Виды мейоза
| |
Мейоз
Основные понятия и определения
Мейозом называется особый способ деления эукариотических клеток, при котором исходное число хромосом уменьшается в 2 раза (от древнегреч. «мейон» – меньше – и от «мейозис» – уменьшение). Часто уменьшение числа хромосом называется редукцией.
Исходное число хромосом в мейоцитах (клетках, вступающих в мейоз) называется диплоидным хромосомным числом (2n) Число хромосом в клетках, образовавшихся в результате мейоза, называется гаплоидным хромосомным числом (n).
Минимальное число хромосом в клетке называется основным числом (x). Основному числу хромосом в клетке соответствует и минимальный объем генетической информации (минимальный объем ДНК), который называется геном. Количество геномов в клетке называется геномным числом (Ω). У большинства многоклеточных животных, у всех голосеменных и многих покрытосеменных растений понятие гаплоидности–диплоидности и понятие геномного числа совпадают. Например, у человека n=x=23 и 2n=2x=46.
Главной особенностью мейоза является конъюгация (спаривание) гомологичных хромосом с последующим расхождением их в разные клетки. Мейотическое распределение хромосом по дочерним клеткам называется сегрегацией хромосом.
Краткая история открытия мейоза
Отдельные фазы мейоза у животных описал В. Флемминг (1882), а у растений – Э.Страсбургер (1888), а затем российский ученый В.И. Беляев. В это же время (1887) А. Вайсман теоретически обосновал необходимость мейоза как механизма поддержания постоянного числа хромосом. Первое подробное описание мейоза в ооцитах кролика дал Уиниуортер (1900). Изучение мейоза продолжается до сих пор.
Общий ход мейоза
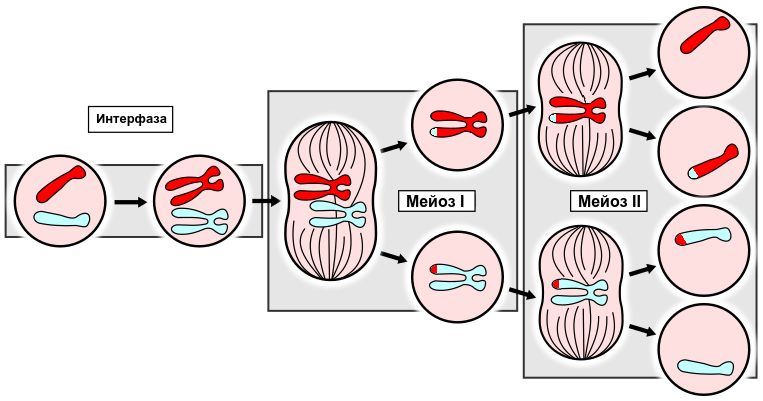
Типичный мейоз состоит из двух последовательных клеточных делений, которые соответственно называются мейоз I и мейоз II. В первом делении происходит уменьшение числа хромосом в два раза, поэтому первое мейотическое деление называют редукционным, реже – гетеротипным. Во втором делении число хромосом не изменяется; такое деление называют эквационным (уравнивающим), реже – гомеотипным. Выражения «мейоз» и «редукционное деление» часто используют как синонимы.
Интерфаза
Предмейотическая интерфаза отличается от обычной интерфазы тем, что процесс репликации ДНК не доходит до конца: примерно 0,2…0,4 % ДНК остается неудвоенной. Таким образом, деление клетки начинается на синтетической стадии клеточного цикла. Поэтому мейоз образно называют преждевременным митозом. Однако в целом, можно считать, что в диплоидной клетке (2n) содержание ДНК составляет 4с.
При наличии центриолей происходит их удвоение таким образом, что в клетке имеется две диплосомы, каждая из которых содержит пару центриолей.
Первое деление мейоза (редукционное деление, или мейоз I)
Сущность редукционного деления заключается в уменьшении числа хромосом в два раза: из исходной диплоидной клетки образуется две гаплоидные клетки с двухроматидными хромосомами (в состав каждой хромосомы входит 2 хроматиды).
Профаза 1 (профаза первого деления) состоит из ряда стадий:
Лептотена (стадия тонких нитей). Хромосомы видны в световой микроскоп в виде клубка тонких нитей. Раннюю лептотену, когда нити хромосом видны еще очень плохо, называют пролептотена.
Зиготена (стадия сливающихся нитей).
