Изогамия у водорослей
1. Типы размножения водорослей: вегетативное, бесполое, половое размножение. Примеры жизненных циклов с чередованием ядерных фаз и поколений у представителей различных отделов водорослей.
У водорослей различают бесполое и половое воспроизведение, причем бесполое размножение делится на вегетативное и собственно бесполое. Вегетативное размножение осуществляется:
— простым делением надвоепутем перешнуровки одетых оболочками одноклеточных форм. Обеспечивает равномерное распределение между двумя дочерними организмами всех компонентов материнской клетки, включая клеточные покровы, например, пелликулы у эвгленовых или теки у динофитовых.
– распадом колоний на отдельные части,
– фрагментацией слоевищ у многоклеточных.
У некоторых водорослей формируются специальные образования, служащие для вегетативного размножения: клубеньки харовых водорослей, выводковые почки у бурых.
кие как – акинеты – специальные клетки с утолщенными оболочками и большим запасом питательных веществ, служащие кроме размножения для переживания неблагоприятных условий. Акинеты известны, например, у синезеленых водорослей (Anabaena, Aphanizomenon), нитчатых зеленых водорослей (например, Ulothrix) и некоторых других. У более высоко организованных зеленых, бурых, красных и харовых водорослей вегетативное размножение осуществляется за счет дополнительных побегов и столонов (стелющихся по грунту нитей из нескольких рядов клеток у бурых водорослей, например Sphacelaria).
Собственно бесполое размножение осуществляется при помощи специальных спор. При их образовании протопласт недифференцированной материнской клетки или дифференцированной (спорангия) преобразуется и в ней формируется одна или несколько спор. Они могут быть подвижными (=зооспоры), имеющие жгутик и неподвижными. В образовании оболочек спор оболочка материнской клетки участия не принимает. Спорообразование сопровождается выходом продуктов деления материнской клетки или спорангия из оболочки, что отличает собственно бесполое размножение от вегетативного. Организм возникает из гаплоидной клетки. При освобождении спор из спорангия в его оболочке образуется отверстие или щель, или оболочка спорангия разрушается на две или более частей, или ослизняется.
По способу образования различают экзо- и эндоспоры.
Экзоспоры (конидии) характерны для цианопрокариот, например, Chamaesiphon. Они возникают как выросты протопласта на вершине клетки водоросли в результате ее поперечного деления и дальнейшего отчленения.
Эндоспоры (зооспоры, спорангиоспоры) возникают внутри материнских клеток или внутри специализированных спорангиев и наблюдаются, например, у водоросли Endonema.
Различают имеющие монадную структуру зооспоры и неподвижные апланоспоры. Зооспоры могут иметь от одного до многих жгутиков, которые в этом случае располагаются венцом на ее переднем конце (например, у зеленой водоросли Oedogonium) или парами по всей поверхности (желтозеленая водоросль Vaucheria). Апланоспоры – это неподвижные, лишенные жгутиков споры. К ним относятся моно-, би- и тетраспоры красных водорослей, образующиеся в спорангии в количестве одной, двух или четырех (Callithamnion, Ceramium), тетраспоры бурых водорослей (например, Dictyota). Апланоспоры, которые еще внутри спорангия принимают форму материнской клетки, называются автоспорами (зеленые водоросли Chlorella, Scenedesmus). В том случае, когда апланоспоры после выхода из спорангия переходят в состояние длительного покоя, вырабатывая утолщенную оболочку, они называются гипноспорами. Зооспоры, утратившие жгутики, но сохранившие пульсирующие вакуоли и стигму, называют гемизооспорами. У подавляющего числа видов растений в спорангиях образуется большое количество спор.
Половое размножение (воспроизведение) у водорослей сопровождается половым процессом, заключающемся в слиянии двух гаплоидных клеток или гамет, различных в половом отношении. При этом образуется диплоидная зигота, вырастающая в новую особь или формирующая зооспоры. Половой процесс может быть нескольких типов и осуществляется в простейших случаях без формирования специализированныхклеток или гамет, и с образованием гамет.
