоза выпадает из сообщества, вся система не нарушается, так как используются другие источники питания. Чем больше видовое разнообразие в биоценозе, тем он устойчивее. Например, в цепи питания растения-заяц—лиса всего три звена. Но лиса питается не только зайцами, но и грызунами и птицами. У зайца также есть альтернативные виды корма — зеленые части растений, сухие стебли (“сено”), веточки деревьев и кустарников и т.д.[ …]
Человек должен планировать свою хозяйственную деятельность с учетом цикличности природных процессов. Особенно тщательно ее следует учитывать в земледелии, пастбищном животноводстве, водоснабжении, навигации. Распашка, внесение минеральных удобрений, загрязнение нефтью и тяжелыми металлами весьма обедняют фауну почвы. При этом нарушаются и даже полностью выпадают звенья нормальных пищевых цепей и биогеохимических циклов. Реакция почвы на вмешательство человека необычайно велика.[ …]
Сельскохозяйственные угодья Земли включают огромное количество агробиогеоценозов. Агробиогеоценозы занимают около 1,2 млрд га, что составляет более 10 % всей поверхности суши. Пашня дает человечеству 88 % пищевой энергии. Кроме того, примерно 10 % этой энергии оно получает от пастбищных экосистем. Около 2 % пищевой энергии поставляет Мировой океан. Приведенные цифры убедительно показывают значение сельского хозяйства.[ …]
Как было показано выше, в зрелых экосистемах гетеротрофная утилизация первичной продукции в значительной мере связана с потреблением разлагаемого детрита. Нет причин,.по которым человек не мог бы увеличить использование детрита и получать таким образом пищевые и другие продукты от экосистем, относящихся скорее к про-тективным. Это вновь означало бы компромисс, так как «сиюминутный» урожай не был бы при этом таким высоким, как при непосредственной-эксплуатации пастбищной пищевой цепи. Сельское хозяйство, основанное на детрите, имеет, однако, ряд компенсирующих преимуществ. Современное сельское хозяйство основано на селекции растений на быстрый рост и пищевую ценность, что, конечно, делает их восприимчивыми к насекомым-вредителям и болезням. Следовательно, чем интенсивнее мы ведем отбор на такие признаки, как сочные листья и быстрый рост, тем больше усилий мы должны затрачивать на химические средства борьбы с болезнями, а это в свою очередь повышает вероятность отравления полезных животных, не говоря уже о самом человеке. Производство силоса из малоценного корма путем ферментации служит примером такого рода процесса, уже широко используемого человеком. Другой пример — разведение детритоядных рыб на Востоке.[ …]
Ресурсы биологические — организмы, которые являются или могут быть объектами промысла; все живые средообразующие компоненты биосферы (продуценты, консументы, редуценты). Они относятся к категории исчерпаемых возобновимых природных ресурсов. Различают растительные ресурсы, ресурсы животного мира, охотничьи, пастбищные и др. Так, тайга богата охотничьи-промысловыми ресурсами (пушнина, мясо, шкуры ценных животных), лесными (тайга только европейской части России имеет запасы древесины около 5,0 млрд м3), а также пищевыми ресурсами (ягоды, грибы, кедровые орехи и др.). Особо выделяют генетические ресурсы, т. е. наследственная генетическая информация, заключенная в генетическом коде живых существ.[ …]
В них объединены те случаи, когда одно животное поедает другое полностью (например, сова, охотящаяся за мышью), а также те, когда хищник потребляет только часть жертвы, давая ей возможность восстановиться с тем, чтобы использовать ее вновь (пастбищный тип питания). Растительноядных и плотоядных животных мы также рассматриваем вместе. Особое внимание уделено пищевым отношениям между животными и растениями из-за тех тонких приспособлений, которые характеризуют реакцию растений на выедание их животными. Поведение отдельных хищников оказывает глубокое воздействие на динамику популяций как хищника, так и жертвы и анализ последствий пищевых отношений между ними составляет содержание гл. 10. Этот раздел экологии наиболее тесно связан с проблемами управления природными ресурсами—проблемой повышения продуктивности (это может относиться к популяциям рыб, китов, сенокосным угодьям или степям) и проблемой биологического и химического контроля за сельскохозяйственными вредителями и сорняками.[ …]
Видимо, именно поэтому микробиологи, как правило, отмечают, что численность бактериальных популяций в морской воде очень низка. Правда, надо признать, что методы подсчета бактерий и измерения их активности в природе весьма примитивны и далеко не точны.[ …]