оисходит конъюгация гомологичных хромосом (от лат. conjugatio – соединение, спаривание, временное слияние). Гомологичные хромосомы (или гомологи) – это хромосомы, сходные между собой в морфологическом и генетическом отношении. У нормальных диплоидных организмов гомологичные хромосомы – парные: одну хромосому из пары диплоидный организм получает от матери, а другую – от отца. При конъюгации образуются биваленты. Каждый бивалент – это относительно устойчивый комплекс из одной пары гомологичных хромосом. Гомологи удерживаются друг около друга с помощью белковых синаптонемальных комплексов. Один синаптонемальный комплекс может связывать только две хроматиды в одной точке. Количество бивалентов равно гаплоидному числу хромосом. Иначе биваленты называются тетрады, так как в состав каждого бивалента входит 4 хроматиды.
Пахитена (стадия толстых нитей). Хромосомы спирализуются, хорошо видна их продольная неоднородность. Завершается репликация ДНК (образуется особая пахитенная ДНК). Завершается кроссинговер – перекрест хромосом, в результате которого они обмениваются участками хроматид.
Диплотена (стадия двойных нитей). Гомологичные хромосомы в бивалентах отталкиваются друг от друга. Они соединены в отдельных точках, которые называются хиазмы (от древнегреч. буквы χ – «хи»).
Диакинез (стадия расхождения бивалентов). Отдельные биваленты располагаются на периферии ядра.
Метафаза I (метафаза первого деления)
В прометафазе I ядерная оболочка разрушается (фрагментируется). Формируется веретено деления. Далее происходит метакинез – биваленты перемещаются в экваториальную плоскость клетки.
Анафаза I (анафаза первого деления)
Гомологичные хромосомы, входящие в состав каждого бивалента, разъединяются, и каждая хромосома движется в сторону ближайшего полюса клетки. Разъединения хромосом на хроматиды не происходит. Процесс распределения хромосом по дочерним клеткам называется сегрегация хромосом.
Телофаза I (телофаза первого деления)
Гомологичные двухроматидные хромосомы полностью расходятся к полюсам клетки. В норме каждая дочерняя клетка получает одну гомологичную хромосому из каждой пары гомологов. Формируются два гаплоидных ядра, которые содержат в два раза меньше хромосом, чем ядро исходной диплоидной клетки. Каждое гаплоидное ядро содержит только один хромосомный набор, то есть каждая хромосома представлена только одним гомологом. Содержание ДНК в дочерних клетках составляет 2с.
В большинстве случаев (но не всегда) телофаза I сопровождается цитокинезом.
Интеркинез
Интеркинез – это короткий промежуток между двумя мейотическими делениями. Отличается от интерфазы тем, что не происходит репликации ДНК, удвоения хромосом и удвоения центриолей: эти процессы произошли в предмейотической интерфазе и, частично, в профазе I.
Второе деление мейоза (эквационное деление, или мейоз II)
В ходе второго деления мейоза уменьшения числа хромосом не происходит. Сущность эквационного деления заключается в образовании четырех гаплоидных клеток с однохроматидными хромосомами (в состав каждой хромосомы входит одна хроматида).
Профаза II (профаза второго деления)
Не отличается существенно от профазы митоза. Хромосомы видны в световой микроскоп в виде тонких нитей. В каждой из дочерних клеток формируется веретено деления.
Метафаза II (метафаза второго деления)
Хромосомы располагаются в экваториальных плоскостях гаплоидных клеток независимо друг от друга. Эти экваториальные плоскости могут лежать в одной плоскости, могут быть параллельны друг другу или взаимно перпендикулярны.
Анафаза II (анафаза второго деления)
Хромосомы разделяются на хроматиды (как при митозе). Получившиеся однохроматидные хромосомы в составе анафазных групп перемещаются к полюсам клеток.
Телофаза II (телофаза второго деления)
Однохроматидные хромосомы полностью переместились к полюсам клетки, формируются ядра. Содержание ДНК в каждой из клеток становится минимальным и составляет 1с.
Типы мейоза и его биологическое значение
В общем случае в результате мейоза из одной диплоидной клетки образуется четыре гаплоидные клетки. При гаметном мейозе из образовавшихся гаплоидных клеток образуются гаметы. Этот тип мейоза характерен для животных. Гаметный мейоз тесно связан с гаметогенезом и оплодотворением. При зиготном и споровом мейозе образовавшиеся гаплоидные клетки дают начало спорам или зооспорам. Эти типы мейоза характерны для низших эукариот, грибов и растений. Споровый мейоз тесно связан со спорогенезом. Таким образом, мейоз – это цитологическая основа полового и бесполого (спорового) размножения.