-
Простейшим случаем полового процесса считается гологамия или хологамия (holos – целый). При этом происходит слияние двух целых подвижных вегетативных клеток, лишенных клеточной оболочки. При слиянии протопластов двух безжгутиковых вегетативных клеток половой процесс называется конъюгацией. Это единственная форма полового процесса у водорослей класса Zygnematophyceae(Conjugatophyceae). Между двумя конъюгирующими вегетативными клетками формируется конъюгационный канал, по которому продвигаются протопласты либо одной, либо обеих сливающихся клеток, выполняющих функцию гамет.
-
Изогамия – сливаются гаметы одинаковые по величине, строению и подвижности.
-
Под гетерогамией (heteros – разный) или анизогамией понимают такой тип полового процесса, при котором сливаются две подвижные гаметы, имеющие различные размеры, но одинаковую форму. При этом более крупная гамета (макрогамета) считается женской, а более мелкая (микрогамета) – мужской. Гетерогамный половой процесс известен у динофитовых, бурых и зеленых водорослей.
-
Оогамия – это такой тип полового процесса, при котором утратившая подвижность более крупная женская гамета, или яйцеклетка, сливается с подвижной, имеющей жгутики, мужской гаметой, или сперматозоидом (антерозоидом). Гаметангии, в которых формируются яйцеклетки, называются оогониями, а в которых образуются сперматозоиды – антеридиями. Мужские и женские гаметы могут развиваться на одной особи или колонии (однодомные или обоеполые виды) или на разных (двудомные или раздельнополые виды). Оогамия является наиболее высоко организованным типом полового процесса, преобладающим у растений. Она известна у желтозеленых, диатомовых, бурых, зеленых, является единственным типом полового процесса у харовых и красных водорослей.
После слияния гамет образуется диплоидная (2n) зигота; гаметы всегда гаплоидны (n) и содержат одинарный набор хромосом. Зигота прорастает или сразу после образования, или после некоторого периода покоя. У многих водорослей зигота после некоторых изменений превращается в покоящуюся стадию – зигоспору (ооспору). В зигоспоре происходит слияние двух гаплоидных ядер с образованием диплоидного ядра. Зигоспоры дают начало либо новому диплоидному таллому водоросли, либо прорастают после мейоза четырьмя гаплоидными зооспорами, из которых формируются взрослые гаплоидные растения. При этом иногда сохраняется жизнеспособным только одно или два ядра.
У водорослей возможно развитие женских гамет без оплодотворения– партеногенез. Такие развивающиеся без оплодотворения женские гаметы называются партеноспорами или азиготами.
Особым типом полового процесса является автогамия, встречающаяся, например, у диатомовых водорослей. При этом ядро диплоидной клетки делится мейотически на четыре гаплоидных ядра, два из которых разрушаются, а оставшиеся два сливаются в диплоидное, образуя зиготу, которая без периода покоя резко увеличивается в размерах и превращается в особую спору роста – ауксоспору. После созревания ауксоспора развивается в новую клетку, по размерам много превышающую материнскую. Автогамия не сопровождается попарным слиянием двух клеток и увеличением числа особей, происходит только их омоложение; ауксоспора образуется из одной клетки.
У многих водорослей одного вида часто наблюдаются разные формы размножения, бесполое и половое, часто закономерно сменяющие друг друга более или менее регулярно. Развитие вида от одной до другой такой же стадии получило название жизненного цикла. Соотношение гаплоидной и диплоидной фаз в жизненном цикле (ЖЦ) у разных водорослей неодинаково.
1. Зиготический (гаплоидный) ЖЦ – в вегетативном состоянии водоросли являются гаплонтами (содержат гаплоидный набор хромосом). Единственной диплоидной стадией в их жизненном цикле является зигота. Мейоз и редукция числа хромосом при гаплофазном жизненном цикле наблюдается при прорастании зиготы (зиготическая редукция). Гаплофазный цикл наблюдается, например, у многих зеленых водорослей (н-р, вольвоксовые).