Обменный фонд образуется за счет веществ, которые возвращаются в круговорот двумя основными путями: 1) в результате первичной экскреции животными и 2) при разложении детрита микроорганизмами. Если оба пути замыкания обменного фонда реализуются в одной экосистеме, то первый из них доминирует, например, в планктоне и других сообществах, где основной поток энергии идет через пастбищную пищевую цепь; второй путь преобладает в степях, лесах умеренной зоны и других сообществах, в которых основной поток энергии направлен через детритную пищевую цепь.[ …]
В настоящее время известно, что наши представления о механизме формирования первичной продукции в Океане были неполными, а ее количественные характеристики — заниженными. По крайней мере для олиготрофных тропических акваторий Океана с температурой воды выше 20 °С доказано существование не одного, а двух путей создания первичной продукции — путей поступления органического вещества (энергии) в пищевые цепи сообщества (рис. 35). Первый путь — фитопланктонный. Это хорошо известная «пастбищная» пищевая цепь: фитопланктон — растительноядный зоопланктон — хищный планктон — нектон.[ …]
Эти организмы, практически «прописанные» в локальной экосистеме и функционирующие только в пределах, способны обеспечивать протекание всех процессов необходимых для существования круговорота биогенов в пределах локальной экосистемы. При такой ситуации можно предположить, что консументы, формирующие пастбищные цепи, выполняют в круговороте биогенов в локальных экосистемах, по-видимому, какие-то необходимые, но вторичные, подчиненные функции. Такими функциями, как было допущено ранее, могут быть функции стабилизации круговорота веществ в случае различных флуктуаций продукции продуцентов. Флуктуации продуцентов могут возникать из-за колебания масштабов потоков внешней энергии: солнечной радиации, количества осадков, концентрации диоксида углерода в атмосфере и др. В случае гашения таких нерегулярных колебаний метаболической мощности локальных экосистем консумента-ми пастбищных цепей устраняется необходимость соответствующих колебаний в функционировании основных групп организмов, осуществляющих процессы круговорота, что обеспечивает стабилизацию био-химических циклов биогенов.[ …]
Особую опасность для животных (и людей) представляет загрязнение среды бациллами (спорами) сибирской язвы. Споры возбудителя сибирской язвы устойчивы к воздействию факторов природной среды. Они сохраняются в грунте десятки лет и содержат потенциальную угрозу заражения животных и людей. При разливах полых и ливневых вод, размывающих почвы, проведении мелиоративных работ, строительстве дорог споры возбудителя сибирской язвы из глубоких слоев грунта могут быть вынесены на поверхность земли. Био-геохимическая пищевая цепь почва -»растения -> животные -> человек загрязняется, и риск возникновения вспышек эпизоотий и эпидемий сибирской язвы резко возрастает. Охрана пастбищных биогеоценозов и их окружения от бактериальных загрязнений представляет собой острую экологическую, ветеринарно-медицинскую, санитарно-гигиеническую проблему.[ …]
В случае ацидофикации водотока изменения, происходящие в его экосистеме, во многом имеют другую направленность. Несмотря на то, что биологическое разнообразие экосистемы уменьшается, общая структура речного континуума сохраняется. При этом подавляются процессы деструкции органического вещества бактериями и значительно уменьшается биомасса первичных консументов, что часто приводит к увеличению биомассы и усложнению пространственной структуры перифитона. Резко увеличивается роль вторичных консументов, среди которых доминируют хищные личинки водных насекомых. Многие из них имеют многолетний жизненный цикл и могут быть отнесены к r-стратегам. В целом ацидофикация приводит к преобладанию пастбищных пищевых цепей, снижению скорости деструкции органического вещества и увеличению отношения Р/R и К2 экосистемы и, следовательно, вызывает сдвиг функционирования экологической системы водотока к равновесному состоянию.[ …]