Биологическое значение мейоза заключается в поддержании постоянства числа хромосом при наличии полового процесса. Кроме того, вследствие кроссинговера происходит рекомбинация – появление новых сочетаний наследственных задатков в хромосомах. Мейоз обеспечивает также комбинативную изменчивость – появление новых сочетаний наследственных задатков при дальнейшем оплодотворении.
Ход мейоза находится под контролем генотипа организма, под контролем половых гормонов (у животных), фитогормонов (у растений) и множества иных факторов (например, температуры).
Источник: megalektsii.ru
МЕЙОЗ (греч. meiosis уменьшение) — специальный тип клеточного деления, который обязательно проходят половые клетки во время их созревания.
присущ всем видам растительных и животных организмов, размножающихся половым путем, и его основные закономерности универсальны. Биол, назначение М. двояко. С одной стороны, для сохранения биол, вида необходимо, чтобы присущее ему число хромосом (диплоидное число, у человека равное 46) поддерживалось неизменным в ряду поколений организмов. С другой стороны, каждый организм образуется при слиянии двух половых клеток, мужской и женской (см. Оплодотворение). Следовательно, половые клетки в отличие от соматических клеток должны иметь уменьшенное вдвое (гаплоидное) число хромосом (см. Гаметы). Двукратное уменьшение (редукция) числа хромосом при созревании гамет и обеспечивается в процессе М., который поэтому называется еще редукционным делением.
Кроме того, благодаря М. обеспечивается новая комбинация генов в гамете и возникающий в результате оплодотворения новый организм всегда уникален, а не является простой суммой черт своих родителей. Такая генетическая рекомбинация осуществляется в М. двумя путями. Во-первых, в исходной половой клетке (сперматоците или ооците), еще сохраняющей диплоидное число хромосом, происходит временное спаривание (конъюгация, синапсис) гомологичных родительских хромосом и обмен между ними идентичными хромосомными участками (кроссинговер, или внутрихромосомная рекомбинация, или рекомбинация сцепленных, т. е. находящихся в одной хромосоме, генов; рис. 1). Во-вторых, по завершении кроссинговера при первом делении мейотической клетки негомологичные хромосомы расходятся в дочерние клетки случайно, чем обеспечивается для каждой гаметы своя рекомбинация несцепленных, т. е. расположенных в разных хромосомах, генов, или межхромосомная рекомбинация (рис. 2).
Цитологически М. включает в себя два последовательных клеточных деления (первое и второе мейотическое деления) при одной дупликации хромосом, происходящей в интерфазе только первого деления. В результате из исходной мужской половой клетки (сперматогонии) возникают четыре гаметы — спермия, каждый из которых полноценен. Из женской половой клетки (оогонии) в зрелую гамету (яйцеклетку) развивается лишь один из четырех продуктов мейотических делений, остальные три образуют так наз. направительные (полярные) тельца, которые затем дегенерируют.
В соответствии с двумя делениями М. и с фазами каждого деления (см. Митоз) выделяют так наз. стадии М. Профазе I предшествует стадия пролептотены, во время к-рой в клеточном ядре после окончания периода синтеза ДНК происходит временная конденсация хромосом, в результате чего они приобретают вид плотно красящихся нерезко очерченных образований (прохромосом). Значение этого явления пока не ясно. Профазу I М., в отличие от профазы митоза, подразделяют на пять стадий: лептотену, зиготену, пахитену, диплотену и диакинез (рис. 3, 1—5).
В лептотене все пространство ядра заполнено клубком хромосом, которые имеют вид тонких длинных нитей, внешне не разделенных на хроматиды, хотя хроматиды и существуют.
В зиготене гомологичные хромосомы в отдельных, строго гомологичных участках, чаще всего в теломерном и центромерном районах, начинают спариваться (конъюгировать), хромосомы при этом укорачиваются. Спаривание хромосом является важным условием для осуществления обмена генетическим материалом — кроссинговера (см. Конъюгация хромосом).