2. Гаметический (диплоидный) ЖЦ — большую часть жизненного цикла проводят в диплоидном состоянии, гаплоидная стадия представлена только гаметами. диплоидная зигота без редукции числа хромосом прорастает в диплоидное растение (диплонт). Гаметический мейоз. К таким водорослям относятся, например, зеленые сифоновые, диатомовые, циклоспоровые водоросли из бурые.
3. Спорический (диплогаплофазный) ЖЦ – чередование форм развития или генераций: диплоидного бесполого поколения (спорофита) и гаплоидного полового поколения (гаметофита). На диплоидном спорофите с редукцией числа хромосом (спорическая редукция) формируются подвижные зооспоры или неподвижные апланоспоры, прорастающие в гаплоидный гаметофит. На гаплоидном гаметофите формируются половые органы и гаметы. Если оба поколения морфологически одинаковы, то такая смена форм развития называется изоморфной, если различны – то гетероморфной. Изоморфная смена форм развития наблюдается, например, у морских видов Ulva, Enteromorpha, Cladophora из зеленых водорослей, ряда бурых и большинства красных водорослей (несколько усложненная). Гетероморфная смена есть у бурых, встречается у зеленых и красных водорослей.
4. Соматический ЖЦ — мейоз происходит в некоторых соматических клетках диплоидного таллома (соматическая редукция). Из получившихся гаплоидных клеток вырастает половое поколение – гаметофит. У немногих видов водорослей, например у Prasiola (зеленые), Lemanea, Batrachospermum (красные).
 Жизненные циклы водорослей:
Жизненные циклы водорослей:
I – зиготический; II – гаметический; III – спорический.
Доминирующая фаза в случаях I и III бывает многоклеточной, а если она одноклеточная, то наиболее долговечная и способная к митотическому воспроизведению.
У ряда водорослей, имеющих смену форм развития, в результате спонтанного увеличения числа хромосом, партеногенетического развития гамет и других процессов, связанных с трансформацией генома, способ размножения (бесполое и половое) не всегда коррелирует с плоидностью таллома. Спорофиты не всегда могут быть только диплоидными, наблюдаются и гаплоидные, и тетраплоидные, а гаметофиты обнаруживают не только гаплоидный, но и диплоидный набор хромосом. Внешние условия развития водоросли могут способствовать выпадению отдельных стадий из жизненного цикла и т. д.
Источник: StudFiles.net
Бесполое размножение у одноклеточных водорослей осуществляется путем деления клетки, у колониальных и нитчатых – в результате распада колоний или нитей на отдельные фрагменты; у немногих водорослей образуются специальные органы размножения, например, клубеньки у харовых, акинеты (особые клетки с большим количеством запасных веществ и пигментов) – у зеленых и др. Такое размножение часто называют вегетативным.
Бесполое размножение происходит также посредством неподвижных спор (апланоспор) или зооспор (спор со жгутиками), образующихся путем деления протопласта обычных или особых клеток, называемых спорангиями. У ряда представителей зеленых водорослей апланоспоры уже в материнской клетке иногда приобретают все отличительные черты этой клетки. В таких случаях говорят об автоспорах. Размножение при помощи спор называется собственно бесполым размножением.
Половое размножение характеризуется наличием полового процесса, одним из важнейших этапов которого является оплодотворение, т.е. слияние гаплоидных половых клеток – гамет. В результате оплодотворения образуется зигота с новой комбинацией наследственных признаков, которая и становится родоначальницей нового организма.
У водорослей различают следующие формы полового процесса: хологамию – слияние двух одноклеточных особей; изогамию – слияние одинаковых по строению и величине подвижных гамет; гетерогамию – слияние подвижных гамет разных размеров (более крупную считают женской); оогамию – слияние крупной неподвижной яйцеклетки с мелкой подвижной мужской гаметой – сперматозоидом или неподвижным, лишенным оболочки спермацием (у красных водорослей); конъюгацию – слияние протопластов неспециализированных клеток.