Установлено, что одни элементы наиболее прочно удерживаются в живом веществе и почве (азот, фосфор, калий и кальций), а другие интенсивно выщелачиваются и выносятся в реки и моря (хлор, магний, сера). В зависимости от климатических условий, растительного покрова и естественного дренажа местности эта характеристика меняется. К группе активных «путешественников» следует отнести бор, бром, серу, фтор, хлор, а « «ленивцам» — калий, кремний, медь, никель, фосфор и особенно алюминий и железо. Чтобы избежать опасности нарушения природных биологических круговоротов, хозяйственную деятельность человека необходимо планировать с учетом цикличности природных процессов. Особенно тщательно их следует учитывать в земледелии, пастбищном животноводстве, водоснабжении и навигации. Нельзя забывать, что чуткость почвы к вмешательству человека огромна. Распашка, внесение минеральных удобрений, загрязнение нефтью и тяжелыми металлами весьма обедняют ее фауну. В результате нарушаются и даже полностью выпадают звенья нормальных пищевых цепей и биогеохимических циклов.[ …]
При хорошем уходе молочные козы многих пород дают до 1000 кг высококачественного молока за 10 месяцев лактации, а например, зааненская до 3500 кг (рекорд). Уход за ними намного легче, чем за коровой, и производство козьего молока обходится намного дешевле, чем коровьего. Так, затраты корма на производство 1 ц козьего молока в 1,75 раза меньше, чем на производство 1 ц коровьего (соответственно 0,6 и 1,05 ц корм, ед.), а затраты труда меньше в 2,5 раза (соответственно 3 и 7,5 чел./ч). Коза в 2,3 раза дает больше молока на 1 ц живой массы, чем корова (соответственно 18,2 и 8 ц). Поэтому себестоимость козьего молока почти в 2 раза ниже коровьего, а рентабельность гораздо выше. Низкие затраты труда по уходу за козой, невысокий расход кормов позволяют содержать их пожилым людям. Козы хорошо акклиматизируются в различных климатических условиях. Они поедают значительно больше различных растений, чем любое другое домашнее животное, хорошо используют овощи, пищевые отходы и пастбищную растительность. Козы плодовиты и скороспелы. Продолжительность плодоношения 150 дней, хозяйственного использования 7-9 лет. При правильном кормлении и хорошем уходе козы редко болеют, отличаются чистоплотностью, легко доятся, устойчивы к заболеванию маститом.[ …]
Среди консументов могут быть выделены монофаги (животные, питающиеся одним видом жертвы), олигофаги (питающиеся небольшим числом видов жертвы) и полифаги (питающиеся многими видами жертвы). Для удобства животных нередко подразделяют на специалистов (в широком смысле это — монофаги и олигофаги) и универсалов (полифаги). Примеры видов, являющихся монофагами, олигофагами и полифагами, можно найти среди растительноядных организмов, паразитоидов и истинных хищников. Но среди консументов разных типов распространение животных с различной шириной спектра питания различается. Конечно, и среди истинных хищников есть виды, которые специализируются на строго определенной пище, например коршун-слизнеед (Но81гаНатиз зос1аЬШ8) питается почти исключительно слизнями рода Ротасеа. Но у большинства истинных хищников спектр питания относительно широк. Вместе с тем паразитоиды, как правило, специализированы и часто бывают монофагами, тогда как растительноядные организмы достаточно полно представлены во всех категориях. Однако если растительноядные организмы с пастбищным и «хищным» типами питания обычно имеют широкие пищевые спектры, то «паразитические» растительноядные организмы очень часто высокоспеци-ализированы. Например, Джензен апгеп, 1980) изучил в Коста-Рике 110 видов жуков, личинки которых питаются семенами двудольных растений, и обнаружил, что 83 из них поражают только один вид растений, 14 — только два, 9 — три, 2— четыре и по одному виду нападают на шесть и восемь растений. И это — несмотря на то что в районе произрастало 975 видов растений.[ …]
Источник: ru-ecology.info
В природе любой вид, популяция и даже отдельная особь живут не изолированно друг от друга и среды своего обитания, а, напротив, испытывают многочисленные взаимные влияния. Биотические сообщества или биоценозы — сообщества взаимодействующих живых организмов, представляющие собой устойчивую систему, связанную многочисленными внутренними связями, с относительно постоянной структурой и взаимообусловленным набором видов.
Для биоценоза характерны определенные структуры: видовая, пространственная и трофическая.
Органические компоненты биоценоза неразрывно связаны с неорганическими — почвой, влагой, атмосферой, образуя вместе с ними устойчивую экосистему — биогеоценоз.
Биогеноценоз – саморегулирующаяся экологическая система, образованная совместно обитающими и взаимодействующими между собой и с неживой природой, популяциями разных видов в относительно однородных условиях среды.
— функциональные системы, включающие в себя сообщества живых организмов разных видов и их среду обитания. Связи между компонентами экосистемы возникают, прежде всего, на основе пищевых взаимоотношений и способов получения энергии.