В пахитене хромосомы-гомологи спарены по всей длине (биваленты). Их длина составляет всего 1/4—1/6 длины этих же хромосом в лептотене, они имеют четкое хромомерное строение. Половые хромосомы X и Y обычно конъюгируют отдельными участками. На этой стадии, особенно в ее конце, может быть подсчитано число бивалентов, соответствующее гаплоидному числу хромосом, и по специфическому хромомерному рисунку и размеру бивалента может быть проведена их индивидуальная идентификация (пахитенный анализ). Если в хромосомном наборе больше двух гомологичных хромосом (трисомия, тетрасомия и т.д.), их конъюгация дает мультиваленты (триваленты, тетраваленты и т. д.). В клетке полиплоидного организма (триплоидия, тетраплоидия и т. д.) все хромосомы в пахитене могут формировать мультиваленты. У полиплоидных организмов, когда в мейоците имеется четыре гомологичных хромосомы, их спаривание между собой может происходить по-разному. Гомологи, пришедшие от одного родителя, могут конъюгировать только между собой (аутосиндез). Конъюгация может происходить только между гомологами разных родителей (аллосиндез). В одной и той же клетке возможны спаривания обоего типа (ауто-аллосиндез). Хромосома, не имеющая гомолога (моносомия, межвидовые гибриды), будет проходить профазу М., не конъюгируя (унивалент). Аутологичные хромосомы (идентичные между собой по генному составу и структуре), которые могут образоваться в предмейозе в результате дополнительной редупликации, образуют при конъюгации аутобиваленты.
В диплотене происходит отталкивание гомологичных хромосом друг от друга, и в каждой из них начинают различаться сестринские хроматиды, в результате впервые в биваленте проявляется четырехнитчатая структура — тетрада (рис. 4).
В отдельных точках по длине бивалента гомологи остаются в соприкосновении, эти участки имеют вид перекрестов, называются хиазмами и являются цитол, проявлением кроссинговера. Число и локализация хиазм индивидуальны для каждого бивалента. На распределение хиазм влияет топография гетерохроматиновых районов в хромосоме, в таких районах хиазм меньше всего или вовсе нет. Разные биол, виды существенно различаются по хиазмообразованию. Имеются виды насекомых, у которых хиазм не бывает вообще (напр., у самцов дрозофилы) .
Профаза I М. заканчивается стадией диакинеза, в течение к-рой происходит еще большее расхождение гомологичных хромосом при дальнейшем укорочении бивалентов. В ходе этого процесса наблюдается смещение хиазм к концам хромосомы (терминализация хиазм), однако биваленты сохраняются. К концу диакинеза полностью растворяется ядерная оболочка, формируется веретено деления и начинается метафаза I (рис. 3, 6).
В метафазе биваленты располагаются в экваториальной плоскости клетки, при этом центромеры гомологичных хромосом каждого бивалента ориентированы к разным полюсам веретена. В анафазе I завершается распад бивалента и расхождение гомологичных хромосом в дочерние клетки. Этот вид деления называют редукционным (см. рис. 4). В каждом из гомологов сестринские хроматиды в анафазе I остаются связанными друг с другом, гомолог переходит в тело-фазу I в виде двунитчатой структуры (диады). Эти особенности отличают мейотическое деление I от митотического, поэтому его называют гетерокинезом. Интерфазное состояние, предшествующее мейотическому делению II и называемое интеркинезом, кратковременно, поскольку в нем нет синтеза ДНК. У отдельных видов организмов интеркинеза и профазы II не бывает. Профаза II, метафаза II, анафаза II и телофаза II следуют быстро одна за другой (рис. 3). В процессе деления II осуществляется расхождение сестринских хроматид в дочерние клетки. Эти хроматиды равноценны по генетическому содержанию, поэтому деление II называется эквационным (см. рис. 4.) У некоторых биол, видов деление I является эквационным, а деление II — редукционным. Четыре гаплоидных продукта одной и той же оогонии называют тетрадой; вследствие аномального М. продуктом одной гонии может быть одна половая зрелая клетка (монада).
М. у человека полностью соответствует описанной общей схеме. Между мужским и женским М. у человека имеются существенные различия по времени его протекания в онтогенезе и продолжительности стадий. У мужчин мейотические деления начинаются в пубертатном возрасте и продолжаются непрерывно в течение половозрелой жизни. Преобразование сперматогонии в зрелый спермий (см. Сперматогенез) занимает ок. 8—9 нед. Различные стадии М. у мужчин различаются по продолжительности, и с этим связана разная частота, с к-рой обнаруживаются на препаратах биопсийного материала яичек мейоциты разных стадий. Сравнительно редко встречаются зиготена и диплотена, чаще — лептотена, пахитена и диакинез. У женщины преобразование оогоний начинается в раннем эмбриональном периоде, и к 7-му месяцу внутриутробной жизни все оогонии переходят в первичные ооциты, достигая стадии диплотены. На этой стадии происходит деконденсация бивалентов, и ооциты остаются в неизменном состоянии до половозрелого возраста (стадия диктиотены). Мейотическому делению II они подвергаются с началом половозрелого периода жизни женщины, созревая в яйцеклетки по очереди в каждом овуляционном цикле. Деление II ооцита продолжается вплоть до оплодотворения, причем с яйцеклеткой остаются в связи три полярных тельца (см. Оогенез).