меты образуются в клетках, не отличающихся от вегетативных, или в особых клетках, получивших название гаметангии. Гаметангии, содержащие яйцеклетку (редко несколько), называются оогониями, а те, в которых формируются сперматозоиды или cпермации, – антеридиями.
У примитивных водорослей каждая особь способна формировать и споры, и гаметы в зависимости от времени года и внешних условий; у других функции бесполого и полового размножения выполняют разные особи – спорофиты (образуют споры) и гаметофиты (образуют гаметы). В жизненном цикле ряда водорослей происходит строгое чередование поколений – гаметофита и спорофита.
Гаплофазный тип характеризуется отсутствием чередования поколений. Вся вегетативная жизнь водорослей проходит в гаплоидном состоянии, т. е. они являются гаплонтами. Диплоидна лишь зигота, прорастание которой сопровождается редукционным делением ядра (зиготическая редукция). Развивающиеся при этом растения оказываются гаплоидными. Примером являются многие зеленые (вольвоксовые, большинство хлорококковых, конъюгаты) и харовые водоросли.
Диплофазный тип отличается тем, что вся вегетативная жизнь водорослей осуществляется в диплоидном состоянии, а гаплоидная фаза представлена только гаметами. Перед их образованием происходит редукционное деление ядра (гаметическая редукция). Зигота без деления ядра прорастает в диплоидный таллом. Эти водоросли являются диплонтами. Такой тип развития характерен для многих зеленых водорослей, имеющих сифоновую структуру, всех диатомовых и некоторых представителей бурых (порядок Фукальные).
Диплогаплофазный тип характеризуется тем, что в клетках диплоидных талломов (спорофитов) многих водорослей редукционное деление ядра предшествует образованию зоо- или апланоспор (спорическая редукция). Споры развиваются в гаплоидные растения (гаметофиты), размножающиеся только половым путем. Оплодотворенная яйцеклетка – зигота – прорастает в диплоидное растение, несущее органы бесполого размножения. Таким образом, у этих водорослей имеет место чередование форм развития (генераций): диплоидного бесполого спорофита и гаплоидного полового гаметофита.
Оба поколения по внешнему виду могут не различаться и занимать одинаковое место в цикле развития (изоморфная смена генераций) или же резко различаться по морфологическим признакам (гетероморфная смена генераций). Изоморфная смена генераций характерна для ряда зеленых (ульва, энтероморфа, кладофора), бурых и большинства красных водорослей. Гетероморфная смена генераций встречается с преобладанием как гаметофита, так и спорофита (свойственна преимущественно бурым, реже зеленым и красным водорослям).
Источник: students-library.com
Изогамия — половое размножение при использовании одинаковых гамет.[ …]
У бурых водорослей с половым процессом в виде изогамии подвижные клетки (зооиды), которые образуются на гаметофитах, функционируют или как гаметы, сливаясь попарно и воспроизводя спорофиты, или как зооспоры, производя снова слоевища гаметофитов. Такое бесполое размножение гаметофитов у представителей вышеперечисленных порядков бурых водорослей распространено крайне широко. Иногда в лабораторных культурах наблюдается подряд несколько поколений «гаметофитов», не приступающих к половому размножению. Таким образом, они проявляют себя как спорофиты и как гаметофиты, и их лучше называть гаметоспорофитами, чтобы не путать с другими растениями, гаметофиты которых не способны к бесполому размножению спорами.[ …]
Половой процесс у бурых водорослей представлен изогамией, гетерогамией и оогамией, гетерогамия встречается реже двух других форм полового размножения.[ …]
Половой процесс у протококковых водорослей наблюдается сравнительно редко. Обычно это изогамия, реже гетерогамия. Оогамный половой процесс известен только у очень немногих родов. Наиболее обычными стадиями покоя у протококковых водорослей являются акинеты и цисты.[ …]