Экосистема
— совокупность видов растений, животных, грибов, микроорганизмов, взаимодействующих между собой и с окружающей средой таким образом, что такое сообщество может сохраняться и функционировать необозримо длительное время. Биотическое сообщество (биоценоз) состоит из сообщества растений (фитоценоз), животных (зооценоз), микроорганизмов (микробоценоз).
Все организмы Земли и среда их обитания также представляют собой экосистему высшего ранга — биосферу, обладающую устойчивостью и другими свойствами экосистемы.
Существование экосистемы возможно благодаря постоянному притоку энергии извне — таким источником энергии, как правило, является солнце, хотя не для всех экосистем это справедливо. Устойчивость экосистемы обеспечивается прямыми и обратными связями между ее компонентами, внутренним круговоротом веществ и участием в глобальных круговоротах.
Учение о биогеоценозах разработано В.Н. Сукачевым. Термин «экосистема» введен в употребление английским геоботаником А. Тенсли в 1935 г., термин «биогеоценоз» — академиком В.Н. Сукачевым в 1942 г. В биогеоценозе обязательно наличие в качестве основного звена растительного сообщества (фитоценоз), обеспечивающего потенциальную бессмертность биогеоценоза за счет энергии, вырабатываемой растениями. Экосистемы могут не содержать фитоценоз.
Фитоценоз
— растительное сообщество, исторически сложившееся в результате сочетания взаимодействующих растений на однородном участке территории.
Его характеризуют:
— определенный видовой состав,
— жизненные формы,
— ярусность (надземная и подземная),
— обилие (частота встречаемости видов),
— размещение,
— аспект (внешний вид),
— жизненность,
— сезонные изменения,
— развитие (смена сообществ).
Ярусность (этажность)
— один из характерных признаков растительного сообщества, заключающийся как бы в поэтажном его разделении как в надземном, так и в подземном пространстве.
Надземная ярусность позволяет лучше использовать свет, а подземная — воду и минеральные вещества. Обычно в лесу можно выделить до пяти ярусов: верхний (первый) — высокие деревья, второй — невысокие деревья, третий — кустарники, четвертый — травы, пятый — мхи.
Подземная ярусность — зеркальное отражение надземной: глубже всех уходят корни деревьев, близ поверхности почвы расположены подземные части мхов.
По способу получения и использования питательных веществ все организмы делятся на автотрофы и гетеротрофы. В природе возникает непрерывный круговорот биогенных веществ, необходимых для жизни. Химические вещества извлекаются автотрофами из окружающей среды и через гетеротрофы вновь в нее возвращаются. Этот процесс принимает очень сложные формы. Каждый вид использует лишь часть содержащейся в органическом веществе энергии, доводя его распад до определенной стадии. Таким образом, в процессе эволюции в экологических системах сложились цепи и сети питания.
Большинство биогеоценозов имеют сходную трофическую структуру. Основу их составляют зеленые растения — продуценты. Обязательно присутствуют растительноядные и плотоядные животные: потребители органического вещества — консументы и разрушители органических остатков — редуценты.
Количество особей в пищевой цепи последовательно уменьшается, численность жертв больше численности их потребителей, так как в каждом звене пищевой цепи при каждом переносе энергии 80—90% ее теряется, рассеиваясь в виде теплоты. Поэтому число звеньев в цепи ограничено (3—5).
Видовое разнообразие биоценоза представлено всеми группами организмов — продуцентами, консументами и редуцентами.
Нарушение какого-либо звена в цепи питания вызывает нарушение биоценоза в целом. Например, вырубка леса приводит к изменению видового состава насекомых, птиц, а, следовательно, и зверей. На безлесном участке будут складываться другие цепи питания и сформируется другой биоценоз, что займет не один десяток лет.
Цепь питания (трофическая или пищевая)
— взаимосвязанные виды, последовательно извлекающие органическое вещество и энергию из исходного пищевого вещества; при этом каждое предыдущее звено цепи является пищей для последующего.
Цепи питания состоят из нескольких звеньев, в них включаются растения, растительноядные животные, хищники и паразиты.
Цепи питания в каждом природном участке с более или менее однородными условиями существования составлены комплексами взаимосвязанных видов, питающимися друг другом и образующими самоподдерживающуюся систему, в которой осуществляется круговорот веществ и энергии.
— Продуценты — автотрофные организмы (в основном зеленые растения) — единственные производители органического вещества на Земле. Богатое энергией органическое вещество в процессе фотосинтеза синтезируется из бедных энергией неорганических веществ (Н20 и С02).