Стадии лептотены, зиготены и пахитены доступны изучению у лиц обоих полов. У обоих полов до пахитены индивидуальные биваленты выделить не удается. В пахитене, особенно поздней, некоторые индивидуальные биваленты можно различить по длине, положению центромеры, числу и размерам хромомер. Биваленты акроцентрических хромосом идентифицировать легче других по их связи с ядрышком. Полный пахитенный анализ усложнен тем, что получить на препаратах раздельно лежащие биваленты трудно. Половые хромосомы у мужчин, в отличие от половых хромосом женщин, конъюгируют концами; кроме того, они конденсированы и образуют в пахитене так наз. половой пузырек, поэтому их легко обнаружить.
Хромосомное исследование стадий диакинез — метафаза I удалось осуществить только у мужчин. При обычной окраске из 23 бивалентов с уверенностью могут быть выделены лишь XY бивалент и иногда бивалент IX, а также может быть проведено разделение всех бивалентов на группы А — G аналогично групповой идентификации хромосом в метафазе митоза (см. Кариотип). С помощью дифференциальных окрасок по методике Q и С, которые позволяют выявлять индивидуальную морфологию плеч хромосом и положение центромеры, в диакинезе — метафазе I идентифицированы все 23 бивалента и изучено число и локализация хиазм. В среднем на одну клетку приходится ок. 50 хиазм при разбросе чисел от 33 до 66. Хиазмы обнаружены во всех аутосомных бивалентах; число хиазм, приходящихся на один бивалент, колеблется от 1 до 5, увеличиваясь пропорционально длине хромосомы. X и Y хромосомы конъюгируют концами коротких плеч, и в этом биваленте хиазм не найдено. XY бивалент резко конденсирован и интенсивно окрашивается основными красителями (явление так наз. гетеропикноза). Иногда половые хромосомы видны как униваленты. В нормальном хромосомном наборе аутосомные униваленты чрезвычайно редки, мультиваленты также не обнаружены, хотя полиплоидные мейоциты I регулярно встречаются.
Изучение хромосом в медико-генетических целях в основном проводится на стадии диакинез — метафаза I; оно имеет диагностическое и прогностическое значение при хромосомных болезнях (см. Хромосомные болезни) и мужском бесплодии. Как правило, хромосомные болезни человека, обусловленные изменением числа хромосом, возникают вследствие нарушенного расхождения хромосом в мейотических делениях. Установлено, что в гаметогенезе (см.) происходит частичная элиминация гамет, несбалансированных по числу хромосом. Поэтому изучение М. дает ответ на вопрос о частоте нерасхождения разных хромосом в I и II делениях и частоте элиминации гамет, аномальных по тем или иным хромосомам. Поскольку хромосомы в М. конъюгируют строго гомологичными участками, изучение конфигурации спаренных хромосом в диакинезе — мета-фазе I дает дополнительный материал для суждения о происхождении лишних хромосом и о нарушении их структуры. Так, при трисомиях лишняя хромосома образует с двумя гомологами тривалент либо образуются бивалент и унивалент. Транслокации акроцентрических хромосом типа центрического слияния выглядят как триваленты, а Реципрокные транслокации — как квадриваленты разнообразной конфигурации. Иногда причиной мужского бесплодия является та или иная хромосомная аномалия, в т. ч. аутосомная, и изучение поведения хромосом в М. помогает постановке диагноза.