У современных представителей порядка известно только половое размножение. Половой процесс — изогамия. Гаметы образуются в специальных гаметангиях. У большинства дазикладовых гаметангии возникают как шаровидные выросты на вершине или сбоку ветвей первого порядка и соответствуют видоизмененным ветвям второго порядка (рис. 232, 3, 4). У аце-табулярии гаметангии образуются на коротких сегментах первого порядка как специальные выросты (рис. 232, 9). Внутри гаметангия образуются толстостенные многоядерные цисты со специальной крышечкой (рис. 232, 10). Зрелые цисты содержат множество гамет. При разрушении стенок гаметангия цисты выходят во внешнюю среду и уже здесь из них высвобождаются гаметы. Цисты, а не зиготы могут служить покоящимися стадиями. Существует мнение, что образование цист связано с обызвествлением слоевища. У некоторых лишенных извести родов их нет и гаметы образуются непосредственно в гаметангиях.[ …]
Кроме бесполого размножения зооспорами, у этого вида наблюдается и половой процесс в виде типичной изогамии. Интересны здесь гаметы. Они имеют копуляционные хоботки, выступающие через оболочку между основаниями жгутиков (рис. 205, 1, 2), что и послужило основой для видового названия.[ …]
Бесполое размножение осуществляется при помощи зооспор с одним волочащимся сзади жгутиком. Половое размножепие представляет собой слияние подвижных гамет или одинаковых по величине и строению (изогамия), или различных (гетерогамия), образующихся в гаметангиях. При этом подвижные клетки (зооспоры, гаметы) бластокладиевых отличаются от зооспор и гамет других грибов тем, что они более активно движутся, а также наличием хорошо заметной ядерной шапочки, состоящей из массы РНК (см. рис. 4).[ …]
При гетеро- и оогамии мужские и женские гаметы могут развиваться на одной особи или колонии (обоеполые, или однодомные, виды) или на разных (раздельно-полые, или двудомные, виды). Среди водорослей, для которых характерна изогамия, существуют гомоталличные виды (у них сливаются гаметы из одного слоевища или колонии) и гетероталличные (слияние возможно только между гаметами из разных особей), которые ввиду отсутствия морфологических различий обозначаются знаками + и —, соответственно различают — -гаметы и —гаметы.[ …]
У грибов класса хитридиомицетов вегетативное тело представляет собой или плазмодий, или очень слабо развитый мицелий (р и з о-мицелий). Им свойственно бесполое размножение зооспорами с одним задним жгутиком.[ …]
Указанные ценобиальные формы вольвоксо-вых встречаются довольно часто, обычно в небольших стоячих водоемах, где летом нередко вызывают зеленое «цветение» воды. Размножаются они в основном бесполым способом, путем образования в каждой клетке молодых ценобиев. Во всех случаях образовавшаяся зигота переходит в состояние покоя.[ …]
Размножаются золотистые водоросли простым делением клетки (рис. 66, 4), а также путем распада колоний или многоклеточного таллома на отдельные части. Известен также половой процесс в виде типичной изогамии, хо-логамии или автогамии. В результате лолового процесса образуются очень разнообразные по характеру скульптурированности оболочки эндогенные кремнистые цисты (рис. 68, 2; 73, 3), которые помогают золотистым водорослям переживать неблагоприятные условия.[ …]
Макроскопическое слоевище является спорофитом и несет шаровидные или яйцевидные одногнездные спорангии. Гаме-тофит микроскопический, состоит из стелющихся нитей, несущих многогнездные гаметангии. Половой процесс — изогамия. Из зиготы сначала вырастают стелющиеся нити — протонема, на которой позднее появляется макроскопическое слоевище спорофита. Полный цикл у стриарии оттянутой осуществляется не всюду: он известен для этих водорослей у берегов Франции; у индивидов же, растущих в Северном море у острова Гельголанд, в культурах наблюдалось чередование только бесполых слоевищ с одногнездными спорангиями.[ …]
Половое размножение является наиболее эффективным путем воспроизводства организмов, дающим возможность «перетасовки» и комбинирования генов. Предполагают, что оно развилось из бесполого, возникнув около 1 млрд лет назад, причем первые этапы в этом процессе были связаны с усложнением в развитии гамет. Примитивные гаметы характеризовались недостаточной морфологической дифференцировкой, в результате чего для многих организмов ведущей была изогамия (от греч. isos — равный, gamos — брак), когда половые клетки были подвижными изогаметами, еще не дифференцированными на мужские и женские формы. Изогамия встречается у ряда видов простейших.[ …]