— Консументы — растительноядные и плотоядные животные, потребители органического вещества. Консументы могут быть растительноядными, когда они непосредственно используют продуценты, или плотоядными, когда они питаются другими животными. В цепи питания они чаще всего могут иметь порядковый номер с I по IV.
— Редуценты — гетеротрофные микроорганизмы (бактерии) и грибы — разрушители органических остатков, деструкторы. Их еще называют санитарами Земли.
Трофический (пищевой) уровень — совокупность организмов, объединяемых типом питания. Представление о трофическом уровне позволяет понять динамику потока энергии в экосистеме.
- первый трофический уровень всегда занимают продуценты (растения),
- второй — консументы I порядка (растительноядные животные),
- третий — консументы II порядка — хищники, питающиеся растительноядными животными),
- четвертый — консументы III порядка (вторичные хищники).
Различают следующие виды пищевых цепей:
— в пастбищной цепи (цепи выедания) основным источником пищи служат зеленые растения. Например: трава —> насекомые —> земноводные —> змеи —> хищные птицы.
— детритные цепи (цепи разложения) начинаются с детрита — отмершей биомассы. Например: листовой опад —> дождевые черви —> бактерии. Особенностью детритных цепей является также то, что в них часто продукция растений не потребляется непосредственно растительноядными животными, а отмирает и минерализуется сапрофитами. Детритные цепи характерны также для экосистем океанических глубин, обитатели которых питаются мертвыми организмами, опустившимися вниз из верхних слоев воды.
Особенностью пищевых цепей паразитов является то, что они могут начинаться как с продуцентов (яблоня —> щитовка —> наездник), так и с консументов (корова—> паразитические черви —> простейшие —> бактерии —> вирусы).
Сети питания
— сложившиеся в процессе эволюции взаимоотношения между видами в экологических системах, при которых многие компоненты питаются разными объектами и сами служат пищей различным членам экосистемы. Упрощенно пищевую сеть можно представить как систему переплетающихся пищевых цепей.
Организмы разных пищевых цепей, получающие пищу через равное число звеньев этих цепей, находятся на одном трофическом уровне. В то же время разные популяции одного и того же вида, входящие в различные пищевые цепи, могут находиться на разных трофических уровнях. Соотношение различных трофических уровней в экосистеме можно изобразить графически в виде экологической пирамиды.
Экологическая пирамида
— способ графического отображения соотношения различных трофических уровней в экосистеме — бывает трех типов:
• пирамида численности отражает численность организмов на каждом трофическом уровне;
• пирамида биомасс отражает биомассу каждого трофического уровня;
• пирамида энергии показывает количество энергии, прошедшее через каждый трофический уровень в течение определенного промежутка времени.
Правило экологической пирамиды
— закономерность, отражающая прогрессивное уменьшение массы (энергии, числа особей) каждого последующего звена пищевой цепи.
Пирамида численности
— экологическая пирамида, отражающая число особей на каждом пищевом уровне. В пирамиде чисел не учитываются размеры и масса особей, продолжительность жизни, интенсивность обмена веществ, однако всегда прослеживается главная тенденция — уменьшение числа особей от звена к звену. Например, в степной экосистеме численность особей распределяется так: продуценты — 150000, травоядные консументы — 20000, плотоядные консументы — 9000 экз./ар. Биоценоз луга характеризуется следующей численностью особей на площади 4000 м2: продуценты — 5 842 424, растительноядные консументы I порядка — 708 624, плотоядные консументы II порядка — 35 490, плотоядные консументы III порядка — 3.
Пирамида биомасс
— закономерность, согласно которой количество растительного вещества, служащего основой цепи питания (продуцентов), примерно в 10 раз больше, чем масса растительноядных животных (консументов I порядка), а масса растительноядных животных в 10 раз больше, чем плотоядных (консументов II порядка), т. е. каждый последующий пищевой уровень имеет массу в 10 раз меньшую, чем предыдущий. В среднем из 1000 кг растений образуется 100 кг тела травоядных животных. Хищники, поедающие травоядных, могут построить 10 кг своей биомассы, вторичные хищники — 1 кг.