Механизмы М. во многом остаются неясными. Интенсивно проводятся морфологические, биохимические и генетические исследования механизмов перехода от митоза к М., конъюгации и кроссинговера, других этапов дифференцировки мейоцитов в гаметы. Отличия от митоза начинаются в предмейотической интерфазе, когда обнаруживается удлинение периода синтеза ДНК, незавершенность синтеза ДНК и гистонов к моменту наступления профазы. В зиготене наблюдается синтез дополнительной фракции ДНК, в пахитене идет репаративный синтез ДНК. Синтез гистонов продолжается до стадии пахитены. На этих же стадиях обнаруживаются специфические для М. структурные и ферментативные белки. С этими биосинтезами связывают специфические процессы профазы I — конъюгацию и кроссинговер. На субмикроскопическом уровне в конъюгирующих хромосомах обнаружены характерные для М. структуры — синаптонемальные комплексы. Образование сипаптонемальных комплексов начинается в лептотене. Они имеют вид осевого тяжа, состоящего в основном из белков и проходящего по всей длине хромосомы. В зиготене гомологичные хромосомы конъюгируют этими осевыми тяжами (боковыми элементами синаптонемальных комплексов). Считают, что именно эти комплексы обеспечивают временную конъюгацию хромосом по строго гомологичным локусам. Природа сил, осуществляющих сближение гомологичных хромосом перед их конъюгацией, остается нераскрытой. Противоречивы мнения относительно молекулярных механизмов конъюгации как таковой; существует представление, что она определяется взаимодействием не молекул ДНК, а белковых компонентов синаптонемальных комплексов. Не закончена также разработка молекулярной модели кроссинговера; многие данные говорят за то, что рекомбинация молекул ДНК в кроссинговере происходит путем разрыва обеих нитей ДНК и их воссоединения в каждой из гомологичных хроматид и осуществляется с помощью различных ферментов. В процессе М. осуществляются и другие хромосомные преобразования, связанные с дифференцировкой мейоцитов в зрелые гаметы. В лептотене начинается и в диплотене достигает своего максимального выражения транскрипционная активность многих хромосом и их участков, что происходит через деконденсацию функционирующих хромомеров и формирование боковых петель (хромосомы типа «ламповые щетки»). У животных и человека этот процесс происходит как в сперматоцитах, так и особенно — в ооцитах. Транскрибирующаяся ДНК обеспечивает синтезы многообразных белков, протекающие при формировании гамет. Высокую активность в этот период в оогенезе проявляют рибосомные гены. Большая потребность в рибосомной РНК покрывается многократным умножением (амплификацией) рибосомных генов. Цитол, выражением этого является увеличение числа и размеров ядрышек. В оогенезе человека это находит отражение в появлении в диплотене многих дополнительных ядрышек, не связанных с основными ядрышко образующими хромосомами, к-рыми являются пять акроцентрических хромосом.
Мейотический процесс находится под генетическим контролем, будучи хотя и высокоспецифическим, но частным случаем генетически регулируемой клеточной дифференцировки. Генетическая обусловленность М. в целом и его стадий в частности показана путем получения на растениях и животных мутаций, изменяющих течение М. Описаны мутации, при которых М. полностью отсутствует или не происходит первого или второго деления. Получены многочисленные мутации, влияющие на конъюгацию хромосом, частоту кроссинговера (хиазмообразования), терминализацию хиазм; мутации, нарушающие расхождение хромосом в анафазе I или II. Изучение мейотических мутантов позволяет расчленить весь процесс М. на элементарные события и через их морфол., биохимическую и иную характеристики глубже приблизиться к пониманию механизмов этого процесса.
Библиография: Основы цитогенетики человека, под ред. А. А. Прокофьевой-Бельговской, с. 144, М., 1969; Соколов И. И. Цитологические основы полового размножения многоклеточных животных, Руководство по цитол., под ред. А. С. Трошина и др., т. 2, с. 390, М.—Л., 1966, библиогр.; Цитология и генетика мейоза, под ред. В. В. Хвостова и Ю. Ф. Богданова, М., 1975, библиогр.; HultenM. a. Lindsten J. Cytogenetic aspects of human male meiosis, Advanc, hum. Genet., v. 4, p. 327, 1973, bibliogr.; J o h n B. a. L e w i s K. R. The meiotic system, Wien— N. Y., 1965, bibliogr.; Rhoades M. M. Meiosis, в кн.: The cell, ed. by J. Brachet a. A. E. Mirsky, v. 3, p. 1, N. Y.—L., 1961, bibliogr.
А. Ф. Захаров.
Источник: xn--90aw5c.xn--c1avg
Мейоз — это деление диплоидных клеток, в результате которого образуются гаплоидные клетки. То есть из каждой пары гомологичных хромосом материнской клетки в дочерние попадает лишь одна хромосома. Мейоз лежит в основе формирования половых клеток – гамет. В результате слияния мужской и женской гамет диплоидный набор восстанавливается. Таким образом, одно из важных значений мейоза — это обеспечение постоянства числа хромосом у вида при половом размножении.
В клетке, которая приступает к мейотическому делению, уже произошло удвоение (репликация) хромосом, также как это происходит в интерфазе митоза. Так что каждая хромосома состоит из двух хроматид, и количество хромосом диплоидное. То есть по количеству генетической информации клетки вступающие в митоз и мейоз одинаковы.