Филогенетически бластокладиевые можно рассматривать как результат дальнейшей эволюции хитридиевых грибов, которая шла, во-первых, по пути увеличения размеров центральной части таллома (уже крупного, например, у МасгосЬуШшп Ьо1гу(Пос1е8). Во-вторых, форма клетки так же изменялась. В-третьих, осуществлялся переход от изогамии к гетерогамии и выработалась изоморфная смена поколений.[ …]
К этому порядку относятся наиболее примитивные представители класса, вегетативные клетки которых всегда неподвижны, но на всех стадиях развития сохраняют еще пульсирующие вакуоли (рис. 210). У некоторых из них сохраняются также глазок и редуцированные, ставшие неподвижными жгутики, называемые здесь псевдоцилиями (табл. 29, 9). Размножение осуществляется главным образом зооспорами, реже гемизооспорами. У колониальных форм, кроме того, наблюдается деление клеток, за счет которого происходит разрастание таллома. Половой процесс в виде изогамии или гетерогамии известен у немногих видов.[ …]
Цикл развития хордариевых заключается в чередовании большей частью крупного спорофита и микроскопического гаметоспорофита. У хордариевых имеется плетизмоталлюс — микроскопический спорофит, который несет многогнездные спорангии и может в виде выроста производить макроскопическое слоевище спорофита. Из зооспор снова вырастают слоевища плетизмоталлюса. Макроскопические слоевища спорофитов у хордариевых производят одногнездные или многогнездные спорангии или и те и другие одновременно. Мейоз происходит в одногнездных спорангиях. Гаме-тоспорофит хордариевых состоит их стелющихся разветвленных нитей, на которых развиваются многогнездные вместилища. Половой процесс—изогамия, крайне редко — гетерогамия. Зигота прорастает в растеньице, которое сначала внешне подобно гаметоспорофиту, а позднее производит макроскопическое слоевище спорофита или остается в виде плетизмоталлюса. Зооиды без полового процесса снова дают начало гаметоспорофитам. Из спор, производимых макроскопическими спорофитами, могут вырастать как гаметоспорофиты, так и спорофиты. Все это приводит к тому, что в цикле развития хордариевых отсутствует строгое чередование форм развития.[ …]
У сфацеляриевых существует изоморфная смена форм развития. Диплоидные спорофиты могут нести многогнездные и одногнездные спорангии. Зооспоры из многогнездных спорангиев возобновляют диплоидные растения. Мейоз происходит в одногнездных спорангиях, и гаплоидные зооспоры дают гаметоспорофиты, на которых развиваются только многогнездные вместилища. Образующиеся в них зооиды могут прорастать в растения материнского типа или функционировать как гаметы. У сфацеляриевых гаметы, как правило, одинаковые по размерам, но имеют физиологические различия. Мужские гаметы дольше сохраняют подвижность. Слияние гамет наступает после того, как женская гамета становится неподвижной. В тех случаях, когда сливаются подвижные гаметы, зигота может сохранять подвижность в течение нескольких часов. Значительно реже, чем изогамия, у сфацеляриевых наблюдается гетерогамия; при этом образуются многогнездные гаметангии, различающиеся размерами камер. У сфацеляриевых известно несколько случаев оогамии.[ …]