Пирамида энергии
выражает закономерность, согласно которой поток энергии постепенно уменьшается и обесценивается при переходе от звена к звену в цепи питания. Так, в биоценозе озера зеленые растения — продуценты — создают биомассу, содержащую 295,3 кДж/см2, консументы I порядка, потребляя биомассу растений, создают свою биомассу, содержащую 29,4 кДж/см2; консументы II порядка, используя в пищу консументов I порядка, создают свою биомассу, содержащую 5,46 кДж/см2. Потеря энергии при переходе от консументов I порядка к консументам II порядка, если это теплокровные животные, увеличивается. Это объясняется тем, что у данных животных много энергии уходит не только на построение своей биомассы, но и на поддержание постоянства температуры тела. Если сравнить выращивание теленка и окуня, то одинаковое количество затраченной пищевой энергии даст 7 кг говядины и лишь 1 кг рыбы, так как теленок питается травой, а окунь-хищник — рыбой.
Таким образом, первые два типа пирамид имеют ряд существенных недостатков:
— Построение пирамиды численности может быть затруднено, если разброс численности организмов разных уровней велик (например, 500 тыс. злаков в основании пирамиды может соответствовать один конечный хищник). Кроме того, пирамида может оказаться перевернутой (в том случае, если продуцент очень крупный, или если большое число паразитов питаются на немногочисленных консументах).
— Пирамида биомасс отражает состояние экосистемы на момент отбора пробы и, следовательно, показывает соотношение биомассы в данный момент и не отражает продуктивность каждого трофического уровня (т. е. его способность образовывать биомассу в течение определенного промежутка времени). Поэтому в том случае, когда в число продуцентов входят быстрорастущие виды, пирамида биомасс может оказаться перевернутой.
— Пирамида энергии позволяет сравнить продуктивность различных трофических уровней, поскольку учитывает фактор времени. Кроме того, она учитывает разницу в энергетической ценности различных веществ (например, 1 г жира дает почти в два раза больше энергии, чем 1 г глюкозы). Поэтому пирамида энергии всегда суживается кверху и никогда не бывает перевернутой.
Экологическая пластичность
— степень выносливости организмов или их сообществ (биоценозов) к воздействию факторов среды. Экологически пластичные виды имеют широкую норму реакции, т. е. широко приспособлены к разной среде обитания (рыбы колюшка и угорь, некоторые простейшие живут как в пресных, так и в соленых водах). Узкоспециализированные виды могут существовать лишь в определенной среде: морские животные и водоросли — в соленой воде, речные рыбы и растения лотос, кувшинка, ряска обитают только в пресной воде.
В целом экосистема (биогеоценоз) характеризуется следующими показателями:
— видовым разнообразием,
— плотностью видовых популяций,
— биомассой.
Биомасса
— общее количество органического вещества всех особей биоценоза или вида с заключенной в нем энергией. Биомассу выражают обычно в единицах массы в пересчете на сухое вещество единицы площади или объема. Биомассу можно определить отдельно для животных, растений или отдельных видов. Так, биомасса грибов в почве составляет 0,05-0,35 т/га, водорослей — 0,06-0,5, корней высших растений — 3,0-5,0, дождевых червей — 0,2-0,5, позвоночных животных — 0,001-0,015 т/га.
В биогеоценозах различают первичную и вторичную биологическую продуктивность:
ü Первичная биологическая продуктивность биоценозов — общая суммарная продуктивность фотосинтеза, представляющая собой результат деятельности автотрофов — зеленых растений, например, сосновый лес 20— 30-летнего возраста за год производит 37,8 т/га биомассы.
ü Вторичная биологическая продуктивность биоценозов — общая суммарная продуктивность гетеротрофных организмов (консументов), которая образуется за счет использования веществ и энергии, накопленных продуцентами.
Каждый вид на Земле занимает определенный ареал, так как он способен существовать лишь в определенных условиях среды. Однако условия обитания в рамках ареала одного вида могут существенно отличаться, что приводит к распаду вида на элементарные группировки особей — популяции.
Популяция
— совокупность особей одного вида, занимающих обособленную территорию в пределах ареала вида (с относительно однородными условиями обитания), свободно скрещивающихся друг с другом (имеющих общий генофонд) и изолированных от других популяций данного вида, обладающих всеми необходимыми условиями для поддержания своей стабильности длительное время в меняющихся условиях среды. Важнейшими характеристиками популяции являются ее структура (возрастной, половой состав) и динамика численности.
Под демографической структурой популяции понимают ее половой и возрастной состав.
Пространственная структура популяции — это особенности размещения особей популяции в пространстве.
Возрастная структура популяции связана с соотношением особей различных возрастов в популяции. Особи одного возраста объединяют в когорты — возрастные группы.