В отличие от митоза мейоз протекает в два деления. В результате первого деления гомологичные хромосомы каждой пары расходятся в разные дочерние клетки, и образуются две клетки с гаплоидным числом хромосом, но каждая хромосома состоит из двух хроматид. Второе деление протекает также как митотическое, т. к. происходит разделение хроматид каждой хромосомы, и в дочерние клетки попадает по одной хроматиде каждой хромосомы.
Таким образом в результате мейоза образуется четыре клетки с гаплоидным набором хромосом. У самцов все четыре становятся сперматозоидами. А вот у самок только одна становится яйцеклеткой, другие отмирают. Это связано с тем, что только в одной клетке концентрируется запас питательных веществ.
Стадии, или фазы, первого мейотического деления:
- Профаза I. Спирализация хромосом. Гомологичные хромосомы располагаются параллельно друг другу и обмениваются некоторыми гомологичными участками (конъюгация хромосом и кроссинговер, в результате которого происходит перекомбинация генов). Разрушается ядерная оболочка, начинает формироваться веретено деления.
- Метафаза I. Пары гомологичных хромосом располагаются в экваториальной плоскости клетки. К центромере каждой хромосомы присоединяется нить веретена деления. Причем к каждой только одна таким образом, что к одной гомологичной хромосоме присоединена нить с одного полюса клетки, а к другой – с другого.
- Анафаза I. Каждая хромосома из пары гомологичных отходит к своему полюсу клетки. При этом каждая хромосома продолжает состоять из двух хроматид.
- Телофаза I. Образуются две клетки, содержащие гаплоидный набор удвоенных хромосом.
Стадии, или фазы, второго мейотического деления:
- Профаза II. Разрушение ядерных оболочек, формирование веретена деления.
- Метафаза II. Хромосомы располагаются в экваториальной плоскости, к ним присоединяются нити веретена деления. Причем таким образом, что к каждой центромере присоединяются две нити — одна с одного полюса, другая — с другого.
- Анафаза II. Хроматиды каждой хромосомы разделяются в области центромер, и каждая из пары сестринских хроматид уходит к своему полюсу.
- Телофаза II. Формирование ядер, раскручивание хромосом, деление цитоплазмы.

На схеме показано поведение при мейозе только одной пары гомологичных хромосом. В реальных клетках их больше. Так в клетках человека содержится 23 пары. На схеме видно, что дочерние клетки генетически отличны друг от друга. Это важное отличие мейоза от митоза.
Следует отметить другое важное значение мейоза (первое, как уже было указано, – это обеспечение механизма полового размножения). В результате кроссинговера создаются новые комбинации генов. Они же создаются в результате независимого друг от друга расхождения хромосом при мейозе. Поэтому мейоз лежит в основе комбинативной изменчивости организмов, которая в свою очередь является одним из источников естественного отбора, т. е. эволюции.
Источник: scienceland.info
Половое размножение существует почти у всех растений и животных. Оно связано с образованием высокоспециализированных половых клеток — гамет. Гаметы формируются из диплоидных клеток путем специального типа клеточного деления — мейоза, в результате которого в клетках исходное число хромосом уменьшается вдвое (из диплоидного становится гаплоидным).
Несмотря на принципиальное сходство гаметогенеза у самых различных видов организмов, конкретные формы мейоза чрезвычайно разнообразны.
Мужские гаметы созревают в мужских половых железах — семенниках; этот процесс называется сперматогенезом. Женские гаметы созревают в яичниках в процессе овогенеза. В половых железах различают: зону размножения, зону роста и зону созревания; в зоне созревания гаметы окончательно формируются путем мейоза.
Мейоз происходит в результате двух последовательных делений родоначальной диплоидной клетки. Каждое из них включает четыре фазы. Все фазы первого мейотического деления обозначают цифрой I, а все фазы второго деления — цифрой II. Передпрофазой I в клетках, удваивается ДНК и в мейоз клетки вступают с хромосомным набором 2n4с.
В профазе I хромосомы вначале имеют вид тонких нитей, а затем утолщаются. Гомологичные хромосомы сближаются, в пунктах касания они перекрещиваются и обмениваются гомологичными участками- этот процесс называется кроссинговером (и представляет один из источников генотипической комбинативной изменчивости). Каждая хромосома в результате самоудвоения состоит из двух хроматид и называется унивалентой, а после сближения двух гомологичных хромосом (двух унивалент) образуются тетрады (биваленты). Как и в профазе митоза, в клетке в этот период формируется веретено деления, центриоли отходят к полюсам, оболочка ядра распадается, а тетрады движутся к центру клетки.