Самая яркая отличительная черта конъюгат, позволяющая объединять их вместе в особый класс, несмотря на разнообразие внешнего вида,— это полное отсутствие подвижных стадий размножения, т. е. зооспор и подвижных гамет, и наличие своеобразного полового процесса, получившего специальное название — конъюгация (лат. сощ аНо — слияние, соединение). Во время конъюгации происходит слияние протопластов двух клеток, которые выполняют здесь функцию гамет. В результате такого слияния, происходящего через специально образующийся канал, получается одноклеточная, покрытая толстой оболочкой зигота. Внешне две сливающиеся (конъюгирующие) клетки совершенно неразличимы; физиологическая дифференциация на мужскую и женскую клетки хотя и определенна, но узнать ее можно лишь косвенно и спустя некоторое время — по положению зиготы. В том случае, когда скорости передвижения (вернее, перетекания) сжавшихся протопластов-гамет одинаковы, зигота образуется в конъюгационном канале, иногда разрастаясь вплоть до степок конъюгирующих клеток-гаметангиев. В этом случае разделение клеток на мужскую и женскую крайне условно, а сам процесс слияния равноценных по морфологии и скорости перетекания гамет принято называть изогамией (греч. «изос»— равный, «гамос»— брак). Эта форма полового процесса характерна для большинства водорослей-конъюгат. И здесь протопласты-гаметы обеих конъюгирующих клеток одинаковы по размерам, но одна из них (мужская) активно перетекает через конъюгационный канал из отдающей клетки в воспринимающую, где и сливается со сравнительно неподвижной гаметой (женской). В этом случае зигота формируется только в воспринимающей (женской) клетке.[ …]
Источник: ru-ecology.info
РАСТЕНИЯ
НИЗШИЕ РАСТЕНИЯ
ВОДОРОСЛИ
Размножение водорослей
Водоросли увеличивают свою численность различными способами, важнейшими из них являются вегетативное, бесполое и половое размножение.
Вегетативное размножение представляет собой деление клетки одноклеточных водорослей или отделение по различным причинам от слоевища частей (если рассматриваются многоклеточные формы). Таких причин может быть очень много. У нитчатых форм это расщепление нити на две новые нити или распадение их на одно- или многоклеточные части. Слоевища истинно многоклеточных водорослей могут разделяться из-за механического воздействия потоков воды, различных водных обитателей, деятельности человека и т. д. Оторванные части водорослей, прикрепленных к подводному субстрату и входящих в состав бентоса, как правило, не погибают, но их дальнейшее развитие отличается от прикрепленных (бентосных) растений. В частности, у них повторно не образуются ризоиды, поэтому оторванные части переносятся током воды, часто сбиваясь в крупные массивы. Нарастание слоевища при этом не прекращается, но и здесь могут быть обнаружены некоторые морфологические особенности. Другой особенностью оторванных частей является их неспособность образовывать собственные органы размножения (как полового, так и бесполого).
Многие водоросли для вегетативного размножения образуют специальные структуры. Например, некоторые бурые водоросли образуют на талломах своеобразные «почки», которые затем отламываются и дают начало новым слоевищам. Как уже упоминалось, у харовых водорослей на ризоидах и погруженных в грунт участках «стеблей» образуются одно- или многоклеточные клубеньки, которые после периода зимнего покоя прорастают в новые растения. У некоторых нитчатых водорослей при наступлении неблагоприятных условий определенные клетки утолщают свою оболочку и накапливают в протопласте большой запас питательных веществ. Такие клетки способны долгое время находиться в состоянии покоя, что позволяет им переждать неприятную для них ситуацию. Название покоящихся клеток — акинеты указывает на их статичность и неподвижность. От спор они отличаются тем, что оболочка акинеты образуется непосредственно из оболочки клетки, а не внутри нее.
Бесполое размножение осуществляется с помощью различного рода спор. Их образование происходит внутри соответствующих клеток и всегда сопровождается выходом из оболочек последних (этим они отличаются от акинет). Споры развиваются или в обычных вегетативных клетках, или в особых, именуемых спорангиями, которые морфологически отличаются от вегетативных клеток.
Развитие спор сопровождается делением ядра, которое может происходить несколько раз в зависимости от количества образующихся спор (причем у разных групп водорослей ядро может делиться путем митоза или мейоза). Они в дальнейшем окружаются более или менее плотной оболочкой и выходят через образующееся отверстие в оболочке исходной материнской клетки.