В возрастной структуре популяций растений выделяют следующие периоды:
— латентный — состояние семени;
— прегенеративный (включает состояния проростка, ювенильного растения, имматурного и виргинильного растений);
— генеративный (обычно подразделяется на три подпериода — молодые, зрелые и старые генеративные особи);
— постгенеративный (включает состояния субсенильного, сенильного растений и фазу отмирания).
Принадлежность к определенному возрастному состоянию определяется по биологическому возрасту — степени выраженности определенных морфологических (например, степень расчлененности сложного листа) и физиологических (например, способность дать потомство) признаков.
В популяциях животных также можно выделить различные возрастные стадии. Например, насекомые, развивающиеся с полным метаморфозом, проходят стадии:
— яйца,
— личинки,
— куколки,
— имаго (взрослого насекомого).
Характер возрастной структуры популяции зависит от типа кривой выживания, свойственной данной популяции.
Кривая выживания отражает уровень смертности в различных возрастных группах и представляет собой снижающуюся линию:
- Если уровень смертности не зависит от возраста особей, отмирание особей происходит в данном типе равномерно, коэффициент смертности остается постоянным на протяжении всей жизни (тип I). Такая кривая выживания свойственна видам, развитие которых происходит без метаморфоза при достаточной устойчивости рождающегося потомства. Этот тип принято называть типом гидры — для нее свойственна кривая выживания, приближающаяся к прямой линии.
- У видов, для которых роль внешних факторов в смертности невелика, кривая выживания характеризуется небольшим понижением до определенного возраста, после которого происходит резкое падение вследствие естественной (физиологической) смертности (тип II). Близкий к этому типу характер кривой выживания свойствен человеку (хотя кривая выживания человека несколько более пологая и является чем-то средним между типами I и II). Этот тип носит название типа дрозофилы: именно его демонстрируют дрозофилы в лабораторных условиях (не поедаемые хищниками).
- Для очень многих видов характерна высокая смертность на ранних стадиях онтогенеза. У таких видов кривая выживания характеризуется резким падением в области младших возрастов. Особи, пережившие «критический» возраст, демонстрируют низкую смертность и доживают до старших возрастов. Тип носит название типа устрицы (тип III).
Половая структура популяции
— соотношение полов имеет прямое отношение к воспроизводству популяции и ее устойчивости.
Выделяют первичное, вторичное и третичное соотношение полов в популяции:
— Первичное соотношение полов определяется генетическими механизмами — равномерностью расхождения половых хромосом. Например, у человека XY-хромосомы определяют развитие мужского пола, а XX — женского. В этом случае первичное соотношение полов 1:1, т. е. равновероятно.
— Вторичное соотношение полов — это соотношение полов на момент рождения (среди новорожденных). Оно может существенно отличаться от первичного по целому ряду причин: избирательность яйцеклеток к сперматозоидам, несущим Х- или Y-хромосому, неодинаковой способностью таких сперматозоидов к оплодотворению, различными внешними факторами. Например, зоологами описано влияние температуры на вторичное соотношение полов у рептилий. Аналогичная закономерность характерна и для некоторых насекомых. Так, у муравьев оплодотворение обеспечивается при температуре выше 20 °С, а при более низких температурах откладываются неоплодотворенные яйца. Из последних вылупляются самцы, а из оплодотворенных — преимущественно самки.
— Третичное соотношение полов — соотношение полов среди взрослых животных.
Пространственная структура популяции отражает характер размещения особей в пространстве.
Выделяют три основных типа распределения особей в пространстве:
— единообразное или равномерное (особи размещены в пространстве равномерно, на одинаковых расстояниях друг от друга); встречается в природе редко и чаще всего вызвано острой внутривидовой конкуренцией (например, у хищных рыб);
— конгрегационное или мозаичное («пятнистое», особи размещаются в обособленных скоплениях); встречается намного чаше. Оно связано с особенностями микросреды или поведения животных;
— случайное или диффузное (особи распределены в пространстве случайным образом) — можно наблюдать только в однородной среде и только у видов, которые не обнаруживают никакого стремления к объединению в группы (например, у жука в муке).
Численность популяции обозначается буквой N. Отношение прироста N к единице времени dN / dt выражает мгновенную скорость изменения численности популяции, т. е. изменение численности в момент времени t. Прирост популяции зависит от двух факторов — рождаемости и смертности при условии отсутствия эмиграции и иммиграции (такая популяция называется изолированной). Разность рождаемости b и смертности d и представляет собой коэффициент прироста изолированной популяции:
r = b—d.