В метафазе I тетрады выстраиваются в плоскости экватора, гомологичные хромосомы в области центромер отходят друг от друга, оставаясь соединенными в области плеч. Нити веретена прикрепляются к центромерам гомологичных хромосом. Клетка вступает в третью фазу — анафазу I, во время которой нити веретена увлекают униваленты к противоположным полюсам. При этом одна из двух гомологичных хромосом случайно оказывается на одном полюсе, вторая — на другом. Именно в этот период происходит уменьшение вдвое (редукция) числа хромосом и их случайное перераспределение в будущих гаметах. В заключительной фазе клетка вступает в телофазу I. Таким образом, в итоге мейоза образуются две клетки, содержащие лишь по одной из двух гомологичных хромосом, каждая из которых состоит из двух хроматид. Хромосомы в результате кроссинговера обмениваются своими участками и несут, таким образом, перекомбинированный наследственный материал. Телофаза I длится недолго, и клетка переходит в интерфазу (краткую по времени), после которой наступает второе мейотическое деление. Во время интерфазы в отличие от митоза в клетках не происходит синтеза ДНК.
В профазе II по периферии ядра располагаются нитевидные хромосомы — униваленты, образуется веретено деления, хромосомы, приближаются к плоскости экватора и клетка вступает метафазу II. В анафазе II хроматиды расходятся и увлекаются нитями веретена от плоскости экватора к противоположным полюсам. Вслед за этим наступает телофаза II, во время которой хромосомы истончаются, образуя нити, и у полюсов формируются ядра дочерних клеток. В итоге из двух клеток мейоза I в телофазе мейоза II образуются четыре дочерние зрелые гаметы, жаждая из которых несет газплоидное число хромосом. Описанный процесс типичен для формирования мужских гамет. Образование женских половых клеток идет аналогично, но при овогенезе развивается лишь одна зрелая яйцеклетка, а три мелких редукционных тельца впоследствии отмирают.
Мейоз под микроскопом
Биологическое значение мейоза состоит в том, что:
1) образуются хромосомы обновленного генетического состава благодаря кроссинговеру между гомологичными хромосомами;
2) достигается наследственная разнородность гамет, так как во время первого мейотического деления из дары гомологичных хромосом в одну из двух гамет отходит материнская хромосома, в другую — отцовская;
3) после оплодотворения гаплоидные гаметы (1n1с) от отца и матери создают диплоидное ядро зиготы с числом хромосом, присущим данному виду.
Процессы сперматогенеза и овогенеза в принципе сходны, но между ними имеются и различия. В результате сперматогенеза образуется четыре сперматозоида, аовогенез завершается образованием одной яйцеклетки. Это обусловлено тем, что при первом и втором делениях созревания яйцеклетки не делятся пополам, а отделяют маленькие направительные, или редукционные, тельца. Направительные тельца несут полноценные хромосомные наборы, но практически лишены цитоплазмы и вскоре погибают. Биологический смысл образования этих телец заключается в необходимости сохранения в цитоплазме яйцеклетки максимального количества желтка, потребного для развития будущего зародыша.
Таблица Деление клеток (исходная клетка 2п 4с (n — хромосомы, с — хроматиды))
| Тип деления | Фазы | Набор хромосом в результате деления (n — хромосомы, с — хроматиды) |
Число и качество клеток, образую щихся в резуль тате деления |
Клетки, где происходит деление |
Распро-странение среди организмов |
| Митоз (непрямое деление) |
Интерфаза Профаза Метафаза Анафаза Телофаза |
2п 2с (дипло-идный), хромосомы однохрома-тидные |
Две дипло-идные | Сомати- ческие (клетки тела) |
Все животные и расти тельные организмы, кроме бактерий и синезеленых (прокариот) |
| Мейоз: мейоз I (ре дукцион-ное деление) мейоз II |
Интерфаза Профаза I Метафаза I Анафаза I Телофаза I Метафаза II |
In 2с (гапло-идный), хромосомы двухро- матидные 1n 1с (гапло-идный), хромосомы |
Две гапло-идные
Две гапло-идные Всего: четыре |
Половые клетки животных: при овогенезе образуются четыре клетки: одна яйцеклетка и три направи-тельных тельца (отмира- ющие); при сперма- тогенезе все клетки образуют сперма- тозоиды. Сяюрообра-зующие клетки растений: у семенных растений из четырех крупных спор три отмирают, одна остается; мелкие споры все остаются |
Все животные и растения, кроме прокариот |
Источник: www.examen.ru