Классификация спор осуществляется по различным критериям. Если споры снабжены жгутиками и способны активно передвигаться, они называются зооспорами. В тех случаях, когда споры имеют плотную оболочку и лишены жгутиков, а потому неподвижны, их называют апланоспорами. Если образующаяся оболочка имеет очень большую толщину, спора способна к длительному покою и называется гипноспорой. Следует отметить, что большинство водорослей образуют зооспоры. В других случаях при определении спор указывается их численность в материнской клетке или спорангии — моноспоры, биспоры, тетраспоры и т. д.
Половое размножение, в отличие от бесполого, всегда сопровождается слиянием двух клеток, у которых в ядрах имеется лишь по одному набору хромосом, характерному для этого вида организмов, т. е. эти клетки гаплоидны. В результате образуется диплоидная клетка — зигота, из которой развивается водоросль или же формируются зооспоры.
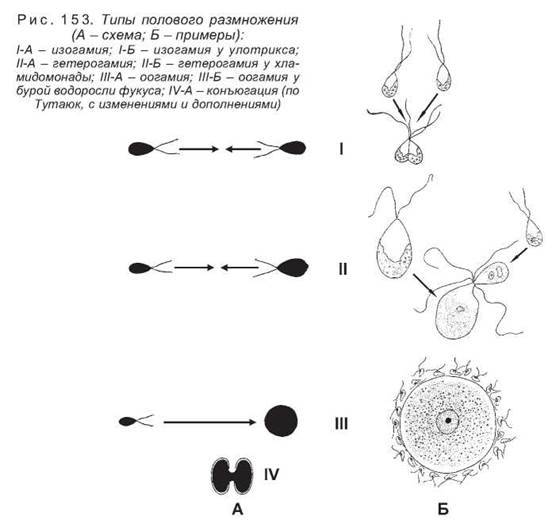
Если сливаются две обычные вегетативные клетки, процесс называется хологамией, такой процесс свойствен некоторым колониальным жгутиковым. Если сливаются вегетативные клетки, не имеющие жгутиков, половой процесс именуется конъюгацией (некоторые зеленые водоросли). У остальных водорослей сливаются клетки, специализированные для полового размножения, — гаметы, которые образуются в половых органах — гаметангиях. Подвижные гаметы принято считать мужскими (они часто имеют меньшие размеры), а неподвижные — женскими. В ряде случаев гаметы выглядят более или менее одинаково и тогда их половую принадлежность определить трудно. В зависимости от строения сливающихся гамет выделяют три типа полового размножения — изогамию, анизогамию и оогамию (рис. 153).
Изогамия — это слияние двух совершенно одинаковых гамет.
Анизогамия или гетерогамия представляет собой слияние гамет, различающихся между собой по размерам или подвижности.
Оогамия является наиболее высокоорганизованным типом полового размножения. В нем участвуют гаметы, отличающиеся как размерами, так и подвижностью. Крупную и неподвижную называют яйцеклеткой, она содержит запас питательных веществ, необходимый для последующего развития зиготы, а значительно более мелкую гамету — сперматозоидом или антерозоидом, она снабжена жгутиком и способна активно перемещаться в поисках яйцеклетки.
Напомним, что гаметы образуются в особых структурах, являющихся, по существу, половыми органами. Женские органы называются оогониями (их не следует путать с одноименными первичными женскими половыми клетками животных), мужские — антеридиями. За исключением харовых водорослей, которые имеют многоклеточные половые органы, у остальных водорослей органы полового размножения состоят из одной клетки.
У некоторых водорослей яйцеклетки и сперматозоиды могут образовываться на разных талломах. Такие формы называют двудомными, в отличие от однодомных, у которых на одном слоевище развиваются гаметы обоих типов. Это относится к водорослям, у которых гаметы различаются между собой. У изогамных форм сливающиеся гаметы также могут образовываться на одном растении, такие водоросли называются гомоталличными, или на разных — у гетероталличных видов. Для различения морфологически одинаковых талломов, но продуцирующих разные типы гамет, используют простые обозначения — плюс (+) и минус (-) талломы. Аналогично называются и гаметы, которые они образуют.
Источник: compendium.su