Устойчивость популяции
— это ее способность находиться в состоянии динамического (т. е. подвижного, изменяющегося) равновесия со средой: изменяются условия среды — изменяется и популяция. Одним из важнейших условий устойчивости является внутреннее разнообразие. Применительно к популяции это механизмы поддержания определенной плотности популяции.
Выделяют три типа зависимости численности популяции от ее плотности.
Первый тип (I) — самый распространенный, характеризуется уменьшением роста популяции при увеличении ее плотности, что обеспечивается различными механизмами. Например, для многих видов птиц характерны снижение рождаемости (плодовитости) при увеличении плотности популяции; увеличение смертности, снижение сопротивляемости организмов при повышенной плотности популяции; изменение возраста наступления половой зрелости в зависимости от плотности популяции.
Второй тип (II) характеризуется постоянным темпом роста численности, который резко падает при достижении максимальной численности (лемминги при достижении максимальной плотности начинают массово мигрировать; достигнув моря, многие тонут). Одним из важнейших факторов поддержания численности популяций является внутривидовая конкуренция. Она может проявляться в различных формах: от борьбы за места для гнездования до каннибализма. Межвидовые отношения также играют важную роль. Отношения паразит — хозяин и хищник — жертва во многом зависят от плотности: распространение болезней происходит быстрее в популяциях с высокой плотностью, которая выступает эпидемиологическим фактором.
Третий тип (III) характерен для популяций, в которых отмечается «эффект группы», т. е. определенная оптимальная плотность популяции способствует лучшему выживанию, развитию, жизнедеятельности всех особей, что присуще большинству групповых и социальных животных. Например, для возобновления популяций разнополых животных как минимум необходима плотность, обеспечивающая достаточную вероятность встречи самца и самки.
Таким образом, оптимальная численность и плотность популяции поддерживаются благодаря внутрипопуляционным механизмам (увеличение/уменьшение рождаемости и смертности, изменение возраста наступления половой зрелости, внутривидовая конкуренция) и междвидовым механизмам (взаимоотношения хищник — жертва и паразит — хозяин).
Тематические задания
А1. Биогеоценоз образован
1) растениями и животными
2) животными и бактериями
3) растениями, животными, бактериями
4) территорией и организмами
А2. Потребителями органического вещества в лесном биогеоценозе являются
1) ели и березы
2) грибы и черви
3) зайцы и белки
4) бактерии и вирусы
А3. Продуцентами в озере являются
1) лилии
2) головастики
3) раки
4) рыбы
А4. Процесс саморегуляции в биогеоценозе влияет на
1) соотношение полов в популяциях разных видов
2) численность мутаций, возникающих в популяциях
3) соотношение хищник – жертва
4) внутривидовую конкуренцию
А5. Одним из условий устойчивости экосистемы может служить
1) ее способность к изменениям
2) разнообразие видов
3) колебания численности видов
4) стабильность генофонда в популяциях
А6. К редуцентам относятся
1) грибы
2) лишайники
3) мхи
4) папоротники
А7. Если общая масса полученной потребителем 2-го порядка равна 10 кг, то какова была совокупная масса продуцентов, ставших источником пищи для данного потребителя?
1) 1000 кг
2) 500 кг
3) 10000 кг
4) 100 кг
А8. Укажите детритную пищевую цепь
1) муха – паук – воробей – бактерии
2) клевер – ястреб – шмель – мышь
3) рожь – синица – кошка – бактерии
4) комар – воробей – ястреб – черви
А9. Исходным источником энергии в биоценозе является энергия
1) органических соединений
2) неорганических соединений
3) Солнца
4) хемосинтеза
А10. Взаимовыгодными можно считать отношения между липой и:
1) зайцами
2) пчелами
3) дроздами-рябинниками
4) волками
А11. В одной экосистеме можно встретить дуб и
1) суслика
2) кабана
3) жаворонка
4) синий василек
А12. Сети питания – это:
1) связи между родителями и потомством
2) родственные (генетические) связи
3) обмен веществ в клетках организма
4) пути передачи веществ и энергии в экосистеме
А13. Экологическая пирамида чисел отражает:
1) соотношение биомасс на каждом трофическом уровне
2) соотношение масс отдельного организма на разных трофических уровнях
3) структуру пищевой цепи
4) разнообразие видов на разных трофических уровнях
А14. Доля энергии, передаваемая на следующий трофический уровень, составляет приблизительно:
1) 10%
2) 30%
3) 50%
4) 100%
Источник: biology100.ru